











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Highlights:
Precipitation partitioning of Schizolobium parahyba and Samanea saman was studied.
The effective rainfall in the two species was higher in dry periods.
S. parahyba, as a smooth-barked species, had more stemflow compared to S. saman.
The higher leaf area index and rougher bark of S. saman caused less interception.
Canopy openness was correlated with precipitation variables, whereas leaf area index was not.
Introduction
The processes involved in precipitation partitioning in a forest environment are influenced by the structure and composition of the trees. When interacting with the forest, some raindrops fall through the canopy openness (throughfall), others interact with the leaves and stems of the trees (stemflow) and a part returns to the atmosphere by evaporation (interception losses) (Bessi et al., 2018; Coenders-Gerrits et al., 2020; Tonello et al., 2021a). In this way, throughfall and stemflow form a fraction of rainfall that reaches the forest floor and is known as net precipitation (Pereira et al., 2022; Tonello et al., 2021a) thus affecting the hydrological balance.
However, the results of rainfall partitioning in its three components (throughfall, stemflow and interception) can vary widely among studies of the same ecosystem (Freitas et al., 2016; Limin et al., 2015). Based on the difficulty for understanding the interaction of variables influencing these processes, these have usually been divided into those related to rainfall or meteorological events (duration and intensity) and those characterizing tree morphological traits (Tonello et al., 2021b; Zabret et al., 2018).
A little studied but relevant part of forest water balance is canopy structure (Allen et al., 2020), where tree canopies are an important variable in precipitation partitioning (Sadeghi et al., 2020; Sun et al., 2018; Tonello et al., 2021a). Especially in tropical forests, the seasonality of environmental changes between dry and rainy periods causes several responses in seedling growth and regeneration and canopy leaf composition. Leaf area index, which is the leaf tissue area per horizontal unit area ratio, is considered the major descriptor of plant structure and is related to climate modeling, productivity, health status, agrometeorology, and hydrology (Wang et al., 2018; Woodgate et al., 2015). Another relevant descriptor is canopy openness, which is defined as the proportion of clear sky seen throughout the hemisphere from a single point below the canopy, and which is related to the amount and quality of available light and, therefore, to understory ecological processes (Bessi et al., 2018; Hardwick et al., 2015; Huang et al., 2019). From an ecohydrological point of view, canopy openness can also define the amount of rainfall (and the elements added) that reaches the forest floor and will ultimately contribute to other processes such as infiltration or surface runoff (Friesen, 2020; Van Stan & Allen, 2020). In commercial reforestation projects, these processes are essential to estimate the amount of water that reaches the forest floor or evaporates (Momolli et al., 2019; Souza et al., 2019); however, there are few studies developed in the tropics that study the rainfall partitioning in these situations.
Two fast growing legumes known as tambor (Schizolobium parahyba [Vell.] S. F. Blake) and samán (Samanea saman [Jacq.] Merr.) have been exploited for different uses in tropical and subtropical America, from Mexico to southern Brazil; for example, in Brazil there is recent interest in S. parahyba for cellulose and paper, both as a pure plantation or in agroforestry systems, because the species is tolerant of low fertility and strong natural acidity of the soil (Barroso et al., 2018). S. saman pods are used in silvopastoral systems as a dietary supplement in ruminant or poultry feed, especially during dry periods. Because of its large size, S. saman provides a good percentage of shade, and is also of interest in urban tree planting because of its visual attractiveness (Vinodhini & Rajeswari, 2018).
The objective of this study was to monitor rainfall partitioning for one year in an experimental reforestation plot using S. parahyba and S. saman, and to determine how possible changes in canopy structure are related to the dynamics of precipitation interception, i.e., throughfall and stemflow.
Materials and Methods
Experimental site
The experimental area is located in La Suiza research center of the Colombian Agricultural Research Corporation (AGROSAVIA), municipality of Rionegro, department of Santander, Andean region of Colombia (7° 22’ 10” N, 73° 10’ 39” W; 550 m elevation). This region has a tropical equatorial climate (Af) and has a bimodal precipitation regime. Historical records from 1989 to 2018 indicate, on average, 27.8 ± 0.5 °C and 1 962 ± 281 mm∙yr-1 (Instituto de Hidrología, Meteorología y Estudios Ambientales [IDEAM], 2022).
