










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkTerra Latinoamericana
versión On-line ISSN 2395-8030versión impresa ISSN 0187-5779
Terra Latinoam vol.26 no.1 Chapingo ene./mar. 2008
División II
Dosis de nitrógeno y su efecto en la producción y distribución de biomasa de algodón transgénico*
Nitrogen Rate and its Effect on Biomass Production and Distribution of Transgenic Cotton
Jorge A. Orozco–Vidal1, Arturo Palomo–Gil1‡, Emiliano Gutiérrez–Del Río1, Armando Espinoza Banda1 y Vicente Hernández–Hernández1
1 Universidad Autónoma Agraria Antonio Narro, Unidad Laguna. Periférico y Carretera a Santa Fe, Colonia Centro, 27000 Torreón, Coahuila, México. ‡Autor responsable (apalomog@mixmail.com)
Recibido: mayo de 2005.
Aceptado: junio de 2007.
RESUMEN
El objetivo del presente estudio fue determinar el efecto de la dosis de nitrógeno (N) en la producción y asignación de biomasa y en los indicadores del crecimiento de la variedad transgénica NuCot 35B de algodón (Gossypium hirsutum L.). Las dosis evaluadas fueron 0, 80 y 160 kg ha–1 de N, las cuales se distribuyeron en un diseño de bloques al azar con seis repeticiones. La siembra se realizó en el sistema de producción de surcos estrechos (0.76 m entre surcos y distancia de 0.20 m entre plantas) para una población de 65 500 plantas ha–1. Para determinar el efecto del N en la dinámica de crecimiento y eficiencia en la producción y distribución de biomasa se realizaron cuatro muestreos destructivos a 69, 82, 105 y 124 días después de la siembra (DDS). En cada muestreo se colectaron dos plantas por parcela para determinar área foliar, peso seco total y peso seco de órganos vegetativos y reproductivos. Con los datos de área foliar y peso de biomasa seca se estimó la tasa de crecimiento del cultivo (TCC), tasa de asimilación neta (TAN), índice de área foliar (IAF), relación de área foliar (RAF), área foliar específica (AFE) y relación de peso foliar (RPF). El N aplicado no afectó la tasa de producción y distribución de biomasa, ni los indicadores de la magnitud de fotosíntesis. A 124 DDS los órganos reproductivos acumulaban 60% del peso seco total de la planta. La mayor TCC (22.1 g m–2 d–1) se presentó entre 69 y 105 DDS. La dosis de N tampoco afectó RAF, AFE y RPF, lo cual indica que, en todas las fases de crecimiento, las plantas acumularon la misma cantidad de fotoasimilados en sus hojas. La RPF obtenida indica que, independientemente de la cantidad de N aplicado, las plantas asignan el mismo porcentaje de fotoasimilados a su estructura foliar.
Palabras clave: Gossypium hirsutum L., materia seca, tasa de crecimiento, tasa de asimilación neta, índice de área foliar.
ABSTRACT
The objective of this work was to study growth indicators and the effect of nitrogen (N) rate on biomass production and distribution of the transgenic cotton (Gossypium hirsutum L.) variety NuCot 35B. N rates were 0, 80, and 160 kg ha–1. A complete randomized block design with six replications was used. Planting was done in the narrow–row cotton production system (rows 0.76 mapart). Adistance of 0.20 m betweenplants resulted in a population density of 65 500 plants ha–1. Four destructive samplings were performed 69, 82, 105, and 124 days after planting (DAP). Plant leaf area, total dry weight, and dry weight of plant and reproductive organs from two plants per plot were measured on each sampling date. Total dry weight and plant leaf area data were used to estimate crop growth rate (CGR), net assimilation rate (NAR), leaf area index (LAI), leaf area ratio (LAR), specific leaf area (SLA), and leaf weight ratio (LWR). N rate did not affect total biomass production, biomass partitioning, photosynthetic apparatus size indicators, nor photosynthetic efficiency of the variety studied. At 124 DAP, 60% of the total plant dry weight was accumulated in the reproductive organs. The highest CGR (22.1 g m–2 d–1) was observed between 69 and 105 DAP. N rate did not affect the LAR, SLA, and LWR estimator which means that, in all growth phases, the plants accumulated the same quantity of photoassimilates in their leaf sheet. According to LWR, and independently of N rate, plants assigned the same percentage of photoassimilates to their respective foliar structure.
