











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introduction
Maize (Zea mays L.) is the most important crop in Mexico and is used for human and livestock feeding (Ureta et al. 2020). This cereal is the most consumed by Mexicans with 297 kg per capita per year, nevertheless, the country faces an approximate deficit of 8.3 million tons per year, which is satisfied with the imports (Ayvar-Serna et al. 2020). Mexico is the second-highest maize importer in the world. Over the past 10 years, the average maize yield in Mexico has been 2.9 t ha-1 (Ureta et al. 2020). In the Yucatan Peninsula maize yield is less than 1 t ha-1 (SIAP 2017). The maize-cropped area in Yucatan was 107 566 ha in 2017, with a total grain yield of 92 564 t mainly produced by small farmers with less than 5 ha during the rainy season (SIAP 2017). Some of these farmers still grow maize in a traditional agroecosystem named milpa, where several species are grown simultaneously. These farmers are preserving maize diversity in situ and it should be made possible for them to satisfy their household consumption needs with their maize production (Dzib-Aguilar et al. 2016, Ureta et al. 2020). Usually, small-scale farmers sow more than one type of maize, including landraces, commercial varieties, and hybrids (Fenzi et al. 2015). Landraces are commonly grown in smaller areas with low cropping technology and, therefore, have lower production costs and yield. Nevertheless, landraces are rustic and adapted to the adverse soil and climate conditions and biotic agents which are frequent in the agroecosystems (De Araujo et al. 2013).
On the other hand, commercial varieties and hybrids are significantly superior to the native landraces in terms of performance (Fernández et al. 2013). During the last decades, several improved maize genotypes have been released for the tropic region of Mexico. However, this innovation has a slow adoption process in part due to the high cost of the technology package (Uzcanga et al. 2017). In Yucatan, approximately 78% of the area cultivated with maize is planted with landraces and 22% with commercial genotypes (Dzib-Aguilar et al. 2011). The area dedicated to each type of maize depends on many factors such as the type and amount of seed available, seed preference, soil type, among others (Fenzi et al. 2015, Uzcanga et al. 2017).
In Yucatan, agriculture practice is difficult and presents many challenges (Fonteyne et al. 2023). More than 80% of soil corresponds to the unit Leptosol, characterized by shallow soils with a scarce amount of land and a large amount of stone or rocky outcrops (Bautista-Zúñiga et al. 2002). Fenzi et al. (2015), noted that farmers typically opt for sown improved maize varieties and hybrids in the local soil known as kankabal (Cambisol), characterized by its deep, reddish-black color considered superior. Nevertheless, this soil type typically constitutes a small percentage of the total cultivated areas for maize (Fenzi et al. 2015). The climate is warm-subhumid with summer rains (García 2004). Precipitation during the growing season arrives primarily in the form of convection thunderstorms, which sweep across the Yucatan Peninsula in temporally and spatially unpredictable patterns (Tuxill et al. 2010). Under these specific agroecological conditions, the decision to choose an optimal sowing date and to select maize genotypes with both a short growth cycle and productive characteristics proves challenging on an annual basis (Aguilar-Carpio et al. 2015). The study of crop behavior and production potential of maize genotypes grown in different areas provides valuable information to assist farmers and technicians in making cropping decisions, especially because it helps to identify limiting production factors (De Araujo et al. 2013). According to Fonteyne (2023) is keenly important the study about of potential of maize productivity and forms of intensification to tropical agricultural development. This knowledge is crucial for maximizing agricultural productivity across the diverse range of soils within the agroecosystem. In this way is important to explore the type of maize genotypes that should be planted in each agroecosystem, and the most appropriate aptitude for better use of its genetic potential (Aguilar-Carpio et al. 2015). Therefore, this study aimed to investigate the ecophysiological attributes related to the growth and yield determinants of six maize genotypes under the soil and climate conditions prevalent in Yucatan.
