











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Cacao (Theobroma cacao L., Malvaceae) is a perennial tree grown in humid tropical parts of the world (Soto et al. 2008). Cacao beans are under constant demand; from 2007 to 2017 a 39 % increase in crop area was reported worldwide (FAOSTAT 2019). In 2017, it was reported that 5.2 million tons of cacao beans were produced around the world (FAOSTAT 2019).
The main form of cacao crop management in Latin America is through agroforestry systems (AFS) (Pacheco et al. 2011); in Mexico, several tree species are associated with cacao AFS, such as: Tabebuia rosea DC., Colubrina arborescens Sarg., Cedrela odorata L., Mangifera indica Wall., among others of the Inga, Gliricidia, Erythrina, Leucaena and Albizia genera, whose primary function is to provide shade to the cacao (Delgado 1996, Valenzuela & Alcudia 2010).
Certain criteria are considered in the selection of shade trees, such as canopy opening, phenology and use (Minorta-Cely & Ordoñez 2015, López-Baez et al. 2015). A higher incidence of disease (particularly fungal), has been reported in crop sites with greater canopy coverage and so there is a tendency towards thinning the canopy as well as reducing or eliminating shade trees (Valenzuela & Alcudia 2010, Gidoin et al. 2014, Hernández-Gómez et al. 2015). In light of this, cacao crops can be found with degrees of canopy opening from closed to open canopy, depending on the management by each producer (Carr & Lockwood 2011). The intensity of canopy management is responsible for the microclimate inside the crop area, therefore, the microclimate in an open canopy system is expected to contrast significantly with one in a closed canopy system (Babic et al. 2015, Niether et al. 2018).
Similarly, variation in the microclimate has repercussions on the physiological development of the cacao trees (Tezara et al. 2016). For example, the amount of solar radiation received by the plant determines its longevity, photosynthetic rate and leaf weight; leaves on the upper part of the canopy experience higher photosynthetic rates and lower water content and longevity, compared to the leaves in the lower canopy (Miyaji et al. 1997). The photosynthetic rate of adult cacao trees is affected by high air temperature and vapor pressure deficit (VPD) (Balasimha et al. 1991).
Furthermore, it has been reported that cacao trees tolerate up to 13 months of drought (with an 80 % reduction in rainfall) with no decrease in leaf, stem or branch growth, although bean production decreases significantly (Moser et al. 2010). Another study found that seasonality has an effect on leaf water potential in cacao, being lower in the dry season than in the rainy season (Ávila-Lovera et al. 2016). The measurement of sap flow density is a technique that helps to determine water use in plants in response to the environment (Köhler et al. 2014); different studies associate an increase in sap flow with high vapor pressure deficit, solar radiation and air temperature (Brien et al. 2004, Ortuño et al. 2006, Zhang et al. 2008, Li et al. 2016, Zhao et al. 2017). In the case of cacao trees, Köhler et al. (2010) report that sap flow diminishes after a 71 % reduction in rainfall. Similarly, Köhler et al. (2014) report that sap flow density increases in shaded trees compared to trees that grow under conditions of direct solar radiation; the authors suggest that this response is due to less solar radiation received and higher vegetative growth in trees under shaded conditions.
Canopy management has direct repercussions on the physiological development of cacao, therefore it is important to understand those effects in order to generate knowledge that contributes to the design of cacao AFS with appropriate canopy management in one of the main producing regions in Latin America. In this regard, and considering that changes in the canopy opening can influence the microclimate in the crop area and the physiological response of the cacao (Rada et al. 2005, Hasselquist et al. 2018), this work evaluated the influence of canopy management on the water content of cacao and the microclimate in the crop area, considering open canopy and closed canopy crops.
Material and methods
Study area. The study was conducted in the municipality of Comalcalco, Tabasco, Mexico, located at 18° 15' N and 93° 16' W, at an altitude of 15 m asl; the site is immersed in the main cacao producing area in Mexico. The climate is humid, warm with a total annual rainfall of 1,866 mm; March has the lowest rainfall at 49.2 mm, while July records approximately 109.5 mm of rainfall (INEGI 2018, CONAGUA 2018). Average annual temperature is 27.5 °C, maximum temperatures were 33.0 and 35.2 °C for March and July 2018, respectively (CONAGUA 2018).