Keywords: Gossypium hirsutum L., dry matter, growth rate, net assimilation rate, leaf area index.
INTRODUCCIÓN
El crecimiento y rendimiento del algodón (Gossypium hirsutum L.), al igual que la mayoría de las especies cultivadas, muestra alta dependencia al nitrógeno (N) y al agua durante su ciclo biológico. El N es el nutrimento más crítico en un programa de fertilización en virtud de que es esencial para un desarrollo óptimo del cultivo, pero hay que evitar excesos que puedan ocasionar serios problemas de manejo del cultivo, y pérdidas en la producción. McConnell et al. (1989) enfatizan que la sobrefertilización produce plantas con gran crecimiento vegetativo sin que esto se refleje en el rendimiento, además de que se incrementan las probabilidades de que se pierda el N del sistema suelo–planta. Dosis altas de N también impactan negativamente el medio ambiente, ya que aumentan la contaminación de aire, suelo, plantas y acuíferos. La contaminación del aire es producto de la volatilización del N en forma de óxido nitroso, el cual destruye la capa de ozono en la estratosfera, creando "agujeros de ozono" (Wojcik, 2001).
La dosis óptima de N está determinada por muchas variables ambientales, como clima, tipo de suelo, cultivar, fertilidad residual, humedad disponible, plagas, entre otros factores (Gaylor et al., 1983; McConnell et al., 1989). Entre 1960 y 1970, en la Comarca Lagunera se realizaron numerosos estudios para determinar la dosis óptima de fertilización nitrogenada para el cultivo del algodón, la cual se situó en 120 a 150 kg ha–1 de N. Como estos estudios se realizaron con variedades más frondosas y de ciclo más largo que las variedades actualmente cultivadas, a partir de 1992 se reanudaron las investigaciones sobre fertilización nitrogenada de las cuales se concluyó que las nuevas variedades sólo requieren de 80 kg ha–1 de N para mostrar su potencial productivo (Palomo et al., 1999; 2003).
El rendimiento de algodón depende en gran proporción de la cantidad de biomasa producida y de la eficiencia con que ésta es asignada a las estructuras reproductivas, lo cual es medido mediante el índice de cosecha. Wells y Meredith (1984) y Unruh y Silvertooth (1996) indican que la superioridad productiva de las nuevas variedades se debía a que acumulaban una mayor cantidad de materia seca en los órganos reproductivos y a que su mayor desarrollo reproductivo ocurría cuando el área foliar alcanzaba su valor más alto. Estos mismos investigadores señalaron que ese mayor potencial productivo se debía a la gran sincronía existente entre esos dos procesos, es decir, a través del aumento del desarrollo reproductivo cuando hay un máximo peso y área foliar. Algunas características de la planta que influyen en esta sincronía de procesos son precocidad, poca altura, ramas cortas, estructura foliar eficiente en la captura de luz, y una alta tasa de crecimiento del cultivo hacia la formación de fibra (Kerby et al., 1990).
Indicadores del crecimiento, como tasa de crecimiento del cultivo (TCC), tasa de asimilación neta (TAN), relación de área foliar (RAF), área foliar específica (AFE), entre otros, son de gran utilidad para conocer como el ambiente o una práctica de manejo del cultivo afecta la eficiencia fotosintética de las plantas. Sin embargo, existen pocos estudios al respecto y en éstos se han utilizado variedades frondosas y de ciclo largo (hoy obsoletas), y que originalmente se desarrollaron para sistemas de producción en que se utilizaban surcos amplios, de 0.90 a 1 m (Mohamad et al., 1982). En la actualidad, se cultiva el algodón en el sistema de producción conocido como de surcos estrechos, donde la distancia entre surcos es de 0.76 m. Aunado a lo anterior, las nuevas variedades son de menor estructura vegetativa y de ciclo más corto que las variedades cultivadas hasta antes de 1990. Tal es el caso de la variedad transgénica NuCot 35B que posee un gen, obtenido de la bacteria Bacillus thuringiensis que le confiere resistencia genética al daño de plagas, como el gusano rosado (Pecthinophora gossypiella S.) y a las dos especies de gusano bellotero (Heliothis zea B. y Heliothis virescens). Bhatt et al. (1974) señalan que las variedades de gran masa foliar requieren de una mayor cantidad de N que las variedades precoces y de poca estructura vegetativa. Hodges (1991) confirmó los resultados anteriores y enfatizó que esto es una consecuencia de la arquitectura cónica y menor masa foliar de las nuevas variedades. El objetivo de la presente investigación fue determinar el efecto de la dosis de N en la producción y asignación de biomasa y en los indicadores del crecimiento de la variedad transgénica NuCot 35B.