Monitoring of hydrometeorological parameters
Open precipitation, throughfall also known as direct precipitation, and stemflow were monitored from February 15th 2019 to February 15th 2020 for S. parahyba (4.4-5.4 years) and from July 1st 2020 to June 30th 2021 for S. saman (5.8-6.8 years), on a 1 000 m2 plot per species.
Six-month-old seedlings grown from commercial seed were established in September 2014 at a planting density of 5 m x 5 m (400 trees∙ha-1). Variables for survival, diameter at breast height (DBH: 1.3 m above ground level), total height and at crown initiation, crown projection area, tree canopy width, and bark texture were collected from the forest inventory of the plots. Total height was calculated with a clinometer (Pm-5/360 P, Suunto) and DBH was measured with a diameter tape. When monitoring S. parahyba, incident precipitation (mm) was estimated through the mean of three 11.4 cm diameter PVC collectors at 1.3 m above the ground, according to the methodology of Ferreto et al. (2021), while for S. saman, records were taken from a mobile weather station (Watchdog 2900ET) (Figure 1A and 1C). Both measuring instruments were installed in a clear area 500 m from the experimental plot.
The throughfall was collected in the central area of each plot and under the canopy using four 10.5 cm wide x 312 cm long x 7 cm high linear PVC rain gauges, 60 cm above the ground and with a slope of 7 %. Two rain gauges were installed in the furrow position (Figure 1B) and two between the tree furrows (Figure 1D). The amount of rainfall was calculated by dividing the volume of water collected (L) by the collector area (m2).
Stemflow was quantified in three (S. parahyba) and five (S. saman) trees similar to the mean DBH value of the plot. For the selected tree, a 1 in (2.54 cm) plastic hose was installed 1.3 m above the ground spirally, cut longitudinally and sealed with silicone, rounding at least one turn of the stem (Figure 1B and 1D).
The water from stemflow was drained by the hose and stored in 15 L collectors. The volumes of stemflow (L) were divided by the projected crown area (m2) of each tree, calculated from eight directional crown radii (Tonello et al., 2014). Because of restrictions on travel to the experimental site, throughfall and stemflow were quantified simultaneously on a weekly basis to obtain the monthly accumulation; that is, a field collection could include one or several rainfall events.
Net precipitation corresponded to the sum of water runoff down the stem and throughfall in each forest formation, and monthly interception losses were the difference between throughfall and net precipitation.

Canopy structure analysis
In the same plot of the hydrometeorological monitoring and another plot located at 500 m with the same size, planting distance and age, hemispheric photographs were taken by using a fisheye lens - 180° attached to the camera of an Android 6.0 phone. The position leveled with a fixed support and defined in the central area of each plot at 1.3 m height was georeferenced with GPS; magnetic north corresponded to the top of the photograph. The activity was carried out every two weeks on a day with uniformly cloudy skies and low wind speed, avoiding exposure of sunlight to the lens.
A total of 46 high-resolution (3 264 x 2 448 megapixel) hemispheric images were totaled per species and saved in JPEG format during the 12 months of monitoring (S. parahyba = February 15th 2019 to February 15th 2020 and S. saman = July 1st 2020 to June 30th 2021). These were analyzed with the free software GLA 2.0 (Frazer et al., 1999) adjusting the regions to 16 azimuth and 9 zenith. Using the procedure of classifying the photograph between white (sky) and black (non-sky or vegetation, both foliage and wood) known as 'binarization', the same experienced user (first author) determined the best grayscale threshold for each of the hemispheric photographs. The software estimated the leaf area index (LAI, m2∙m-2) and canopy openness (CO, %). The results of the two measurements recorded per month were averaged to determine the mean monthly value.
Statistical analysis
Based on monthly records, annual precipitation distribution data for each species were distributed in a rainy period (March to May and September to November) and dry period (June to August and December to February) and compared using the parametric t-student test, once the assumptions of normality (Shapiro-Wilk test) and homoscedasticity of variances (Levene's test) were fulfilled. A linear regression analysis was carried out using the accumulated monthly data for the 12 months per species in order to identify a possible linear effect of throughfall, stemflow and net precipitation with respect to open precipitation. Finally, Pearson's correlation was used to analyze the relationship of canopy openness and monthly LAI with throughfall, stemflow, interception losses and net precipitation as a percentage of monthly open precipitation. All analyses were performed with the SAS 9.4 statistical program at 5 % significance.