Prior to starting the study, the leaf area index (LAI) was measured to select three AFS under open canopy (OC) and three crops under closed canopy (CC) management. LAI was determined using a LAI 2200C portable leaf area meter (LI-COR Inc., Lincoln, Nebraska, USA); 12 LAI points were taken (one point every 15 meters) along a 180-meter transect within each crop. Based on the above, an average was obtained and three OC AFS were selected with an LAI of 1.6 ± 0.06, and three CC AFS with an LAI of 4.2 ± 0.23.
The density of cacao trees per hectare in OC was 950 ± 150 with 63 ± 24 shade trees; both types of trees were pruned. The density of cacao trees per hectare in CC was 600 ± 100 with 50 ± 11 shade trees; none of the trees were pruned. The variety of cacao studied under both canopy conditions was Guayaquil, aged from 50 to 60 years, with heights of 4.2 ± 0.12 m under OC and 4.9 ± 0.21 m under CC, with a diameter at breast height of 12.2 ± 0.6 in trees under OC and 13.3 ± 0.5 in trees under CC. From March 13 to 19 (dry season) and July 12 to 18 (rainy season), the following measurements were taken:
Soil texture. A sample of soil was taken in five random points on each crop. The samples where mixed (a final sample for crop) and trasnported to the Plant Ecophysiology and Agroforestry Systems Laboratory (Laboratorio de Ecofisiología Vegetal y Sistemas Agroforestales, LEVSA) of the Colegio de la Frontera Sur to determine soil texture characteristics.
Microclimate. A microclimate station was installed in each crop (three under OC and three under CC), as well as a reference station placed under the open sky. Each microclimate station was composed of one HOBO U30 data logger (Onset Computer Corporation; Bourne, MA, USA), connected to sensors to record relative humidity and air temperature (RH and Ta; S-THB-M002), soil temperature (TS; S-TMB-M006) and photosynthetically active radiation (PAR; S-LWA-M003). The reference station also recorded rainfall with a rain gauge (S-RGB-M002). Each microclimate station was programmed to record data every 10 minutes for 24 hours a day, for 7 days per season of the year. The percentage of light received under the canopy was determined, using the amount of light recorded in the reference station as the maximum. The atmospheric vapor pressure deficit (VPD) was calculated based on the relative humidity and air temperature data following the approach proposed by the FAO (2006). First, the water vapor saturation pressure ( was determined using the following equation:
where T is the air temperature (°C). Next, the real vapor pressure (e a ) was calculated:
where RH is the relative humidity of the air (in percentage). Finally, the vapor pressure deficit (kPa) was calculated:
Soil water potential. Soil samples were collected at three randomly selected points in each crop; at each point samples of 150 g were collected at depths of 10, 30 and 60 cm. The samples were homogenized by depth, giving three samples per crop and a total of nine samples per canopy condition, and depth. Each sample was placed in aluminum foil and placed in a hermetically sealed plastic bag. The bags were deposited in an icebox for transportation to the LEVSA, where they were processed. The samples were taken between 05:00 and 07:30 h. The soil water potential (Ψs) was obtained using a WP4C water potential meter (Meter Group Inc. USA).
Leaf Water Potential. Four leaves per tree were collected from the middle part of the canopy of five randomly selected specimens from each crop (15 trees under OC and 15 under CC); mature leaves with a complete leaf blade were used in all cases. The sampling was taken before dawn (between 05:00 and 06:00 h) and the measurement taken immediately. The leaf water potential (Ψf) was measured using a Scholander pressure chamber (model 1505D, PMS Instrument Company, OR, USA).