MATERIALES Y MÉTODOS
El presente trabajo se llevó a cabo en 2003 en el Campo Experimental de la Universidad Autónoma Agraria Antonio Narro, Unidad Laguna, localizado en Torreón, Coah., México (25º 32' N, 103º 14' O, a una altitud de 1120 m).
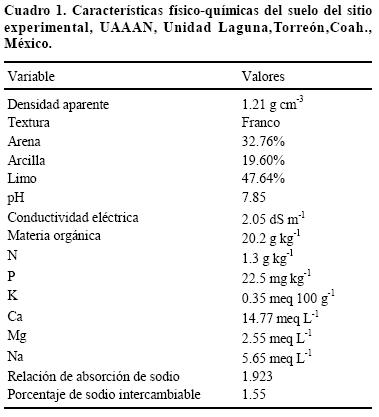
En estudios de fertilización es necesario conocer la fertilidad del suelo, con el propósito de determinar los nutrimentos limitantes y subsanar su deficiencia. Esto permite tener una mejor respuesta al nutrimento que se estudia, en este caso el N. El suelo del área experimental donde se estableció el trabajo es de textura franco limosa clasificado como Xerosol, serie coyote, medianamente alcalino (pH 7.85), con 2.02% (20.2 g kg–1) de materia orgánica y 0.13% de N total. El contenido de fósforo (P) es moderadamente alto, encontrándose a niveles de suficiencia para el cultivo del algodón con respuesta escasa o nula a la aplicación del nutrimento (Cuadro 1). El contenido de potasio (K) es bajo, como en la mayoría de los suelos calcáreos; sin embargo, su alto contenido de arena (32.76%), en comparación con la de arcillas, suministra momentáneamente más K a la solución del suelo que la arcillosa en la cual hay más fijación (Tisdale et al., 1993). El contenido de calcio (Ca) es alto y el de magnesio (Mg) es bajo (Cuadro 1). Los métodos utilizados para la determinación de N y P fueron los descritos en la NOM–021–RECNAT (2002), correspondiendo a procedimiento de digestado (AS 25) y procedimiento de Olsen (AS–10), respectivamente, y para la determinación K, Ca y Mg mediante el método AS 12, con acetato de amonio.
En la variedad transgénica NuCot 35B se evaluaron dosis de 0, 80 y 160 kg ha–1 de N se aplicó una dosis uniforme de 40 kg ha–1 de P2O5. Las dosis de N se distribuyeron en un diseño de bloques al azar con seis repeticiones. La siembra se realizó el 18 de abril en el sistema de producción de surcos estrechos, a una distancia de 0.76 m entre surcos y de 0.20 m entre plantas, para contar con una población de 65 500 plantas ha–1. Se aplicó un riego de presiembra y tres de auxilio a 60, 80 y 100 días después de la siembra (DDS). Durante el ciclo del cultivo la única plaga problema fue la conchuela (Chlorochroa ligata), la cual se controló con cuatro aplicaciones de insecticida (Cuadro 2). La maleza se controló en forma manual y química. La parcela experimental total consistió de seis surcos de 6 m de longitud y las plantas muestreadas se obtuvieron de los dos surcos centrales.