Materials and methods
Field experiments and treatments
The field experiment was conducted at the National Technological Institute of Mexico, Conkal campus in Yucatan (21° 04' 36" N y 89° 29' 60" W), during the rainy season from July to October 2017. The sowing date was July 26, 2017. Local weather conditions during the field experiment were Tm 26.7 °C, Tmax 33.6 °C Tmin 19.7 °C, and mean monthly precipitation of 124 mm (CONAGUA 2017). The soil was classified as Leptosol with 0.93% N, and the total content of P, K, Ca, and Mg were 2.45, 3.50, 49.38, and 2.63 g kg-1, respectively. Six treatments consisting of three maize landraces and three commercial genotypes were evaluated. The landraces are locally named Elotillo, Chimbo, and San Pableño, and were collected from farmers in the states of Yucatan and Chiapas in southern Mexico. The specific collected locations and agronomic and physiologic information on these landraces have been published elsewhere (Dos Santos et al. 2019). The commercial genotypes are highly cultivated in the region and correspond to VS-536 (INIFAP, Mexico), hybrid Dekalb 390 (Monsanto, Illinois, USA), and Impacto (Syngenta, Basel, Switzerland).
Experimental design
The experiment was arranged in a randomized complete block design, with six replicates to mitigate the effect of soil heterogeneity. Plants were distributed in rows 1.2 m wide. Two seeds were sown at 0.3 m space within each row. Each useful plot comprised a row with a length of 10 meters, accommodating a total of 66 plants. The extrapolated population density was 55 000 plants ha-1. The plants were fertilized with a dosage of 120-80-80 (kg ha-1 of N-P-K, respectively), applied in the band and a fractional manner; half of N and all P and K at three weeks after emergence, followed by the remaining half of nitrogen two weeks later. Weeds were manually controlled. There were no applications of insecticides or fungicides during crop growth.
Response variables
Response variables were the days to silking (DS, days) determined when 50% of the ears in a plot attained silking. Then, vegetative characteristics of plant height (PH, cm), measured from the base of the plant to the tip of the tassel, the total number of leaves (TNL, units), and leaf area (LA, cm2) per plant were taken in five plants randomly of each experimental unit (30 repetitions). Leaf area was measured with a leaf-area meter (LI-COR 3000C, Lincoln, NE, USA). To evaluate the growth rate of maize genotypes, destructive samplings of four plants in each experimental unit (24 repetitions) were made at 15, 30, 45, and, 60 days after sowing (das). Dry matter (DM, g m-2) was determined after the samples were dried at 70 °C in a forced air circulation oven until constant weight. The leaf area index (LAI) is the product of LA and the plant density per m-2 (Lukeba et al. 2013).
With previews data, the crop growth rate (CGR, g m-2 day-1) and net assimilation rate (NAR, g m-2 day-1) were calculated using the equations:
where: DM2 and DM1 indicate dry weight of the aerial plant per unit area gained at time T1 and T2 respectively.
where: DM2 and DM1 express the dry matter weight of the plant, ln LAI2 and ln LAI1, natural logarithm of the leaf area index at time T2 and T1, respectively (Aguilar-Carpio et al. 2017).
At physiological maturity, twelve plants per treatment were carefully extracted from the soil. After rinsing the roots, plants were cut and separated by tissues (roots, stems (including leaf sheath), leaves, and, tassel. The ear was separated into husk leaves, grain, and cob. Samples were dried in an oven at 70 °C to a constant weight and plant dry matter (DM, g plant-1) was calculated.
Grain yield (GY, kg ha-1), was estimated with the cob weight harvested per experimental unit, adjusted to 14% moisture, multiplied by the shelling rate. The harvest index (HI) was calculated by dividing the weight of maize grains by the total weight of the plants.
Data analysis
The analysis of variance (ANOVA) was performed after checking data for normality and homoscedasticity. Means were grouped using the Scott Knott multiple comparison test (p < 0.05) (Jelihovschi et al. 2014). The average correlation among the vegetative traits was obtained using Pearson correlation coefficients (rxy). All analyses were performed in Infostat software, version 2020 (Di Rienzo et al. 2020).
