











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Plant-plant associational effects are common and frequently documented in plant communities, but the mechanisms are not always evident (Barbosa et al. 2009). Abiotic stressful conditions in dry environments usually pose a significant threat to plant survival, growth, and reproduction by reducing the intensity of competition for resources among plant species (Callaway & Walker 1997). However, species associations through facilitation are common since they can attenuate the negative effects of stressful conditions in these environments (Callaway 1995, Flores & Jurado 2003, Brooker et al. 2008, Bonanomi et al. 2011). Facilitation is a type of commensalism whereby some species function as providers of improved abiotic and biotic microenvironmental conditions without obtaining any benefit (host or nurse species) and other species take advantage of these conditions (focal or commensal species), increasing their establishment (Holmgren et al. 1997), reproduction (Valiente-Banuet et al. 1991), and survival (Smith & Huston 1989). This positive neighbor effect of nurse species on focal species can result from direct facilitation through a reduction in stressful physical conditions (Shreve 1931, Valiente-Banuet & Ezcurra 1991) or from indirect facilitation through an increase in marginal benefits for the focal species (Sosa & Fleming 2002, Tálamo et al. 2015). However, this biotic interaction may attract herbivores that could counteract the benefits of protective plants. The magnitude of indirect herbivory damage on focal species can be reduced by density- and/or frequency-dependent neighbor effects (associational effects) as well as by the density and/or frequency of the focal species’ own defensive genotypes (Tahvanainen & Root 1972, Agrawal et al. 2006, Barbosa et al. 2009, Underwood et al. 2014, Kim & Underwood 2015, Tálamo et al. 2015, Garrido et al. 2016). In other cases, associational effects have been observed to increase susceptibility to damage (Underwood et al. 2014, Tálamo et al. 2015, Garrido et al. 2016). Even though there are reports of positive and negative effects of facilitation on focal species, there is still a significant gap in the empirical evidence that does not allow the inference of general patterns (reviewed in Barbosa et al. 2009 and Underwood et al. 2014).
Species benefited by facilitation can be associated with only one or several different nurse species (Bonanomi et al. 2011), and the overall benefits of direct and indirect facilitation may also vary according to the taxonomic identity of the nurse species (Callaway 1998, Castillo-Landero & Valiente-Banuet 2010). Even though indirect facilitation through reduced damage has been observed in several cases (Bonanomi et al. 2011), how this effect varies with the identity of nurse species is less understood. If increased resistance results from the deterrent effects of specific nurse species traits against natural enemies of focal species, taxonomically distinct nurse species should provide different levels of associational resistance (Barbosa et al. 2009) to focal species. On the other hand, if a similar level of indirect protection is provided by different nurse species, other mechanisms acting at the neighbor level should be involved, such as density, frequency, and diversity of neighbor species within patches (Underwood et al. 2014). An improvement in microclimatic conditions within patches where facilitation occurs can increase plant tissue quality and attract a diversity of insect and vertebrate species that would feed on the plants in these patches (Sosa & Fleming 2002). Thus, resource-rich patches resulting from the effect of nurse species may represent a risky habitat for focal species independently of the taxonomic identity of the nurse species.
The Cactaceae family has ≈ 200 genera and 2,500 species, mostly adapted to xeric conditions, native to the American continent (Bravo-Hollis 1937), and commonly associated with nurse species (Flores & Jurado 2003). The giant columnar cactus Pilosocereus leucocephalus is one of the main components of food chains in Mexican tropical dry forests, since it provides nectar, pulp, seeds (Miranda-Jácome et al. 2020, Vázquez-Castillo et al. 2019, Miranda-Jácome & Flores 2018, respectively), and sites for animal reproduction (Miranda-Jácome et al. 2020, 2021). This species exhibits a significant spatial association with woody species, which results in increased germination, survival, and growth during establishment, suggesting a benefit due to the attenuation of solar irradiance (Miranda-Jácome et al. 2013). The distribution of diametric size frequencies of P. palmeri (synonym of P. leucocephalus) has been fitted to a log-normal curve, and thus it is unimodal and indicates that there is constant recruitment in the population (Lezama-Delgado et al. 2020). In addition, P. leucocephalus is regularly bitten by one or two species of opossums (Didelphis marsupialis, Caluromys derbianus).