Para determinar la dinámica de producción de materia seca se realizaron cuatro muestreos destructivos, a 69, 82, 105 y 124 DDS. En cada muestreo se tomaron dos plantas con competencia completa por parcela. A cada planta se le separaron los órganos vegetativos (hojas, tallos y ramas) y los reproductivos. Para el secado de estos órganos, los tallos y las ramas, las hojas, y los frutos se colocaron en bolsas de papel por separado y se sometieron a secado en estufa marca "Felisa" a una temperatura de 65 °C durante 72 h, después de lo cual se obtuvo su peso seco; la suma de estos representó el peso seco total por planta (rendimiento biológico). Antes de someter las hojas a secado se les determinó su área foliar con un medidor de área LI–COR modelo L 1–3100. Con los valores de materia seca de láminas foliares, materia seca total, área foliar y intervalo de tiempo entre muestreos, se calcularon los siguientes índices de crecimiento, de acuerdo con Radford (1967) y Hunt (1978):
1. Tasa de crecimiento del cultivo (TCC), mide el incremento de biomasa por unidad de tiempo (g m–2 d–1):
TCC = P2 – P1 /A(t2 – t1)
donde: A = área donde el peso seco fue registrado; P1 = peso seco de muestra 1; P2 = peso seco de muestra 2; t1 = fecha de muestreo 1 (en DDS); t2 = fecha de muestreo 2 (en DDS).
2. Tasa de asimilación neta (TAN), estima la eficiencia fotosintética de la planta (g m–2 d–1):
TAN = [(PS2 – PS1) / (AF2 – AF1)] x [(Lne AF2 – Lne AF1) /(t2 – t1)]
donde: Lne = logaritmo natural, PS = peso seco de las muestras en t1 y t2; AF = área foliar en el periodo t1 y t2.
3. Relación de área foliar (RAF), indica el tamaño del aparato fotosintético de la planta y se obtiene de dividir el área foliar de la planta entre el peso seco total de la misma (cm2 g–1):
RAF = AF/PS
donde: AF = área foliar; PS = peso seco total.
4. Área foliar específica (AFE), mide el grosor de la hoja y representa la superficie foliar por gramo de peso seco de la hoja (cm2 g–1):
AFE = AF/PSAF
donde: PSAF = peso seco del área foliar
5. Relación de peso foliar (RPF), determina la distribución de asimilados hacia las hojas, y es un indicador de la frondosidad de la planta (g g–1):
RPF = PSAF/ PS
6. Índice de área foliar (IAF), es el área foliar presente por unidad de superficie de suelo (m2 m–2):
IAF = AFT/S
donde: AFT=áreafoliar total; S= área de suelo ocupada.
A partir de los datos de peso seco obtenidos en la superficie de muestreo se calcularon los datos por planta y por metro cuadrado. Para todas las variables se realizaron análisis de varianza por muestreo, así como un análisis combinado que incluyó épocas de muestreo y dosis de N. Cuando se detectaron diferencias, en la comparación de medias se utilizó la prueba de Tukey (α = 0.05) de significancia.
RESULTADOS Y DISCUSIÓN
Producción y Distribución de Biomasa
En el Cuadro 3 se presentan los resultados de la producción y distribución de biomasa del cultivo del algodón en los tratamientos de N ensayados. En ninguna fase de crecimiento del cultivo se encontraron diferencias estadísticamente significativas entre dosis de N para producción de biomasa total, ni en la cantidad asignada a los órganos vegetativos o reproductivos. Esto probablemente es una consecuencia del contenido de N total (0.13%) del sitio experimental, que se clasifica como de nivel medio. Castellanos et al. (2000) indican que el nutrimento que clasifica en esta categoría se encuentra a niveles generales de suficiencia para la mayoría de los cultivos y es de esperar que sólo los cultivos de alta demanda del nutrimento o de sistema radical limitado respondan a la adición del nutrimento. Como es natural, en las primeras etapas de desarrollo del cultivo, la acumulación de materia seca en los órganos vegetativos fue mayor que en los reproductivos. Sin embargo, conforme avanzaba el crecimiento de las plantas y aumentaba el número y tamaño de los órganos reproductivos la proporción fue invirtiéndose, lo cual coincide con lo reportado por Gaytán–Mascorro et al. (2001), en algodón, y por Sarmah et al. (1994) y Escalante–Estrada (1995) en girasol, indicando que antes del inicio de la floración, la biomasa se distribuye entre tallos, hojas y raíces, y posteriormente en la inflorescencia y los órganos reproductivos.