Results and discussion
Vegetative characteristics
The ANOVA showed significant differences among maize genotypes in all traits evaluated (Table 1). These results indicated that farmers in Yucatan have available maize genotypes with different vegetative characteristics. The variable DS ranges from 49 days in the landrace Chimbo to 56 days in the landrace San Pableño. The three commercial genotypes took approximately 52 days to silk, at least three days before Elotillo and San Pableño. All maize genotypes were categorized as short-cycle maize genotypes. Precocity is an important factor that offers the possibility of completing flowering and ear maturation before the canicula (periodic short-term interaestival drought conditions) and denotes adaptation to the agroecosystem (Tuxill et al. 2010). In the work conducted by Fenzi et al. (2015) in Yaxcaba, Yucatan, the authors reported a farmer’s trend for sowing shorter-cycle maize genotypes as an additional way to ensure production in the context of increased climate uncertainty. Environmental factors such as recent changes in rainfall regimes or hurricanes influence farmers’ decision-making to change the milpa planting months that have moved from May/June to June/July (Fenzi et al. 2015). In the study conducted by Dzib-Aguilar et al. (2011) with other short-cycle maize genotypes in Yucatan, maize genotypes took a mean of 71 days to silk, larger than what was observed here. The observed variation was attributed to the fact that maize genotypes interact differently with environmental conditions, emphasizing the need for quantifying distinct experimental conditions.
Table 1 Vegetative characteristics (mean ± ES) evaluated in six tropical maize genotypes in Yucatan, Mexico.
| Genotype | Type | DS (days) | PH (cm) | TNL | LA (cm2) |
|---|---|---|---|---|---|
| Elotillo | Landrace | 55.3 ± 0.3a | 166.0 ± 2.9c | 14.0 ± 0.2b | 5 032 ± 226b |
| Chimbo | Landrace | 49.0 ± 0.4c | 226.5 ± 3.3a | 13.4 ± 0.2c | 5 410 ± 202b |
| San Pableño | Landrace | 56.0 ± 0.5a | 233.2 ± 4.8a | 15.1 ± 0.2a | 6 873 ± 388a |
| VS-536 | Improved | 52.2 ± 0.5b | 210.7 ± 3.8b | 14.5 ± 0.2a | 6 842 ± 330a |
| DK-390 | Improved | 52.8 ± 0.2b | 208.9 ± 4.4b | 15.0 ± 0.2a | 6 877 ± 326a |
| Impacto | Improved | 51.3 ± 0.3b | 209.1 ± 3.8b | 14.3 ± 0.2b | 5 897 ± 337b |
| MSD | 1.65 | 15.66 | 0.78 | 1245 | |
| CV (%) | 1.78 | 10.16 | 7.39 | 24.54 |
DS: days to silking; PH: plant height; TNL: total number of leaves; LA: leaf area per plant; Distinct letters in the same column indicate significant differences (Scott-Knott, P ≤ 0.05); n = 30. MSD: minimum significant difference; CV: Coefficient of variation.
For PH, Elotillo had shorter plants with 166 cm, followed by the improved genotypes VS-536, DK-390, and Impacto with 211, 209, and 209 cm heights, respectively. Maize landraces San Pableño and Chimbo were 10% taller than the improved genotypes. Typically, taller plants exhibit a prolonged maturation cycle, and their height is considered undesirable due to increased vulnerability to wind-induced stress and this susceptibility often leads to lodging (Rodríguez et al. 2016). Nevertheless, Chimbo and San Pableño were 14% and 11% shorter than the local landrace Nalxoy with 259 cm, evaluated by Dzib-Aguilar et al. (2011) in Yucatan. In TNL, maize genotypes ranged from 13 leaves in the landrace Chimbo to 15 leaves, in San Pableño, DK-390, and VS-536. Likewise, LA ranged from 5 032 cm2 in Elotillo to 6 877 cm2 in DK-390, statistically similar to San Pableño and VS-536 with 6 873 and 6 842 cm2, respectively (Table 1). The traits TNL and LA were positively related and genotypes with high amounts of leaves (VS-536, DK-390, and San Pableño) also had high LA (Table 1). Castro-Nava et al. (2014) assessed different maize genotypes, both commercial and native, in Tamaulipas, Mexico. They reported a mean leaf area of 4 731 cm2 in commercial genotypes and 4 906 cm2 in native maize genotypes, which is lower than that observed in maize landraces from Yucatan.