Results
Table 1 shows the information collected from the forest inventory of the hydrometeorological monitoring plots of S. parahyba and S. saman. Open rainfall was similar between the two monitoring periods. The periods considered rainy (March to May and September to November) totaled 1 292 mm for S. parahyba and 1 482 mm for S. saman, 4.3 and 19.7 % higher, respectively, than the historical average for the region (Figure 2).
Table 1 Variables recorded in the hydrometeorological monitoring plot of Schizolobium parahyba and Samanea saman at 4.4 and 5.8 years of planting, respectively, in Rionegro, Santander, Colombia.
| Height category (m) | Diameter category (cm) | |
|---|---|---|
| Group 1 | Highest tenth part (35-36 m). | Highest fourth part (70-80 cm). |
| Group 2 | (i) Trees located in the tenth decile of the height category, which do not belong to the fourth quartile of diameters; (ii) trees in the fourth quartile of diameters which do not belong to the tenth decile of the height category; (iii) trees located in the ninth decile of the height category and in the third quartile of the diameter category. | |
| Group 3 | Trees that do not meet the criteria of groups 1, 2 or 4. | |
| Group 4 | Lowest tenth part (9-13 m). | Lowest fourth part (15-25 cm). |
DBH = diameter at breast height (1.3 m above the ground), ICH = initial crown height, CPA = tree crown projection area, A:AN = ratio of tree canopy height to tree canopy width. ± standard deviation of the mean.
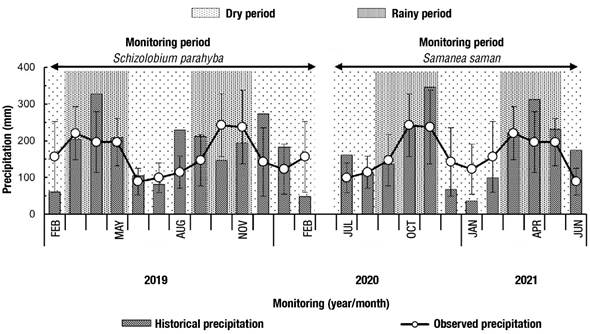
Figure 2 Monthly values of historical (1983-2018) and observed (2019-2021) precipitation during the annual monitoring period of Schizolobium parahyba and Samanea saman in Rionegro, Santander, Colombia. Variation bars represent the standard deviation of historical precipitation.
The lowest contribution to net precipitation was for S. parahyba with 78.3 % of the open precipitation, where throughfall and stemflow corresponded to 77.5 and 0.46 %, respectively (Table 2). Although 57 % of the total was concentrated in the rainy period, throughfall as a percentage of open precipitation was slightly higher (P = 0.0107) during the dry versus rainy period (79.6 % versus 76.0 %, respectively). Stemflow had no significant differences (P = 0.5738) between the two periods (46 %). Interception losses ranged from 20.5 to 24.8 %, with a higher percentage in the rainy period (P = 0.0106).
Table 2 Cumulative values of throughfall (Tf), stemflow (Sf), interception losses (I) and net precipitation (Np) in relation (%) to open precipitation (P) during annual monitoring of Schizolobium parahyba and Samanea saman in Rionegro, Santander, Colombia.
| Species | Period | P (mm) | Tf | Sf | Np | I | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| (mm) | (%) | (mm) | (%) | (mm) | (%) | (mm) | (%) | |||
| S. parahyba | rainy period | 1 292 | 982 | 76.0 b | 5.9 | 0.46 ns | 988 | 76.5 b | 304 | 23.5 a |
| dry period | 978 | 779 | 79.6 a | 4.5 | 0.46 ns | 783 | 80.0 a | 195 | 20.0 b | |
| total per year | 2 270 | 1 761 | 77.5 | 10.5 | 0.46 | 1 771 | 78.3 | 499 | 22 | |
| S. saman | rainy period | 1 482 | 1 239 | 83.6 b | 0.95 | 0.12 a | 1 240 | 83.7 b | 242 | 16.3 a |
| dry period | 652 | 568 | 87.1 a | 0.26 | 0.03 b | 568 | 87.2 a | 84 | 12.8 b | |
| total per year | 2 134 | 1 807 | 84.7 | 1.21 | 0.06 | 1 808 | 84.7 | 326 | 15.3 | |
Means followed by different letters in the columns indicate significant differences (P < 0.05) between the rainy and dry periods, according to the t-student test (P = 0.05). ns = not significant.