In this study, we measured herbivory damage on the columnar cactus P. leucocephalus (focal species) in open spaces (3 × 3 m spaces without canopy of woody plants and with direct solar radiation) and mediated by the presence of seven species of shrubs, as well as the size of the cactus. Herbivory was measured through the observation of the number of scars from bites of small vertebrate species on the stem of the cacti. We hypothesized that the outcome of the interaction would be regulated by the characteristics of the interacting species and that herbivores would counteract the benefits of the protective plants.
In addition, herbivory intensity (total number of scars) would be predicted by cactus size, since damage probability would increase with the tissue or area exposed to herbivores.
Materials and methods
Species and study site. P. leucocephalus (Cactaceae) is a hermaphroditic, self-incompatible, bat-pollinated giant cactus species (up to 3 m tall) with a distribution from Guatemala to the northeastern arid areas of Mexico (Guzmán et al. 2003, Munguía-Rosas et al. 2009). It exhibits second- and third-order branches starting almost from the base, ≈ 15 erect or ascending stems, and a diameter of 6-10 cm without leaves. The stems have 7-10 bare ribs with spines only on the edge; with a single 2-3 cm long central spine and 8-12 slender 1-2 cm long radial spines.
In the State of Veracruz (Mexico), this species is present in dry tropical forests from 50 to 1,200 m asl (Bravo-Hollis 1937). Within the studied region in Central Veracruz, this is the only species that can survive above 1,100 m asl, whereas at lower elevations, it coexists with other columnar cactus species (Neobuxbaumia scoparia and Stenocereus griseus; Medina Abreo & Castillo-Campos 1993). The studied population, San Ignacio (19 ° 35 ´ 26 ´´ N, 96 ° 58 ´ 38 ´´ W), is located on the uppermost limit of the distribution range of the species, at 1,150 m asl, in a lava flow substrate deposited during the Holocene 10,000 years ago (Negendank et al. 1985). P. leucocephalus individuals are more frequently distributed under the canopy of woody species such as Casearia corymbosa Kunth, Cnidoscolus aconitifolius (Mill.) I.M. Johnst., Lysiloma acapulcense (Kunth) Benth., Quercus oleoides Schlcht & Cham., Fraxinus schiedeana Schlcht & Cham., and Zanthoxylum fagara (L.) Sarg. Seedlings and juvenile individuals are also found under adult plants of P. leucocephalus, which could come from seeds dispersed by gravity from the mother plant, and juvenile adults are also found in open spaces without vegetation (Miranda-Jácome & Flores 2018). The average temperature in the area is 20.4 ° C, with May being the hottest month and January the coldest. The annual precipitation is 1,300.0 mm, 79.1 % of which is concentrated between May and October (Fernández-Eguiarte et al. 2012, period 1903-2010). Even though the annual precipitation corresponds to that expected for a tropical dry forest and not a desert community, there is limited soil development in the lava flow substrate half of the soil surface is exposed (data taken from 5 × 50 m transects, not shown), which prevents the retention of humidity from rainfall. The dominant tree species are Dodonaea viscosa, Quercus oleoides, Cnidoscolus aconitifolius and Tonduzia longifolia (Miranda-Jácome et al. 2013).
In the study area, the stems of the columnar cactus P. leucocephalus suffer regular bites (5 × 3 × 3 cm) between 1.20-2.30 m from the ground, which are likely caused by one or two Didelphimorph species (Didelphis marsupialis, Linnaeus, 1758; Caluromys derbianus (Waterhause 1841)). Bites can be easily identified as brown scars that remain on the cactus for several years, since the damaged surface is not restored.