Dry matter accumulation and maize growth rates
During the first 45 das, DM accumulation was low, and mean values of DM were 3.9, 72.1, and 156.4 g m-2 at 15, 30, and 45 das, respectively (Figura 1a). At 60 das, values of DM ranged from 600.4 g m-2 in the landrace Elotillo to 861.2 g m-2 in the DK-390. The landrace Elotillo had accumulated approximately 27% less DM than the other genotypes at 60 das. Similarly, the CGR was relatively low during the first 45 das with a mean of 0.3, 4.5 and, 5.6 g m-2 d-1 at 15, 30 and, 45 das respectively (Figure 1b). At 60 das, the CGR highly increased in all genotypes to a maximum of 45.5 g m-2 d-1 in the landrace Chimbo. At the same age, CGR observed in Elotillo was lower with 33.9 g m-2 d-1. Maize net assimilation rate (NAR), which is an estimator of photosynthetic activity, increased during the first 30 d in all genotypes (Fig. 1c). At 30 das, values of NAR ranged from 23.0 g m-2 d-1 in the Elotillo to 47.5 g m-2 d-1 in the DK-390. Between 30 to 45 das, the NAR remained relatively stable with a mean value of 38.7 g m-2 d-1 at 45 das. Then, values of NAR increased again in all genotypes from 75.2 g m-2 d-1 in the Elotillo to 109.5 g m-2 d-1 in the landrace Chimbo with a mean of 92.5 g m-2 d-1 (Figura 1c)
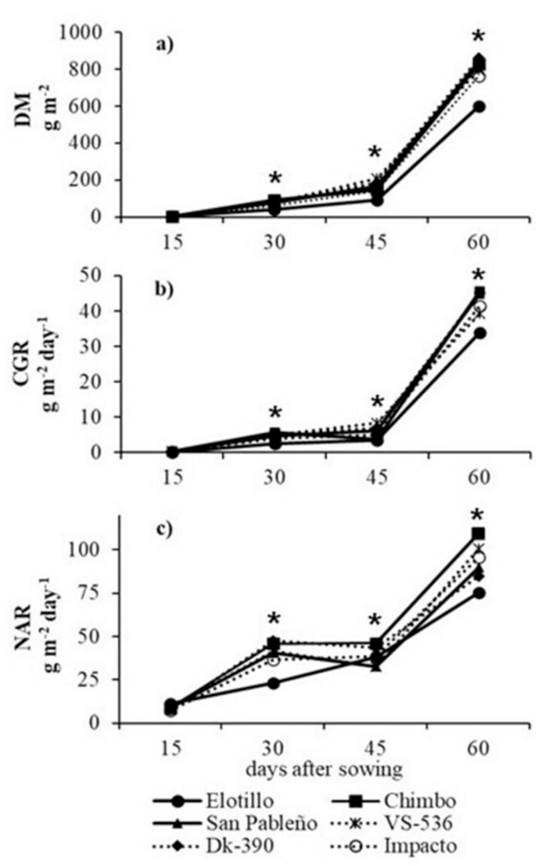
Figure 1 a) Dry matter accumulation (DM), b) Crop growth rate (CGR), and c) Net assimilation rate (NAR) of six tropical maize genotypes in Yucatan, Mexico. * Significant differences between genotypes (Tukey, P ≤ 0.05).