For S. saman, the net precipitation corresponded to 84.8 %, distributed in 84.7 % of throughfall and 0.06 % of stemflow. Between periods, throughfall had a higher contribution (P = 0.0147) during the dry period compared to the rainy period. Stemflow had the opposite behavior (P = 0.0479). Interception losses ranged from 12.8 to 16.3 %, in relation to open precipitation, with a higher contribution during the rainy period (P = 0.0106). This increase of 27.5 % was relatively stronger than that observed in S. parahyba (+17.5 %; Table 2). Statistically, differences in annual net precipitation were found between the two periods.
According to Figure 3, the two species had a strong positive and significant linear positive response of throughfall and net precipitation (R2 > 0.93) with respect to open precipitation, but the response was low in interception losses (R2 between 0.33 to 0.58). Stemflow also had a positive linear response with R2 = 0.74.
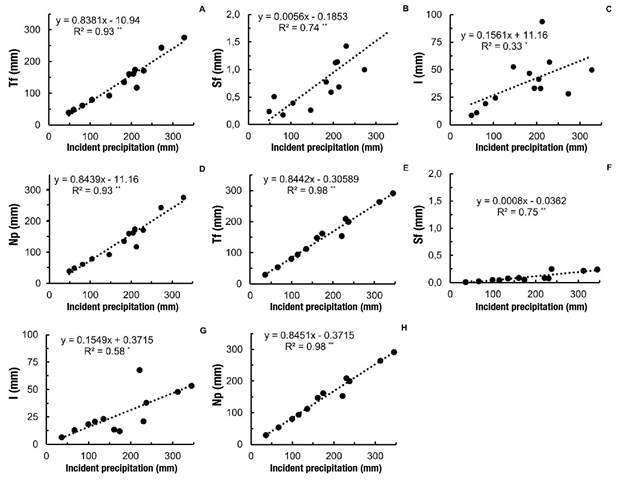
Figure 3 Linear regression between open precipitation and throughfall (Tf), stemflow (Sf), interception losses (I) and net precipitation (Np) in Schizolobium parahyba (A, B, C and D) and Samanea saman (E, F, G and H) in Rionegro, Santander, Colombia. Statistical significance: *P < 0.05 and **P < 0.01.
Canopy structure assessment
Figure 4 shows the analysis of hemispheric images with a marked monthly variation of canopy structure in both species. In the case of S. parahyba, in February 2019, canopy openness started with a low value (38 %), but gradually increased to reach a maximum in April 2019 (52.6 %). In total, four minimum and three maximum peaks occurred during the monitoring year (mean = 46.0 ± 8.9 %). The behavior of IAF was inverse; that is, at the start of monitoring in February 2019 it was a maximum (1.26 m2∙m-2; Figure 5A) dropping to a minimum in April 2019 (0.68 m2∙m-2; Figure 5B). Mean was 0.99 ± 0.3 m2∙m-2.
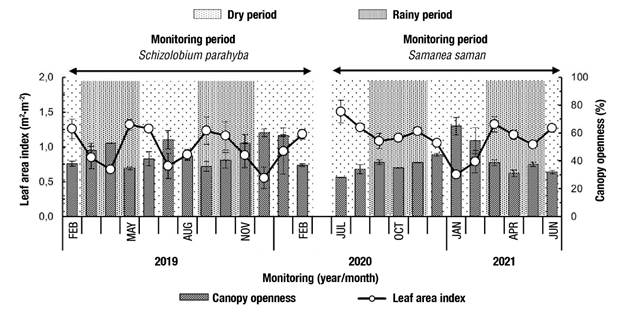
Figure 4 Monthly values of leaf area index and canopy openness during the annual monitoring period in Schizolobium parahyba and Samanea saman in Rionegro, Santander, Colombia. Variation bars represent the standard error of the mean (n = 2).