Sap flow. Sap flow density (SF) was measured in 12 trees per condition, the trees being the same ones used to determine Ψf. The heat dissipation method was used with an FLGS-TDP XM1000 sap flow meter (Dynamax Inc. USA). Sensors of 50 mm in length were used at a height of 1.30 m above ground and connected to a CR1000 data-logger (Campell Scientific, INC., UT, USA). The upper sensor was fed with 250 mW, while the lower sensor was used to measure the temperature difference between the two needles. Sensors were shielded by a protection made from insulating styrofoam, covered in reflective foil that was fixed with plasticine and adhesive tape. This prevented rainwater entering the setup and limited temperature gradients caused by incident solar radiation heating the trunk. The device was programmed to take one measurement every 60 seconds, averaged per hour, for 24 h in all cases. Sap flow density was calculated following the approach proposed by Granier (1987):
where V is the sap flow velocity (cm s−1), and K was determined as:
where d indicates change, T is the current temperature and TM is the maximum temperature of the day.
where SF is the sap flow density (g cm−2 h−1), and As is the sapwood area.
The sapwood area of the cacao was determined by taking the diameter at breast height (DBH) and the trunk radius was obtained. The radius was adjusted considering the reported conductive sapwood area for cacao trees (Köhler et al. 2009). The criterion of Clearwater et al. (1999) was then applied to eliminate interference from the bark section of the trees. Based on the above, the maximum average sap flow density was calculated between 11:00 and 16:00 h. Finally, the sum total of sap flow density per day was obtained for each individual to determine daily water use (Reyes-García et al. 2012).
Cacao production. The owners of the six lots included in this work were asked to provide the average production of their cacao trees (kg of dry bean per hectare per year) during 2018, the year which the study was conducted.
Statistical analysis. A repeated measures ANOVA was carried out of the microclimatic variables: PAR, air temperature, soil temperature, relative humidity and VPD, and of the physiological parameters: sap flow and leaf water potential, taking the season of the year and canopy condition as factors. A multifactor ANOVA was carried out to determine soil water potential, taking soil depth and canopy condition as factors. A simple ANOVA was used to contrast cacao production between canopy conditions. In each analysis, an α = 0.05 and Tukey post hoc test were considered when necessary. In all cases, normality and homogeneity of variance of the data was determined prior to the analyses; where variables did not meet normality assumptions, the Kuskal-Wallis test was performed. A Pearson correlation was performed to analyze the influence of microenvironmental conditions on the water content of cacao, considering all the microclimatic and physiological variables. In all cases, the Rstudio module (Rstudio Inc. Boston MA, USA) was used in R software ver. 3.5.2 (R Core Team 2018).
Results
Characterization of the trees and soil. The height of the cacao trees was statistically different between canopy conditions: the trees under open canopy were 1.1 ± 0.1 m taller than those under closed canopy (F = 11.6; p = 0.002); however, no significant differences in DBH were found between trees in both canopy conditions (F = 1.691; p = 0.204). On the other hand, soils were of the type clay-loam with a high content of clay and silt for both open and close canopy conditions (Table 1).
Table 1 Textural characterization of soil in open and closed canopy cacao crops.
| Canopy | Clay + silt (%) | Sands (%) | Clays (%) | Silt (%) | Textural class | Code |
|---|---|---|---|---|---|---|
| Open | 66 | 34 | 28 | 38 | Clay-loam | Cr |
| Open | 66 | 34 | 30 | 36 | Clay-loam | Cr |
| Open | 66 | 34 | 30 | 36 | Clay-loam | Cr |
| Closed | 70 | 30 | 30 | 40 | Clay-loam | Cr |
| Closed | 70 | 30 | 28 | 42 | Clay-loam | Cr |
| Closed | 70 | 30 | 28 | 42 | Clay-loam | Cr |
Microclimatic variables. The received light percentage inside the crops under OC was higher than in crops under CC, in both the dry and rainy seasons (F = 113.3, p < 0.05; Table 2). No significant differences were found in the received light percentage between seasons in crops under the same canopy condition (F = 0.9, p > 0.05; Table 2).