Field work. From April to May 2019, we randomly selected 46 P. leucocephalus plants within an area of 3 ha growing underneath the canopy of C. corymbosa (n = 5), D. viscosa (8), F. schiedeana (7), L. acapulcense (9), P. rubra (5), Q. oleoides (9), and Z. fagara (3), and 14 P. leucocephalus plants growing ≥ 3 m away from other species and exposed to full sunlight conditions (N = 60). We obtained the size (height of main stem from ground to apex) of the focal P. leucocephalus plants in each microenvironment and determined the intensity (total number of bites on the cactus stem) of vertebrate herbivory.
In order to characterize the environmental temperature and luminosity in spaces with and without canopy, both variables were estimated every hour between 01:00-00:59 h on a clear day in May 2022, we measured daily temperature (°C) and luminosity (lum/ ft2) using four HOBO Pendant Temperature/Light 64K Data Loggers (Onset Computer Corporation, Bourne, Massachusetts) with thermal shields. Two data loggers were placed at ground level in two open spaces separated by 500 m. The other two data loggers were placed at the same distance from the ground in two sites with trees.
Data analyses. An ANOVA was used to evaluate the relation between the size (previously transformed to hierarchical ranks (see Conover & Iman 1981)) of P. leucocephalus and the eight microenvironments (open spaces and underneath the canopy of the seven nurse plants) by estimating the significant differences in the mean size of P. leucocephalus individuals in the eight microenvironments. A repeated measures ANOVA was performed to compare the daily temperature (°C) and luminosity (lum/ ft2) records between the shaded microenvironment and the open space.
An analysis of covariance was performed to determine the effect of the microenvironment (open spaces, shade of C. corymbosa, D. viscosa, F. schiedeana, L. acapulcense, P. rubra, Q. oleoides, and Z. fagara) and the size of P. leucocephalus individuals on the number of bite scars on the stem (Crawley 2007). After the analysis of covariance, multiple comparisons between nurse species and open spaces were done using a least significant difference test and a Bonferroni correction. A linear regression model was fitted to number of bite scars and plant size. To improve the normality of the data and satisfy the criteria of normality, homogeneity of variances, and independence of errors of the statistical models, number of bite scars was square-root transformed. All statistical analyses were performed using R version 4.02 (R Core Team 2020) and the package agricolae (Rice 1989).
Results
The mean height of the P. leucocephalus individuals in the eight microenvironments only showed significant differences between the individuals established under the canopy of L. acapulcense and Q. oleoides (mean 296.67 ± 26.69 vs. 188.55 ± 24.30 SE, respectively) (Table 1, Figure 1). There was also a significant effect of time and of the interaction time × environment on the daily temperature and luminosity, where the values of both variables were higher in the open space environment (Table 2, Figure 2a, b). The number of bites on the stems of P. leucocephalus cacti established in open spaces was significantly higher (mean 36.64 ± 1.27 SE) compared to cacti growing underneath nurse plants (8.02 ± 1.13 SE) (Figure 3, Table 3). In the last microenvironment, the number of bites on the stems of P. leucocephalus significantly depended on the nurse plant species, with a mean number of bites fluctuating between 16.67, underneath L. acapulcense, and 3.80, underneath C. corymbosa. Cactus size was a determining factor in herbivory damage intensity, since number of bites increased significantly (P < 0.0001) with plant size, both in open spaces and under the shade of nurse plants (Table 4, Figure 4).
Table 1 Anova of size of P. leucocephalus in relation to the eight microenvironments (open spaces, underneath the canopy of the seven nurse plants). The data were previously transformed to hierarchical ranks (Conover & Iman 1981).