The patterns of crop growth varied between maize genotypes. Usually, DM increases to a maximum point and maintains stable until the end of the growing season (Heggenstaller et al. 2009). The CGR dynamics generally increase to a maximum point near silking and subsequently decline throughout the remainder of the growing season as a result of the cessation of vegetative growth, loss of leaves, and leaf senescence and NAR begins at a low level, increases to a maximum point relatively early in growth, and then decline as result from the onset of light limitation, as increases in LAI beyond an optimum level that began to shade lower leaves in the canopy, reducing overall photosynthetic efficiency (Heggenstaller et al. 2009, Tajul et al. 2013). Leaf area and leaf photosynthetic efficiency represent two fundamental factors driving crop growth (Heggenstaller et al. 2009). The three commercial genotypes exhibited uniformity in plant growth; however, the landraces displayed heterogeneous plant growth. The growth performance of all maize genotypes evaluated suggested that these genotypes were more efficient in the production of DM after 45 das. Also, genotypes with a longer growth cycle showed higher accumulated dry matter and CGR. At a pre-silking stage, genotypes highly increase the LA and consequently the photosynthetic capacity. This is because the increased leaf area provides more surface for the functioning of the photosynthetic apparatus, contributing to a greater overall efficiency in photosynthesis. Most genotypes exhibited superior CGR performance compared to the landrace Elotillo, reaching maximum values that exceeded 40 g m-2 day-1 post-silking stage. Liu et al. 2023, mentioned that higher proportions of pre-silking growth duration significantly increased total, stem, and leaf dry matter transfer rates while post-silking proportions had the opposite effect. The research highlights the importance of an extended growth duration, especially post-silking, for achieving higher grain yields. Also, have been shown that CGR is influenced by nitrogen (N) fertilization (Aguilar-Carpio et al. 2015, Aguilar-Carpio et al. 2017). Aguilar-Carpio et al. (2015) mentioned that N application increases photosynthetic capacity and efficiency in the production of dry matter during maize development. In this study, plants were fertilized in two applications with 50% of N at 23 das and the rest of N at 36 das. Therefore, the rapid increase of CGR after 45 das might be attributed to fertilization, nevertheless, this hypothesis was not investigated here. The NAR indicates the amount of biomass accumulated in terms of leaf area and time (Díaz-López et al. 2013). Also, the NAR is indirectly related to light penetration in the canopy (Tajul et al. 2013). In this study, NAR showed an increasing trend until 60 das due to an acceleration in the photosynthetic activity. Particularly in the landrace Chimbo, the plants were taller and had a narrow shape of the leaves which allowed received sunlight longer, which may have a positive effect in the NAR at 60 das.
Dry matter allocation, grain yield, and harvest index
The DM showed significant differences between genotypes and ranged from 141.3 g plant-1 in the landrace Elotillo to 258.4 g plant-1 in the hybrid DK-390, with a mean of 230 g plant-1. Maize genotypes DK-390, San Pableño, VS-536, Impacto, and Chimbo were statistically similar and superior to Elotillo (Figure 2a). In the study conducted by Robles-Jimenez et al. (2021), to determine the forage yield of maize landraces in Mexico authors reported an average of 279.1 g plant-1 which is higher than the dry matter observed in our study. These differences may be attributed to differences in maize genotypes and soil fertility. Additionally, the authors reported that the forage yield and the dry matter content were higher in the hybrids than in the native genotypes, which contrasts with our observations.
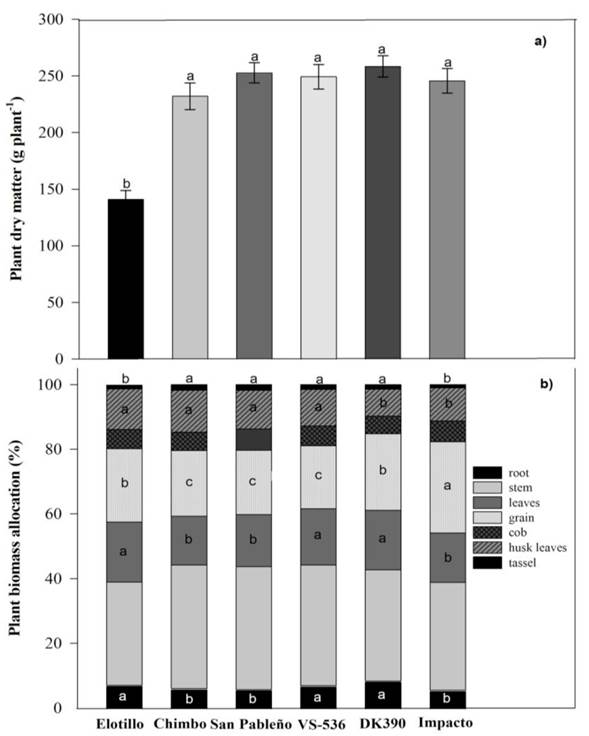
Figure 2 a) Plant dry matter (± ES) and b) biomass allocation by crop section in six tropical maize genotypes in Yucatan, Mexico. Distinct letters in different columns indicate significant differences (Scott-Knott, P ≤ 0.05); n = 24.