Table 2 Light percentage calculated between 07:00 and 19:30 h during the dry and rainy seasons in crops under open and closed canopy. Data are average ± standard error. Different letters indicate significant differences (α = 0.05) between canopy conditions for the same season.
| Condition | Percentage of light received (%) | |
|---|---|---|
| Dry | Rainy | |
| Open canopy | 58 ± 12ª | 64 ± 16a |
| Closed canopy | 3.6 ± 0.1b | 4.9 ± 0.1b |
During the dry season, under both canopy conditions, PAR, VPD and air temperature reached maximum values between 12:00 and 13:00 h (Figure 1 A, B, D); in the same timeframe, the minimum values for relative humidity were recorded under both canopy conditions (Figure 1C). Maximum soil temperature values under OC were recorded between 12:00 and 13:00 h, and between 15:00 and 16:00 h under CC (Figure 1E). In the rainy season in both canopy types, PAR, VPD and air temperature reached maximum values between 12:00 and 13:00 h. On the other hand, relative humidity recorded minimum values in the said timeframe (Figure 1 F-I). The maximum values of soil temperature under OC were recorded between 12:00 and 13:00 h, and between 15:00 and 16:00 h under CC (Figure 1J).
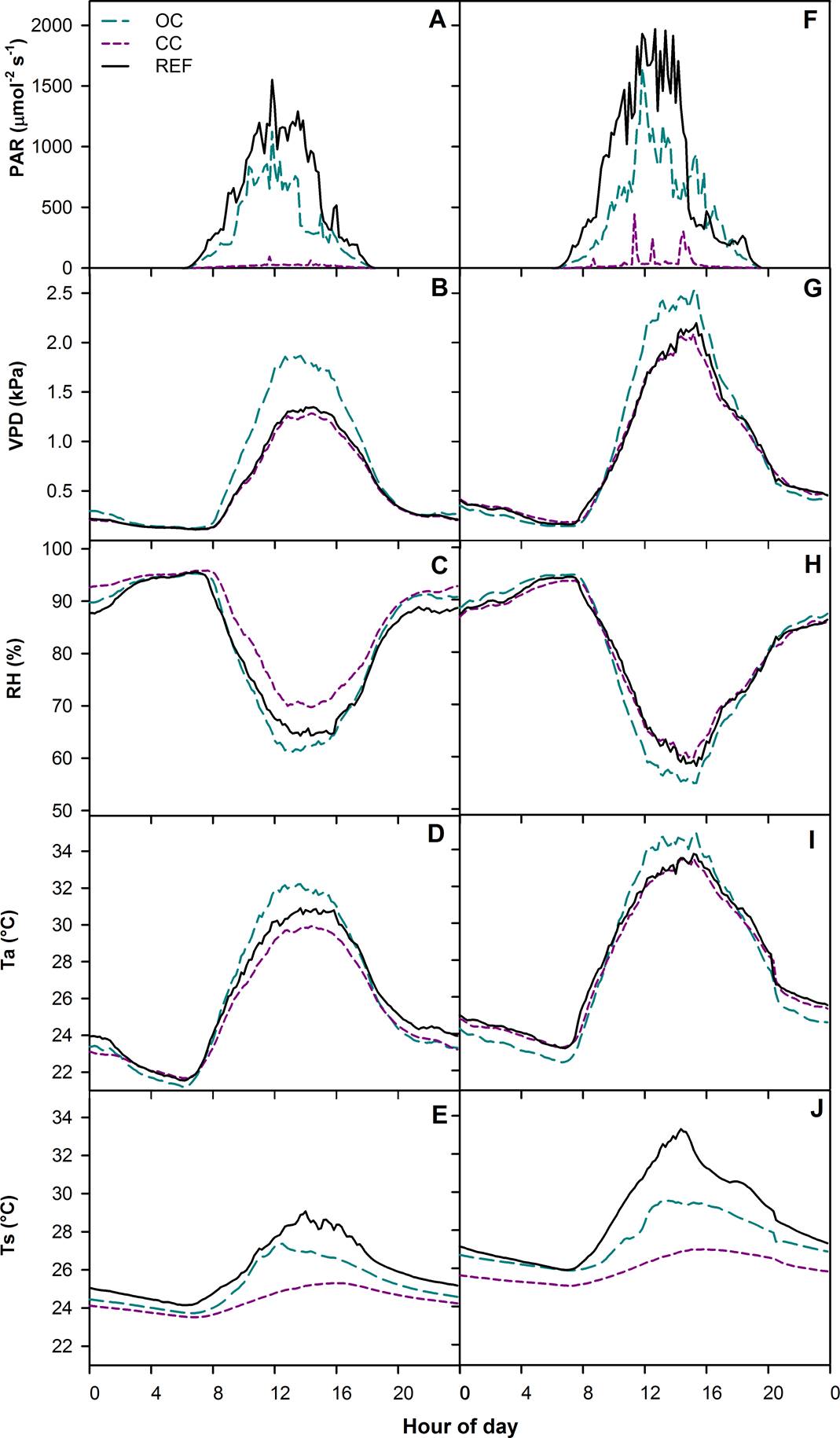
Figure 1 Microclimatic variables: photosynthetically active radiation (PAR), vapor pressure deficit (VPD), relative humidity of the air (RH), air temperature (Ta) and soil temperature (TS). Averages recorded in crops under open canopy (OC) and closed canopy (CC) during the dry season (A-E) and rainy season (F-J). REF.- Reference data.
As can be seen in Table 3, during the dry season from 11:00 to 16:00 h, microclimatic variables such as relative humidity of the air (RH), was up to 8.6 % higher in crops under CC (F = 185.6, p < 0.05), while air temperature (Ta) was up to 2.5 °C higher in crops under OC (F = 282.1, p < 0.05). VPD was up to 0.55 kPa higher in OC than in CC (F = 4.8, p < 0.05). A similar occurrence was observed