| Source of variation | df | SS | CM | F | P |
|---|---|---|---|---|---|
| Microenvironment (M) | 7 | 5,214.9 | 744.99 | 3.041 | 0.0093 |
| Error | 52 | 12,740.6 | 245.01 |
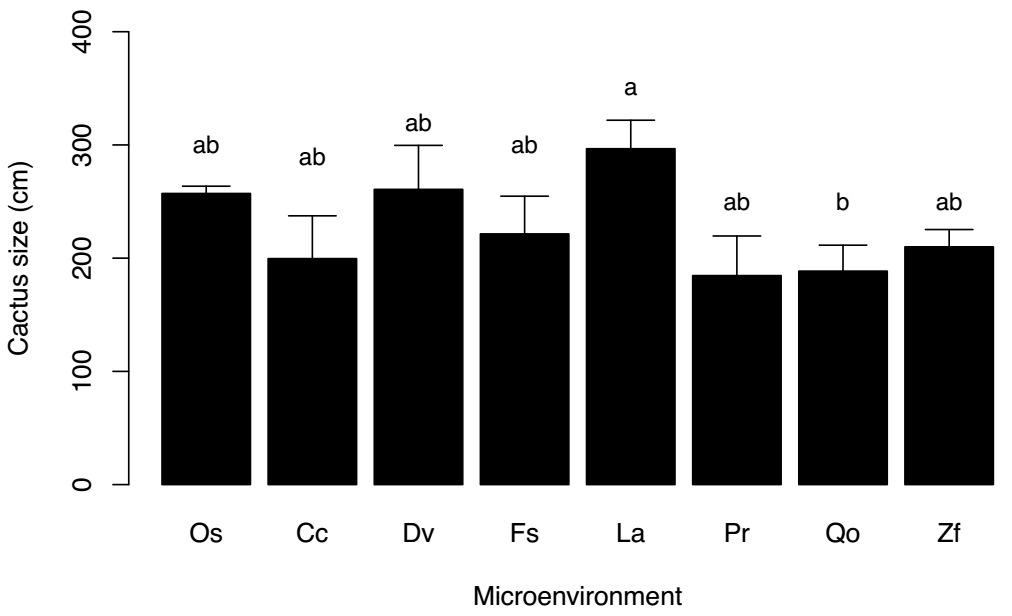
Figure 1 Mean (± SE) size of P. leucocephalus individuals established in the eight microenvironments (open spaces, underneath the canopy of seven nurse plants). Casearia corymbosa (Cc), Dodonaea viscosa (Dv), Fraxinus schiedeana (Fs), Lysiloma acapulcense (La), Plumeria rubra (Pr), Quercus oleoides (Qo), and Zanthoxylum fagara (Zf). Different letters above bars denote significant differences (P < 0.05).
Table 2 Analysis of variance of the daily temperature (°C) in two contrasting and representative environments (open spaces, underneath plant canopy) where individuals of P. leucocephalus are established.
| Source of variation | df | SS | CM | F | P |
|---|---|---|---|---|---|
| Microenvironment (M) | 1 | 14,352 | 14,352 | 47.57 | ≤ 0.0001 |
| Time (T) | 23 | 508,847 | 22,124 | 73.33 | ≤ 0.0001 |
| M × T | 23 | 14,711 | 640 | 2.12 | 0.0041 |
| Error | 143 | 43,143 | 302 |
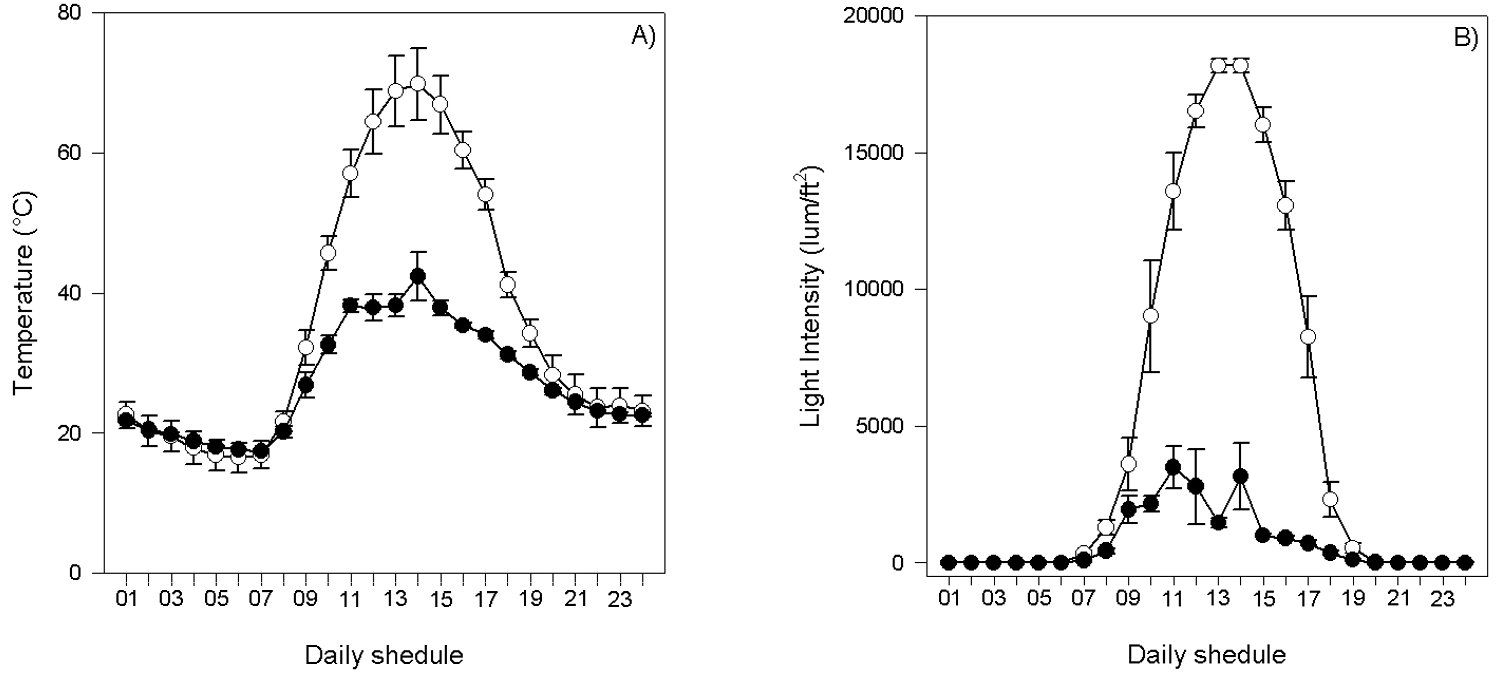