The pattern was different in S. saman because starting in July 2020, the value of canopy openness was minimal (28.2 %) and remained stable for a few months, but from December 2020 (44.5 %) it increased until reaching a single maximum peak in January 2021 (64.9 %). The mean was 39.8 ± 10.5 %. The LAI started with a maximum value in July 2020 (1.51 m2∙m-2; Figure 5C) and decreased to a minimum in January (0.60 m2∙m-2; Figure 5D). The mean was 1.51 ± 0.2 m2∙m-2. According to Table 3, only this species had a negative linear correlation between canopy openness and throughfall and open precipitation (r = -0.64, P = 0.02), and a positive linear correlation between canopy openness and interception losses (r = 0.64, P = 0.02); however, in these correlations, the linear regression analysis showed null significance and R2 values less than 0.12.
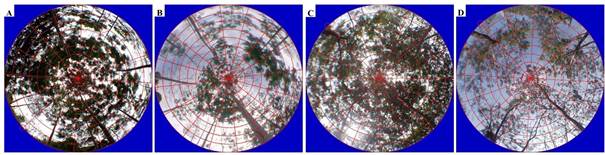
Figure 5 Processing of hemispheric images using GLA 2.0 software for annual monitoring of canopy structure in Schizolobium parahyba (maximum leaf area index in May 2020 [A] and minimum in February 2021 [B]) and Samanea saman (maximum leaf area index in July 2020 [C] and minimum in January 2021 [D]) in Rionegro, Santander, Colombia.
Table 3 Spearman correlation coefficients between precipitation variables versus leaf area index (LAI) and canopy openness (CO) of Schizolobium parahyba and Samanea saman in Rionegro, Santander, Colombia.
| Species | Canopy structure | Throughfall | Stemflow | Interception losses (mm) | Net precipitation |
|---|---|---|---|---|---|
| S. parahyba | LAI (m2∙m-2) | -0.28 (P = 0.35) | 0.13 (P = 0.68) | 0.28 (P = 0.35) | -0.28 (P = 0.35) |
| CO (%) | 0.19 (P = 0.54) | -0.18 (P = 0.56) | -0.19 (P = 0.54) | 0.19 (P = 0.54) | |
| S. saman | LAI (m2∙m-2) | 0.22 (P = 0.48) | 0.25 (P = 0.44) | -0.22 (P = 0.48) | 0.22 (P = 0.48) |
| CO (%) | -0.64 (P = 0.02) | -0.22 (P = 0.50) | 0.64 (P = 0.02) | -0.64 (P = 0.02) |
Discussion
The values of throughfall, interception losses and stemflow, as a fraction of open precipitation, varied widely among species; the results are similar to those reported by Freitas et al. (2016) and Limin et al. (2015) in tropical and subtropical deciduous forest regions, who reported values of 70 to 90 % of throughfall, 10 to 30 % of interception losses and 1 to 3 % of stemflow. In the context of the two plantations, assuming the same meteorological conditions and amount of throughfall, although the canopy of S. parahyba intercepted more rainfall, it also had greater volumes of water reaching the soil as stemflow (22.7 %, when compared to S. saman (15.3 %). These differences could have been caused, in addition to the intensity and frequency of rainfall in each monitoring period, by the nature of the stem bark and the architecture of the tree (Tonello et al., 2021b). Regarding DBH, there were no significant differences between the two species (19.4 a 21.6 cm; t = 0.2857) for the selected trees. These variables have been shown to influence stemflow among trees (Ferreto et al., 2021; Tonello et al., 2021b). This suggests that the smooth bark surface of S. parahyba, a more rectangular crown and a straight stem with branching beginning in the last third (Figure 6A and 6B) played a role in the observed differences. In contrast, the surface of S. saman has rough or scaly bark, a squarer crown, a less erect stem and the beginning of branching in the first third (Figure 6C and 6D). In the 30 most common species of the Cerrado savanna (Brazil), Tonello et al. (2021b) found that the volume of stemflow, as a proportion of open precipitation, increased when the bark surface was smooth and the canopy height-canopy width ratio was greater, i.e., trees with steeper branches.