in soil temperature, which was up to 2.1 ºC higher in crops under OC than in crops under CC (F = 96.8, p < 0.05). During the same timeframe in the rainy season, RH in crops under CC was 6.3 % higher than in crops under OC (F = 34.3, p < 0.05), while air temperature was 2.0 °C higher under OC (F = 41.3, p < 0.05). VPD was similar between canopy conditions (F = 12.0, p > 0.05). Soil temperature was up to 2.9 ºC higher in crops under OC (F = 310.2, p < 0.05).
Table 3 Microclimatic variables recorded between 11:00 and 16:00 h in crops under open canopy and crops under closed canopy during the dry and rainy seasons. VPD.- vapor pressure deficit, RH.- relative humidity of the air, Ta.- air temperature and TS.- soil temperature. Values are averages ± standard error. Different letters indicate significant differences between conditions in the same season (α = 0.05).
| Microclimatic variables | Season | Open Canopy | Closed Canopy | Reference |
|---|---|---|---|---|
|
VPD (kPa) |
Dry | 1.8 ± 0.07a | 1.2 ± 0.03b | 1.5 ± 0.2 |
| Rainy | 2.4 ± 0.1a | 2 ± 0.05b | 2 ± 0.1 | |
|
RH (%) |
Dry | 61.7 ± 0.8b | 70.3 ± 0.7a | 65 ± 4 |
| Rainy | 56.7 ± 0.6b | 63 ± 1a | 62 ± 1.9 | |
|
Ta (°C) |
Dry | 32 ± 0.2a | 29.6 ± 0.1b | 30.5 ± 1 |
| Rainy | 34.6 ± 1.2a | 32.6 ± 0.2b | 32.6 ± 0.1 | |
|
Ts (°C) |
Dry | 27 ± 0.8a | 24.9 ± 0.09b | 28.3 ± 0.8 |
| Rainy | 29.4 ± 0.8a | 26.5 ± 0.05b | 32.2 ± 0.2 |
No significant differences in soil water potential (Ψs; Table 4) were recorded between OC and CC in the dry season (F = 0.2, p > 0.05), or the rainy season (F = 0.04, p > 0.05) at all depths. Comparing the two seasons, it was observed that Ψs was lower in the rainy season, under both canopy conditions (F = 13.5, p < 0.05). Similarly, significant differences were recorded in Ψs between 10 cm and 60 cm depth in soils under CC during the dry season (F = 5.4, p < 0.05), while no significant differences were recorded between depths under OC in any season (F = 0.02, p > 0.05).