Figure 2 Mean (± SE) daily temperature (°C) (A) and luminosity (lum/ ft2) (B) in two contrasting and representative environments (open spaces, underneath plant canopy) where individuals of P. leucocephalus are naturally established. Open spaces (OS) and underneath plant canopy (PC).
Table 3 Analysis of variance of the daily luminosity (lum/ ft2) in two contrasting and representative environments (open spaces, underneath plant canopy) where individuals of P. leucocephalus are established.
| Source of variation | df | SS | CM | F | P |
|---|---|---|---|---|---|
| Microenvironment (M) | 1 | 10,311 | 10,311 | 83.92 | ≤ 0.0001 |
| Time (T) | 23 | 496,314 | 21,579 | 175.64 | ≤ 0.0001 |
| M × T | 23 | 28,251 | 1,228 | 89.99 | ≤ 0.0001 |
| Error | 143 | 17,568 | 123 |
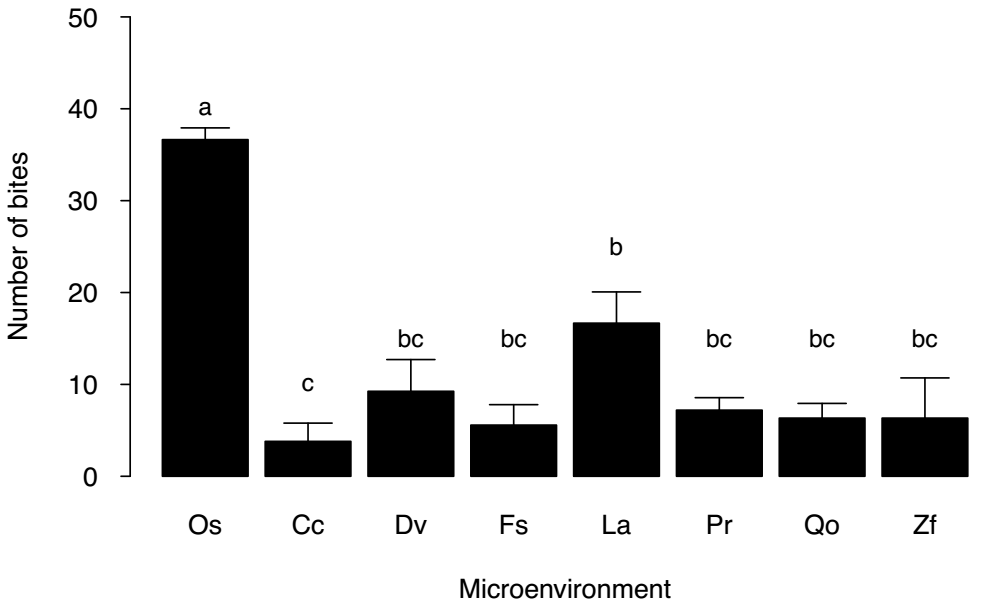
Figure 3 Mean (± 1 SE) number of bites on the stem of individuals of the columnar cactus Pilosocereus leucocephalus established in eight microenvironments: open spaces and underneath the canopy of Casearia corymbosa (Cc), Dodonaea viscosa (Dv), Fraxinus schiedeana (Fs), Lysiloma acapulcense (La), Plumeria rubra (Pr), Quercus oleoides (Qo), and Zanthoxylum fagara (Zf). Different letters above bars denote significant differences (P < 0.05).