The most important component of DM was the stem with a mean of 35.5% of DM, followed by grain and leaves with 22.4% and 16.8%, respectively (Figure 2b). Differences between genotypes were found only in the plant components’ roots, leaves, grain, cob leaves, and tassel. Therefore, maize genotypes DK-390, VS-536, and Elotillo destined 7 to 8% and 18 to 19% of total DM to roots and leaves, respectively. These values represent 20% and 16% more than Chimbo, Impacto, and San Pableño, respectively. A more extensive root system gives better access to nutrients and water, greater resistance to adverse conditions, and increased structural stability. An expanded leaf area enhances both photosynthesis and biomass production, ultimately leading to a more resilient performance and increased productivity (Qi et al. 2012). The best cob leaves were observed in the landraces Chimbo, Elotillo, and San Pableño, and the genotype VS-536 destined 11 to 13% of DM to cob leaves. In the hybrids DK-390 and Impacto, the percentage of cob leaves was less with 8.3% and 10.2%, respectively. Grain percentage ranged from 19.5% in the genotype VS-536 to 28.2% in the hybrid Impacto. The hybrid DK-390 and the landrace Elotillo ranked second with 23.7% and 22.7% respectively, and San Pableño with 19.9%. Lastly, the tassel percentage which represents a mean of 1.4% of DM, ranged from 1.0 to 1.7%. The genotypes Chimbo, San Pableño, VS-536, and DK-390 were statistically superior. Elotillo and Impacto’s genotypes resulted in 1.0% and 1.1%, respectively (Figure 2b). Moreover, consistent with our results, Amador and Boschini (2000) illustrated that the stem exhibited the highest proportion of plant dry matter at 56.6%, with the cob and leaves following at 20.0% and 14.9%, respectively.
The higher grain yield was observed on both hybrids, Impacto and DK-390 with 3 800 and 3 400 kg ha-1, respectively (Figure 3a). The grain yields on the improved variety VS-536 and the landraces San Pableño and Chimbo were statistically similar, with a mean value of 2 700 kg ha-1. This result indicates the potential grain yield of some landraces concerning some commercial genotypes. In addition, in the study conducted by Dzib-Aguilar et al. (2011) in cambisols and luvisols in Yucatan, the GY of the VS-536 was 3 090 kg ha-1 higher than observed here. Therefore, in the Leptosol type where our study took place, the heterogeneous soil conditions, imposed limitations on the growth and potential yield of maize improved variety. In the landrace Elotillo, GY was lower with 1 800 kg ha-1 which may be attributed to the genotype's slow growth rate and vegetative traits. Nevertheless, farmers typically prioritize traits other than grain yield when selecting maize genotypes, such as culinary qualities, resistance to biotic and abiotic factors, and other relevant characteristics.
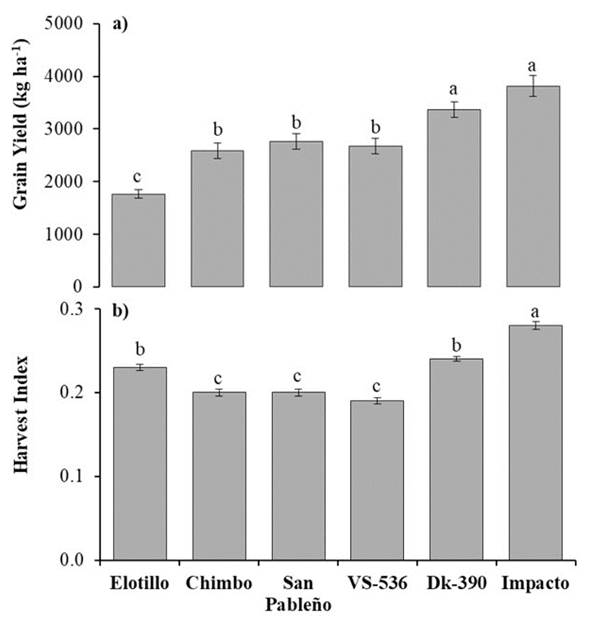
Figure 3 a) Grain yield and b) Harvest index of six tropical maize genotypes in Yucatan, Mexico. Means with a different letter are statistically different (Scott-Knott, P ≤ 0.05).