Índices de Crecimiento
En el Cuadro 4 se muestran los valores calculados de los índices de crecimiento del algodón (TCC, TAN, IAF) en las diferentes dosis de N probadas. Al igual que en producción de biomasa, los análisis de varianza para TCC, TAN e IAF únicamente detectaron diferencias estadísticamente significativas entre épocas de muestreo mas no entre dosis de N, lo cual implica que la cantidad de N aplicado no afectó la actividad metabólica de las plantas. Esto, como ya se indicó, es resultado de la suficiencia en este nutrimento, de acuerdo con Castellanos et al. (2000). La mayor TCC se observó entre 69 y 105 DDS para posteriormente disminuir en el período comprendido entre 105 y 124 DDS, época en que las plantas entran en senescencia después de alcanzar su máximo IAF la cual, independientemente de la cantidad de N aplicado, se obtuvo a 105 DDS. Sin importar de que se haya o no fertilizado, o de la cantidad de N aplicado, la mayor TAN se presentó entre los 69 y 82 DDS, y después de manera gradual declinó corroborando así la asociación negativa existente entre el IAF y la TAN, ya que, como una consecuencia del incremento en sombreo mutuo de las láminas foliares, a medida que aumenta el IAF disminuye la TAN.

La cantidad de N aplicado tampoco afectó a los componentes del tamaño relativo del aparato fotosintético (RAF, AFE y RPF), según se observa en el Cuadro 5. De acuerdo con los valores obtenidos para estos componentes, en todas las fases de crecimiento, las plantas acumularon la misma cantidad de fotoasimilados en sus láminas foliares, que desarrollaron la misma área foliar y presentaron el mismo grosor, de acuerdo con AFE (Cuadro 5). Sin embargo, en altura de planta, la cual se midió semanalmente, sí se observaron diferencias, ya que, en el tratamiento sin fertilizar las plantas alcanzaron su máxima altura (96 cm) a los 89 DDS, una semana antes que en los tratamientos con 80 y 160 kg ha–1 de N cuya altura promedio fue de 96 y 101 cm, respectivamente.

En todas las dosis de N, los valores más altos de RAF, RPF y AFE se obtuvieron en las primeras fases de crecimiento de las plantas y, gradualmente, declinaron conforme avanzó la edad del cultivo, lo que es normal, ya que en las primeras fases de crecimiento las plantas invierten la mayor parte de los fotoasimilados en sus estructuras vegetativas y en el desarrollo de su aparato fotosintético, sucediendo posteriormente lo contrario al establecerse la fase reproductiva, con el crecimiento en número y tamaño de órganos fructíferos (Sarmah et al., 1994; Escalante–Estrada, 1995). Los valores de RPF de las tres dosis de N fueron similares a través del desarrollo del cultivo, lo cual demuestra que la planta, independientemente de la cantidad de N aplicado, regula y distribuye equitativamente en sus órganos, los fotoasimilados que produce (Cuadro 3). Palomo y Godoy (1996) reportan resultados similares.
CONCLUSIONES
En el cultivo del algodón (variedad transgénica NuCot 35B) la dosis de nitrógeno aplicado no afectó la producción de biomasa total, su distribución a los órganos vegetativos o reproductivos, los indicadores del tamaño del aparato fotosintético, ni la eficiencia fotosintética. Los valores de relación de peso foliar obtenidos indican que, independientemente de la cantidad de nitrógeno aplicado, las plantas asignan el mismo porcentaje de fotoasimilados a su estructura foliar.