Table 4 Analysis of covariance of the occurrence of bites on the stem of P. leucocephalus in relation to the microenvironment (open spaces, underneath the canopy of seven nurse plants) and cactus size. The occurrence of bites on the cacti was square-root transformed.
| Source of variation | df | SS | CM | F | P |
|---|---|---|---|---|---|
| Microenvironment (M) | 7 | 153.9 | 21.99 | 22.49 | < 0.0001 |
| Cactus size (S) | 1 | 12.5 | 12.51 | 12.80 | 0.0008 |
| M × S | 7 | 13.3 | 1.90 | 1.94 | 0.0850 |
| Error | 44 | 43.0 | 0.98 | ||
| Total | 59 | 222.78 |

Discussion
Our results indicate a beneficial effect of nurse plants on herbivory, since individuals of P. leucocephalus under the canopy of nurse plants showed a significantly lower number of bites in general than individuals established in open spaces.
Columnar cactus size is determined by intrinsic factors such as the age and physiology of the plant, as well as extrinsic factors such as resource availability (Steenbergh & Lowe 1977, 1983). For example, a positive correlation between age and size has been observed in Pachycereus pringlei and Carnegiea gigantea (Delgado-Fernández et al. 2016, Drezner 2003, respectively), which are columnar cacti of the same subfamily to which P. leucocephalus belongs, Cactoideae. However, the authors mention that there could be significant errors in the estimation of age using height if there is no model that allows to correct the prediction by the factor of site. A comparative study between P. leucocephalus and Cephalocereus scoparius showed a higher growth exponent in height in the former than in the latter species, possibly due to the allocation of more biomass to vertical development during the early stages of growth (Lezama-Delgado et al. 2022). The authors mention that the hydraulic capacity that allows the transport of water through the stems could also determine the height of the plants. We did not estimate the age of the plants, but the individuals established in the sampled microenvironments were of similar size and thus had a similar probability of being attacked by herbivores.
Our results contribute with new field evidence indicating that the beneficial effect of nurse plants of reduced herbivory damage may also depend on the nurse species identity. This beneficial effect of plant-plant facilitation dependent on nurse species identity (Castillo-Landero & Valiente-Banuet 2010) could promote non-random patterns of plant-plant spatial associations. These patterns would be related to the specific effects of nurse plants on the physical microenvironmental conditions, which would reduce the stress of focal species, the predation of seedlings, and the increase in selective dispersal (Sosa & Fleming 2002). Associational damage, defined as the probability of being attacked depending on the neighboring species, is a common phenomenon (Barbosa et al. 2009, Underwood et al. 2014) and has been recently demonstrated to occur among genotypes within species (Garrido et al. 2016). This form of plant-herbivore interaction can condition the cost of a nurse plant for a focal species by reducing or increasing the benefits of facilitation.