Table 4 Soil water potential (Ψs) in cacao AFS under open canopy (OC) and under closed canopy (CC). Values are averages ± standard error. Different letters indicate significant differences between soil depths (α = 0.05) for the same canopy condition and in the same season.
| Season | Depth (cm) | OC (MPa) | CC (MPa) |
|---|---|---|---|
| Dry | 10 | -0.31 ± 0.04a | -0.33 ± 0.04a |
| 30 | -0.29 ± 0.01a | -0.22 ± 0.04a | |
| 60 | -0.31 ± 0.08a | -0.11 ± 0.05b | |
| Rainy | 10 | -0.76 ± 0.37a | -0.80 ± 0.27a |
| 30 | -0.62 ± 0.32a | -0.81 ± 0.30a | |
| 60 | -0.51 ± 0.36a | -0.91 ± 0.22a |
Sap flow, daily water use and leaf water potential. Flow density presented differences between trees under OC and CC, in both seasons of the year. In the dry season, trees under OC recorded a maximum average sap flow density of 17.71 ± 1.28 g m-2 h-1, while trees under CC recorded 20.88 ± 0.60 g m-2 h-1 (F = 4.79, p < 0.05, Figure 2A). In the rainy season (Figure 2B), trees under OC recorded a maximum average sap flow density of 14.99 ± 0.52 g m-2 h-1, while trees under CC recorded 21.69 ± 1.95 g m-2 h-1 (F = 10.99, p < 0.05). However, no significant differences were recorded between seasons (Figure 2; F = 0.591, p = 0.45). Daily water use in the dry season was significantly lower (F = 1.7, p > 0.05) in trees under OC (11.4 ± 2 kg d-1) than in trees under CC (17.2 ± 3.9 kg d-1). The same behavior was observed in the rainy season: daily water use in trees under OC was 12.21 ± 0.1 kg d-1 and 20 ± 0.6 kg d-1 in trees under CC (F = 30, p < 0.05). Leaf water potential recorded in the dry season in OC conditions was -0.38 ± 0.03 MPa and in CC conditions -0.45 ± 0.03 MPa, with no significant differences between canopy conditions (F = 3.5, p > 0.05). In the rainy season, leaf water potential was -0.37 ± 0.04 MPa under OC and -0.48 ± 0.01 MPa under CC. No differences were recorded between seasons in trees under both OC and CC (F = 0.2, p > 0.05).
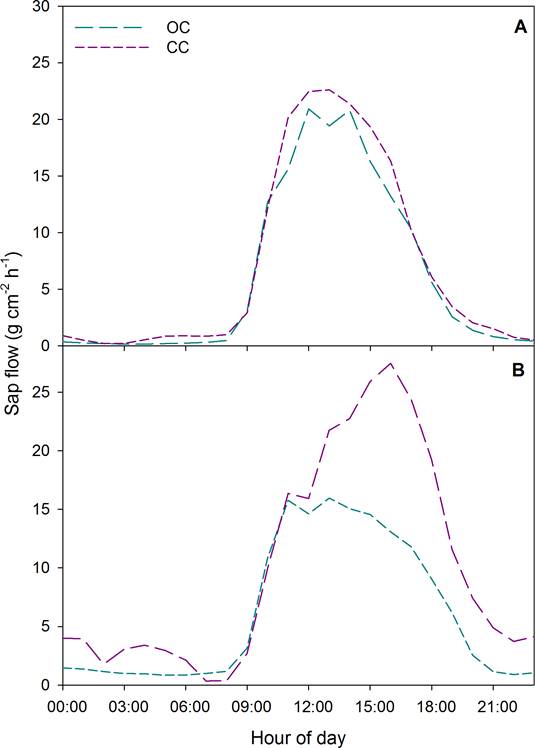
Figure 2 Sap flow density (SF) in cacao trees under open canopy (OC) and under closed canopy (CC) during the dry season (A) and rainy season (B). The data represented are an average of the measurements taken of 12 trees per canopy condition (closed and open), per season.