LITERATURA CITADA
Bhatt, J. G., T. Ramanujam, and E. Appukuttan. 1974. Growth and nutrient uptake in a short branch strain of cotton. Cotton Growing Rev. 51: 130–137. [ Links ]
Castellanos, J. Z, J. X. Uvalle–Bueno y A. Aguilar–Santelises. 2000. Manual de interpretación de análisis de suelos y aguas. Colección INCAPA. Instituto de Capacitación para la Productividad Agrícola. Celaya, Guanajuato, México. [ Links ]
Escalante–Estrada, J. A. 1995. Aprovechamiento del recurso agua en cultivos de secano. pp. 28–32. In: Agroproductividad. Colegio de Postgraduados. Montecillo, Estado de México. [ Links ]
Gaylor, M. J., G. A. Buchanan, F. R. Guilliland, and R. L. Davis. 1983. Interaction among a herbicide program, nitrogen fertilization tarnished plant bugs, and planting dates for yield and maturity of cotton. Agron. J. 75: 903–907. [ Links ]
Gaytán–Mascorro, A., A. Palomo–Gil y S. Godoy–Ávila. 2001. Eficiencia en la producción y distribución de biomasa en variedades precoces de algodón. Rev. Fitotec. Mex. 24: 197–202. [ Links ]
Hodges, S. 1991. Nutrient uptake by cotton. A review. pp. 938–940. In: Proc. Beltwide Cotton Conf. National Cotton Council of America. Memphis, TN, USA. [ Links ]
Hunt, R. 1978. Plant growth analysis. Studies in Biology 96. Edward Arnold. London, UK. [ Links ]
Kerby, T. A., K. G. Cassman, and M. Keerly. 1990. Genotypes and plant densities for narrow–row cotton systems. I. Height, nudes, earliness, and location of yield. Crop Sci. 30: 644–649. [ Links ]
McConnell, J. S., B. S. Frizzell, R. L. Maples, M. L.Wilkerson, and G. A. Mitchell. 1989. Relationships of irrigation methods and nitrogen fertilization rates in cotton production. Arkansas Agricultural Experimental Station. Fayetteville, AR, USA. [ Links ]
Mohamad, K. B., W. P. Sapenfield, and J. M. Poehlman. 1982. Cotton cultivars response to plant populations in a short season, narrow–row cultural system. Agron. J. 74: 619–625. [ Links ]
Norma Oficial Mexicana NOM–021–RECNAT. 2000. Diario Oficial de la Federación. Diciembre de 2002. México, D. F. [ Links ]
Palomo–Gil, A. y S. Godoy–Ávila. 1996. Análisis del crecimiento de la nueva variedad "Laguna 89" y del cultivar "Deltapine 80". Agricultura Técnica en México 22: 145–156. [ Links ]
Palomo–Gil, A., S. Godoy–Ávila y J. F. Chávez–González. 1999. Ahorro en la fertilización nitrogenada con nuevas variedades de algodón: rendimiento, componentes de rendimiento y calidad de fibra. Agrociencia 33: 451–455. [ Links ]
Palomo–Gil, A., A. Gaytán–Mascorro y S. Godoy–Ávila. 2003. Rendimiento, componentes del rendimiento y calidad de fibra del algodón en relación con la dosis de nitrógeno y la densidad poblacional. Rev. Fitotec. Mex 26: 167–171. [ Links ]
Radford, P. J. 1967. Growth analysis formulae – their use and abuse. Crop Sci. 7: 171–175. [ Links ]
Sarmah, P. C., S. K. Katyal, and A. S. Faroda. 1994. Response of sunflower (Helianthus annuus L.) cultivars to fertility level and plant population. Indian J. Agron. 39: 76–78. [ Links ]
Tisdale, S. L., W. L. Nelson, J. D. Beaton, and J. H. Havlin. 1993. Soil fertility and fertilizers. 5th ed. Macmillan. New York, NY, USA. [ Links ]
Unruh, B. L. and J. C. Silvertooth. 1996. Comparison between an Upland and Pima cotton cultivar. II. Nutrient uptake and partitioning. Agron. J. 88: 589–595. [ Links ]
Wells, R. and W. R. Meredith Jr. 1984. Comparative growth of obsolete and modern cotton cultivars. II. Reproductive dry matter partitioning. Crop Sci. 24: 863–868. [ Links ]
Wojcik, P. 2001. Ecological impact of nitrogen fertilization. J. Fruit Ornamental Plant Res. IX: 117–127. [ Links ]
* Publicado en Terra Latinoamericana 26: 29–35.