Our results indicate that the beneficial effect of nurse plants was reduced when the cacti were associated with more palatable species, such as L. acapulcense (Pennington & Sarukhan 1998), which allowed more understory radiation and higher herbivory levels compared to C. corymbosa (Farji-Brener 2001).
However, if more palatable species attract large herbivores, this may favor an increase in the deposition of feces underneath the canopy of these species, which would increase soil nitrogen, and nitrogen-fixing species, such as L. acapulcense, can also increase the soil nitrogen available to other species (García-Moya & Mckell 1970, Virginia & Jarrell 1983, Pugnaire et al. 1996). In turn, this positive effect on soil nutrient conditions can reduce the costs of herbivory damage and/or facilitate cactus growth. Thus, the specific effect of nurse species on the community of vertebrate herbivores is expected to condition the optimal protection of focal species.
The occurrence of plant-plant facilitation has been initially related to stressful environments where the benefits of being spatially aggregated outweigh the costs of negative interactions (Callaway 1995, Callaway & Walker 1997). It is possible that, after establishing, benign microenvironmental conditions underneath the nurse plant canopy will continue and allow cacti to maintain a reduced cost due to lower herbivory damage resulting from denser vegetation.
In our study site, the temperature and light conditions were significantly different between the environments without vegetation and under the canopy of a nurse plant, with the highest values of both variables in the spaces without vegetation cover. Therefore, the beneficial abiotic conditions determined by lower temperature and light intensity and the biotic conditions determined by lower herbivory result in an important role of nurse plants in the establishment and survival of P. leucocephalus.
There are many studies on the negative and positive effects of herbivory on plant growth (leaves, shoots, stems, and root tissues) and reproduction (buds, flowers, fruits, and seeds). In the first case, herbivory represents a selective pressure for the evolution of plant defenses (Mattson & Addy 1975, Morrow & LaMarche 1978, Whitham & Mopper 1985). In the second case, plants may benefit from being eaten by responding by overcompensating and ultimately achieving higher fitness (Vickery 1972, Hilbert et al. 1981, Inouye 1982). Herbivory has been studied in natural conditions by field observations or experimentally in greenhouses (Britton 1988, Baldwin 1990, Hatcher et al. 1995, Agrawal 1998, Tiffin & Inouye 2000, Lehtilä & Boalt 2008), and in different herbivore guilds, such as invertebrates (Bowers & Puttick 1988, Després et al. 2007, Wiens et al. 2015, War et al. 2018) and vertebrates (Choat & Clements 1998, Iason & Van Wieren 1999, Hans-Dieter 2000, Wynne-Edwards 2001). However, there is little evidence on how plant size could deter or attract herbivores of certain vertebrate species. In this study, we predicted a positive relationship between herbivory intensity (number of bites) and plant size because there would be an increase in the tissue or area exposed to herbivores. The results supported this hypothesis, since herbivory (number of bites) on the stem increased with the size of the central plant. This could be explained by an increased inter-rib distance in the stem resulting in less disturbance to the feeding behavior of herbivores caused by the spines of the cactus. This inhibitory effect of spines on herbivore feeding has been demonstrated in species of ungulates foraging on plant species with spines, with the effect being much lower on small species (impalas and goats) than on large ones (kudus). The spines could therefore limit the loss of tissue caused by large herbivores (Cooper & Owen-Smith 1986). However, it is not known if increasing the volume of the stem (height) exposes areas free of thorns in P. leucocephalus, making them more susceptible to bites, an important fact to study in future research. While there is recent evidence on the incidence of columnar cacti damaged by insect herbivores and pathogens (Bravo-Avilez et al. 2014), herbivory by large vertebrates in columnar cacti has been poorly studied (Bravo-Avilez et al. 2019). Therefore, this is the first report of herbivory in columnar cacti caused by vertebrates considering the size of the focal plant.
The null interaction between microenvironment and plant size suggests that herbivores visit and consume in both environments larger plants non-randomly with increased light exposure (open spaces), followed by nurse species that allow more light radiation in the understory (L. acapulcense). The above could represent two scenarios, with the first involving a triple plant-environment-herbivore interaction, where the herbivore could obtain a higher quantity of food and of better quality by consuming larger stems exposed to higher light radiation (abiotic factor), which would be reflected in the content of nutrients in the diet (e.g. carbohydrates, sugars) due to increased photosynthesis rates produced by the light environment and the greater amount of exposed tissue.