Correlation of hydric state-microclimate. In the dry season, sap flow density presented a significantly positive correlation with VPD (r = 0.8), Ta (r = 0.7) and TS (r = 0.6), while leaf water potential presented a significantly positive correlation with Ta (r = 0.6) and TS (r = 0.6) and a significatively negative correlation with RH (r = -0.6). In the rainy season, sap flow density presented no correlation whatsoever with microenvironmental variables; leaf water potential showed a direct correlation with PAR (r = 0.8) and TS (r = 0.8).
Cacao production. According to the information provided by the producers of the six lots included in this work, cacao dry bean production is approximately 2,633.34 ± 133.34 kg/ha/year-1 in open canopy, while in closed canopy is 1,133.34 ± 185.59 kg/ha/year-1 (F = 43.09; p = 0.0028).
Discussion
The crops under open canopy recorded higher photosynthetically active radiation, air temperature, soil temperature, vapor pressure deficit and relative humidity in both seasons of the year. The results are similar to those reported in cacao agroforestry systems in Brazil, where the decrease in tree cover increases the incidence of radiation, temperature and vapor pressure deficit (Niether et al. 2018), while in crops with a 2.2 % canopy opening, i.e., greater canopy coverage, a decrease in air temperature and soil temperature is reported compared to crops with a 7.7 % canopy opening (Pinheiro et al. 2013).
Nevertheless, canopy management did not affect water availability in the soil, since the water potential at a depth of 10 cm presented no differences between canopy conditions in the two seasons of the year; this behavior is contrary to that reported in agroforestry systems with greater canopy coverage which recorded less water loss, above all in the first layers of soil (Lin 2007). Our results may be related to the soil characteristics at the work site. The soils in the work plots were found to be clay-loam (Table 1); these soils are characterized by a moderately firm texture and a medium to high water retention capacity (Rucks et al. 2004). This water retention capacity helps to explain the similarity in water potential in the most superficial layer of soil between canopy conditions. In addition, the work region is located in the inactive fluvial delta plain, which contains soils formed by alluvial deposits that do not currently receive sediment discharge and whose main characteristic is the proximity of the phreatic mantle to the surface, which helps to maintain water availability in the soil throughout the year (Palma-López et al. 2016, Zavala-Cruz et al. 2016).
On the other hand, the soils under closed canopy recorded significant differences in water potential between 10 and 60 cm depth in the dry season, with higher potential at a greater depth, while in open canopy soils the water potential was similar between depths in both seasons of the year. These results may be related to the density of the cacao trees, where, contrary to what might be expected, a lower tree density per hectare can produce a difference in soil water potential at different depths. This phenomenon has been observed in other agroforestry systems; for example, in coffee plantations Bermúdez-Florez et al. (2018), report differences in water content throughout the soil profile when the planting density is 5,000 plants per hectare, while in plantations of 10,000 plants per hectare the water content was similar at different soil depths. In our case, the closed canopy systems had 600 cacao trees per hectare, while the open canopy systems had 950 trees per hectare. Tree density can affect soil hydraulic properties by increasing water demand (Bermúdez-Florez et al. 2018), which may explain the difference found in our work.
Contrary to what was expected, a lower soil water potential was recorded in the rainy season, in both canopy conditions. It has been reported that in fluvial soils rainfall increases the volumetric soil water content (Geris et al. 2017), however, our results report the opposite. The difference in soil water potential between seasons may be related to the physiological response of the cacao. It has been shown that trees increase water extraction from the soil with the increase in air temperature (Zweifel et al. 2007), as long as the water available in the soil is not a limiting factor. In our case, increases in air and soil temperatures, relative humidity and vapor pressure deficit were recorded during the rainy season, causing the cacao trees to extract a greater amount of water from the soil, as corroborated by our sap flow data. The above causes a reduction in soil water potential, mainly under closed canopy conditions.