The higher HI was observed in the hybrid Impacto with a 0.28 index value, followed by the hybrid DK-390 and the landrace Elotillo with 0.24 and 0.23 index values, respectively (Figure 3b). In these genotypes, most of the dry matter accumulation was allocated for grain production. The landraces San Pableño and Chimbo had HI values of 0.20 and, the genotype VS-536 a 0.19 index value, respectively. So, maize landraces had lower HI values than the hybrids, that were developed with grain production objectives. These results are similar to what was reported by Rodríguez et al. (2016) with four tropical landraces in the region La Frailesca, Chiapas. The authors reported an HI of 0.17, 0.18, and 0.23 in the landraces Campeón, Maíz negro, and Macho, respectively.
Correlation between vegetative characteristics, dry matter, and grain yield
The results showed a positive and significant correlation between GY and DM (rxy = 0.65, p < 0.001), GY and HI (rxy = 0.64, p < 0.001), and GY and PH (rxy = 0.37, p < 0.05) (Table 2). Moreover, the variable DM showed a positive and significant correlation with DS (rxy = 0.51, p < 0.01), PH (rxy = 0.63, p < 0.001), and LA (rxy = 0.52, p < 0.01). Furthermore, LA also showed a positive significant relationship with PH and TNL. Finally, the variable DS had a positive and significant correlation with PH (rxy = 0.68, p < 0.001) and a negative significant correlation with HI (rxy =-0.33, p < 0.05).
Table 2 Correlation among vegetative traits of six tropical maize genotypes evaluated in Yucatan, Mexico.
| Trait | DS | PH | TNL | LA | DM | GY |
|---|---|---|---|---|---|---|
| PH | 0.68*** | |||||
| TNL | -0.05 | 0.23 | ||||
| LA | 0.21 | 0.34* | 0.33* | |||
| DM | 0.51** | 0.63*** | 0.22 | 0.52** | ||
| GY | 0.13 | 0.37* | 0.20 | 0.18 | 0.65*** | |
| HI | -0.33* | -0.14 | 0.01 | -0.29 | -0.16 | 0.64*** |
DS: days to silking; PH: plant height; TNL: total number of leaves; LA: leaf area; DM: plant dry matter; GY: grain yield; HI: Harvest index; * P < 0.05; ** P < 0.01; *** P < 0.001. n = 36.
Some general trends can be observed in Table 2. Plants with a longer growth cycle showed higher accumulated dry matter. The increased dry matter (DM) can be attributed to greater plant height (PH) and a larger leaf area (LA). Additionally, a greater number of leaves and a larger leaf area result in increased radiation interception, leading to higher plant dry matter (Díaz-López et al. 2013). Plants with higher dry matter content tend to have a higher grain yield, and the increased grain yield was explained by the higher harvest index (Table 2). Most studies reported positive correlations between dry matter and grain yield and the HI increases significantly with increasing grain yield (Liu et al. 2023).
In conclusion, all genotypes exhibited early maturity, which is a favorable trait under the agroecological conditions of Yucatan. Most genotypes exhibited superior growth rate and dry matter content compared to the landrace Elotillo. The highest grain yield was observed in both hybrids, DK-390 and Impacto. However, it is important to note that the profitability of these genotypes was not explored. Additionally, factors such as culinary qualities, resistance to biotic and abiotic stressors, and other relevant characteristics for farmers were not explored in this study.