Figure 6 Illustration of bark texture, stem straightness and initiation of branching in Schizolobium parahyba (A and B) and Samanea saman (C and D) in Rionegro, Santander, Colombia.
Stemflow is a particularly important process in anthropic environments, because it allows rainwater to be channeled to the soil with a lower velocity, which promotes infiltration and reduction of soil erosion by runoff. The difference in hydrological processes between the two species studied reaffirm that the structure of the trees defines the preferential water flow paths connecting the canopy and the deeper understory (Metzger et al., 2021).
On the other hand, although stemflow as a percentage of open precipitation was lower versus the other components in the two species (<0.5 %, Table 2), these small volumes are an important concentrated supplement of water and nutrients to the forest floor (Niemeyer et al., 2014; Tonello et al., 2021a), contaminants (Ponette-González et al., 2020), inorganic matter (Cayuela et al., 2019) and metazoans (Guidone et al., 2021; Lima et al., 2022; Ptatscheck et al., 2018). In the case of S. parahyba, a recent study noted that about 16 individuals∙L-1 of arthropods (Insecta + Collembola + Arachnida) are transported by stemflow (Lima et al., 2022). This is an important aspect, since the channels opened by these arthropods can affect water absorption and percolation rates, soil aeration, soil fauna community structure living near tree stems, and cause potential indirect effects of stemflow on this near-stem community, such as soil pH (Jozwiak et al., 2013; Kaneko & Kofuji, 2000).
In S. parahyba, Pineda-Herrera et al. (2012) found that leaf fall was negatively related to precipitation, while the opposite was true for leaf sprouting in the rainforest of Oaxaca, Mexico, characterized by seasonal rainfall. In the present study, it was found that the means of LAI (1 m2∙m-2; 0.13 a 0.9 m2∙m-2) and canopy openness (50 %; between 42 and 85 %) in S. parahyba were similar to those found during the middle of the dry period for a deciduous forest in Brazil (Freitas et al., 2016). Therefore, it becomes evident for future studies to monitor the phenology of the species.
High percentages of interception by the forest canopy are associated with high values of LAI (Ferreto et al., 2021), as occurred in S. saman, whose canopy was denser for more months compared to S. parahyba (Table 3; Figure 4). According to Kaushal et al. (2017) and Tonello et al. (2021a), wind speed and direction, phenology, leaf shape and orientation, branching pattern and angle, and other canopy properties, which were not assessed in the present study, also influence precipitation redistribution. This fact could justify the lack of correlation observed in S. parahyba. Limin et al. (2015) found no significant correlation between internal precipitation and canopy openness of Syzygium aromaticum (L.) Merr. & L. M. Perry, with very low R2 values (0.05). The same occurred in Eucalyptus benthamii Maiden & Cambage and Eucalyptus dunnii Maiden, between throughfall and LAI with an R2 less than 0.36 (Ferreto et al., 2021). The larger the tree canopy, the greater the rainfall catchment area; however, other mechanisms will influence the subsequent path of that intercepted portion.
Finally, it is important to highlight that, according to IDEAM (2015), 7.6 % (232 000 ha) of the land surface of the department of Santander presents severe (completely removed superficial soil horizons and exposed subsurface horizons) and very severe (total loss of superficial horizons and partial loss of subsurface soil horizons) erosion. Thus, studies and effective monitoring of hydrological processes, both in degraded areas and in the process of forest restoration (native or introduced) or even areas of forest overexploitation are of extreme importance and urgency, in order to support sustainable actions and soil and water conservation. This could minimize the severe stages of degradation that can also become irreversible, particularly in the department of Santander and other tropical areas of the Andean region of Colombia.
Conclusions
The rainfall partitioning of each species shows different paths in the same studied environment, especially in relation to rainfall seasonality; however, the highest net precipitation was associated with the dry period in both species. Although Samanea saman had a higher leaf area index, reforestation resulted in lower rainfall interception, higher net precipitation volume and lower stemflow than S. parahyba. On the other hand, the dynamics of rainfall partitioning, especially the processes of throughfall and interception were correlated with canopy openness only for the S. saman population. This information reinforces the importance of studying the hydrological processes in reforestation environments considering the morphological structure of the species involved, as well as the seasonality of rainfall.