Contrary to that reported by Ávila-Lovera et al. (2016), who found lower leaf water potential in cacao trees during the dry season (−0.41 ± 0.05 MPa) compared to the rainy season (−0.21 ± 0.02 MPa), in this work we found that canopy management did not affect leaf water potential values of the cacao trees growing under both canopy conditions nor between seasons, suggesting similar hydric behavior throughout the year. It is possible that the cacao trees have had access to a permanent water source throughout the year, due to the presence in the area of a phreatic mantle close to the surface; during sampling, it was possible to observe water at little more than 60 cm deep in some cacao plantations. It has been reported that the decrease in volumetric water content in the soil has a negative effect on leaf water potential: in cacao plants without irrigation (during 60 days), leaf water potentials from -2.0 to -2.5 MPa were reported (Santos et al. 2014, García & Moreno 2016), while in this work, the lowest leaf water potential was -0.48 ± 0.01 MPa in trees under CC during the rainy season (although not significantly different from the trees under OC). Although it would be expected for soil and leaf water potential values to be similar since they were taken before dawn (Nobel 2009), the access of the deep roots of the cacao to the phreatic mantle could contribute to the leaves presenting a less negative water potential than the superficial layers of the soil. Therefore, the availability and distribution of water in the soil layers appears to play a more important role in leaf water content in cacao trees than other microclimatic changes produced by contrasting canopy management.
Sap flow density was modified by canopy management, since higher values were recorded in closed canopy trees compared to open canopy trees. This behavior is similar to that reported by Köhler et al. (2014), where the cacao crop with lower light incidence recorded higher sap flow than trees with direct radiation. Moreover, several studies have shown that the increase in photosynthetically active radiation, vapor pressure deficit, and air and soil temperature increase the density of sap flow (Brien et al. 2004, Zhang et al. 2008, Chang et al. 2014, Zhao et al. 2017). However, our results show the opposite, since in the open canopy crops with higher values of the aforementioned microclimatic variables, sap flow density was lower compared to closed canopy crops. This result may be due to a high physiological plasticity reported in cacao to grow under direct solar radiation through greater control of the stomatic opening, which reduces water loss by transpiration and increases water use efficiency (Almeida et al. 2018). This mechanism may lead to a reduction in sap flow, as found in this work.
Water use was recorded at 20 ± 0.6 kg day-1 in trees under closed canopy, higher than that found by Köhler et al. (2010) who reported 10.6 kg day-1 in cacao trees growing under a canopy of Gliricidia. This difference in water use may be due to the density of trees per hectare and the difference in diameter at breast height: the higher the DBH, the higher the percentage of conductive area (Köhler et al. 2014). In the works of Köhler et al. (2010), DBH was 10 cm, while the trees studied have a DBH of up to 17 cm, which may suggest a greater conductive area.
The production data reported by the producers match previous reports which indicate that in humid regions, cacao production is lower in closed canopy agroforestry systems than intermediate canopy systems (Abdulai et al. 2018). In our case, the open canopy classification does not imply the elimination of shade trees, nor does it correspond to the cacao plantations with greatest canopy opening found in this region of Mexico; the leaf area index recorded for these cacao plantations (4.2 ± 0.23) suggests that shade trees intercept between 35 % and 40 %, approximately, of the total light incidence on the crop area, and therefore continue to play an important role in the management scheme.
In conclusion, despite the fact that canopy management significantly alters microclimatic variables, there was no significant change in the water content of cacao trees between open and closed canopy conditions, the only exception being sap flow, which was higher in trees under closed canopy compared to open canopy. Therefore, crops under open canopy (according to the LAI characterization done in this work) can function as a good management scheme, since it is possible to have a greater density of cacao trees per hectare, and in turn, promote microenvironmental conditions less favorable to fungal diseases, which may significantly increase cacao bean production, as reported by the producers at the work site. In addition, this management system only implies greater canopy management of the shade trees, not their elimination from the crop area, and therefore, like closed canopy cacao plantations, they contribute significantly to the conservation of biodiversity in the region. Complementary studies are needed to help understand, for example, the photosynthetic behavior of cacao trees in order to have a complete panorama of the physiological performance under both management schemes.