It is known that abiotic factors significantly participate in chemical variations (e.g., carbon/nutrient balance, ethanol, etc.) of natural plant populations and this causes differential susceptibility of individuals to natural enemies (herbivores) (Sudatti et al. 2011). For example, temperature affects the physiological state in the response of secondary metabolites (Connan et al. 2007), while light intensity contradictorily affects the terpenoid compounds of plant populations (Cronin & Hay 1996). This represents a trade-off for individuals in their vital processes, including anti-herbivore chemical defenses. In fact, this production, transport, and storage of defensive complexes (e.g., secondary metabolites) requires the allocation of abiotic resources (e.g., light), which implies a cost for the organism (Cronin 2001).
Light intensity or PAR (photosynthetically active radiation) levels have been shown to influence plant responses to herbivores, where increased light is positively correlated with an increased production of anti-herbivore volatiles in Zea mays (Gouinguené & Turlings 2002, Frenkel et al. 2009), higher densities of defensive foliar trichomes and associated allelochemicals in tomatoes against the caterpillar Manduca sexta (Kennedy et al. 1981), and the induction of other leaf secondary metabolites such as flavonoids and phenolic acids (see Escobar-Bravo et al. 2017), which has even been tested in macroalgae (Pavia & Toth 2008). However, it is not known if abiotic variation can affect the production of defense metabolites or the palatability of columnar cacti. Our results suggest that the abiotic component (open spaces with high irradiance vs. underneath the canopy of nurse plants) and cactus size allow greater rewards for herbivores, which is reflected in the number of bites, possibly due to the palatability or nutritional content of the plants in open spaces or underneath the canopy of nurse species that allow more radiation (e.g., L. acapulcense).
The second scenario involves a triple plant-herbivore-predator interaction, where the predator is attracted by the plant volatiles induced by the herbivore, which can function as environmental signals that ensure the presence of the prey (see Clavijo McCormick 2016). It is known that predator-prey abundance (herbivore-predator in the specific case of this study) is determined by the abiotic environment, which can generate variation in the intensity (e.g., moderate vs. strong) and direction (positive vs. negative) of biotic interactions. For example, a light environment (open meadows vs. shaded understory) has been observed to increase ant protection and result in a higher abundance of Aphis heleanthi, which feeds on the grass Ligusticum porter, where the abiotic factor favored the ant-aphid mutualism against ladybugs (Coccinelidae), fly larvae (Syrphidae), Lygus bugs (Miridae), and red spider mites (Acaridae) (Mooney et al. 2016). This demonstrates that the abiotic environment can shape the predator-prey cycle (see Wilmers et al. 2007) in a specific case of herbivory (see Bale et al. 2002). However, little is known about the interaction cactus-vertebrate herbivore and its natural enemies, since it is not known whether the location of the cactus (e.g., open spaces vs. under the canopy of some nurse species) could represent an unsafe site for the herbivore with respect to its natural enemies. The individuals of P. leucocephalus located under the canopy of nurse species showed the lowest number of bites, and thus this could be an unsafe site for herbivores with respect to their natural enemies, since nocturnal raptors could use the branches of nurse plants as perching and stalking sites and nocturnal mammals could use these plants for camouflage and burrows.
Finally, our study suggests that the plant-plant interaction had a negative effect on the cactus-herbivore interaction, increasing the benefits of facilitation. This pattern may result from a higher predation risk for the herbivore inside than outside the vegetation. Given that most of the feeding activity of the two Didelphimorph species in the study site occurs at night, the open habitat may provide a less risky microenvironment. Furthermore, we showed that larger individuals in spaces without vegetation showed increased herbivory, possibly due to an increase in the quality and quantity of the reward for the herbivore. In the first case, due to a larger inter-rib separation in the stem (where spines are formed), and in the second case, due to a higher concentration of nutrients resulting from higher radiation.

