











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Los ecosistemas forestales tienen una función importante en el ciclo del carbono porque almacenan grandes cantidades de dicho elemento en la vegetación y en el suelo, al intercambiarlo con la atmósfera a través de la fotosíntesis y la respiración (Domínguez, 2011). Esas comunidades constituyen verdaderos sumideros de carbono para formar las estructuras de los organismos y que después se convierten, parcialmente, en materia orgánica (Pardos, 2010), por lo que la distribución de la biomasa en los árboles es crucial en la velocidad de acumulación de esta última y en el flujo de C (Chou y Gutiérrez, 2013).
El componente aéreo de los árboles es uno de los principales reservorios de biomasa; su producción y su presencia en las estructuras del árbol son dos características relevantes para su estimación en especies forestales (Pardos, 2010). La capacidad de los bosques para almacenar carbono en biomasa aérea varía por su composición florística, edad y densidad de población en cada estrato de la comunidad vegetal (Laumonier et al., 2010).
El manejo de los bosques bajo un enfoque de sustentabilidad ha conducido a la generación de ecuaciones alométricas que estiman la biomasa en diferentes secciones del árbol. Potencial que desde hace décadas se ha comprobado y utilizado para interpretar variaciones dimensionales y volumétricas en plantas (Niklas, 1994; Enquist y Niklas, 2002); y su aplicación ha sido útil en agricultura, en el estudio del funcionamiento de algunos ecosistemas y de los bosques (Soto et al., 2010; Chave et al., 2014).
Estas ecuaciones son útiles en la estimación del almacenamiento de carbono y en el cálculo parcial de la productividad primaria de muchos tipos de ecosistemas (Rodríguez et al., 2008; Návar, 2009; Acosta et al., 2011; Carrillo et al., 2014). La productividad primaria es un indicador del balance de carbono entre la vegetación y la atmósfera, así como una variable de interés en investigaciones de cambio climático global (Schlesinger, 1997; Clark et al., 2001).
Tema que se ha abordado en plantaciones de Hevea brasiliensis Muell. Arg (Monroy y Návar, 2004) y Bambusa oldhamii Munro (Castañeda et. al., 2005); en sistemas agroforestales (Callo et al., 2004; Roncal et al., 2008; Casanova et al., 2011; Soto et al., 2010); en selvas caducifolias (Rodríguez et al., 2008; Návar et al., 2013), subperennifolias (Aquino et al., 2015) y perennifolias (Montero et al., 2008; Douterlungne et al., 2013). Destaca el hecho de que aun cuando las selvas poseen una alta concentración de biomasa, todavía muchas especies no se han estudiado a cabalidad.
En el norte del estado de Oaxaca se localiza la tercera masa forestal más extensa del trópico húmedo mexicano (De Teresa et al., 2009), la cual ha sido sometida a fuertes procesos de disturbio que inciden en la dinámica del carbono, como la deforestación y el cambio de uso del suelo. Situación que hace necesario cuantificar o inventariar las reservas actuales de biomasa y carbono de estos ecosistemas. Zanthoxyllum kellermanii P. Wilson es una especie integrante de las selvas secundarias de La Chinantla, constituye un componente valioso en la sucesión ecológica y es apreciada por su madera (Manzano et al., 2010).
Por lo anterior, el objetivo del presente estudio fue estimar la biomasa aérea total, su distribución en el fuste, las ramas y el follaje en árboles de Zanthoxyllum kellermanii en una selva secundaria del norte de Oaxaca.
Materiales y Métodos
Área de estudio
El trabajo de campo se llevó a cabo en la localidad de San Martín Soyolapan, municipio Comaltepec, estado de Oaxaca, que se ubica fisiográficamente en la cuenca del río Papaloapan y en las estribaciones de la Sierra Norte de Oaxaca (17°41'53.5'' N y 96°16'57.3'' W) (Figura 1), a una altitud de 160 m.

Figura 1 Zona de estudio. Localización del estado de Oaxaca en los Estados Unidos Mexicanos (A), Región Sierra Norte (B), Santiago Comaltepec (C).
El clima se clasifica como Am(w)g, cálido húmedo con lluvias en verano, con una temperatura media anual de 24 °C y precipitación anual de hasta 3 590 mm (García, 1987). Geológicamente se observan afloramientos de rocas ígneas extrusivas y calizas con los siguientes grupos de suelos: lateríticos rojo-amarillentos, latosólicos amarillo-rojizos, litosuelos kársticos, migajones rojos y lateritas (Inegi, 2011). La vegetación predominante corresponde a selva alta perennifolia con abundancia de las siguientes especies arbóreas: Andira galeotiana Standley, Brosimum alicastrum Sw., Dialium guianense (Aub) Sandw., Terminalia amazonia (J. F. Gmel.) Exell y Vochysia guatemalensis Donn. Sm. (Miranda y Hernández, 1963).
Descripción de la especie
Zanthoxyllum kellermanii es un árbol que alcanza de 10 a 25 m de altura y hasta 80 cm de diámetro. Tiene corteza pardo-grisácea. Hojas pinnadas de 20 a 50 cm de largo, foliolos dispuestos en espiral de dimensiones 4 x 2 a 1.5 x 5.5 cm, de forma orbicular a elíptica. Especie dioica, las inflorescencias masculinas de hasta 30 cm de largo y femeninas de hasta 10 cm, ambas en panículas subterminales y axilares. Frutos capsulares, triangulados, moreno negruzcos, de 5 a 7 mm. Se desarrolla en los bosques húmedos, a una altitud de 50 a 700 m, desde el sur de México a Perú. Forma parte del estrato medio o superior de selvas altas perennifolias, especialmente de Terminalia amazonia, en suelos laterítico profundos derivados de arrastres aluviales antiguos. Su madera tiene un bello veteado, por lo que se le utiliza para el aserrío y en la fabricación de tablas para construcciones rurales (Pennington y Sarukhán, 2005).
Construcción de las ecuaciones alométricas
Se seleccionaron dos árboles por cada una de las siguientes categorías diamétricas: 10, 15, 20 y 25 cm, lo que responde a que este tamaño de muestra ha sido aplicado en otros estudios del trópico con resultados aceptables (Brown et al., 1995). Antes de que los ejemplares fueran derribados se les midió el diámetro normal (DN) con una cinta diamétrica Ben Meadows de 5 m de longitud. Una vez derribados, cada árbol fue seccionado en tres partes: fuste, ramas y follaje.
Los fustes al igual que las ramas se cortaron en tramos de dos metros y el follaje se separó de las ramas y se empaquetó en porciones de 0.5 kg en bolsas de plástico (Domínguez, 2011). Cada componente se colocó de manera independiente en una báscula Torrey de 500 kg para obtener su peso fresco in situ.
Se obtuvieron cinco muestras por sección de cada individuo de las categorías diamétricas de 10 y 15 cm, y 10 muestras por sección de cada árbol de las categorías de 20 y 25 cm. Las correspondientes a las ramas y fustes fueron secciones trasversales (rodajas) de 3 y 5 cm de grosor, respectivamente. El peso fresco se registró in situ para todas las partes con una báscula de balanzón Oken modelo C-A de 5 kg de capacidad y una aproximación de 5 g. La totalidad de las muestras se depositó en bolsas de plástico para su traslado a los Laboratorios Generales del Colegio de Postgraduados Campus Montecillo, previo registro de sus datos de control (Figueroa et al., 2005; Domínguez, 2011).
En el laboratorio, las muestras de ramas y follaje se guardaron en bolsas de papel y así se colocaron en estufas de secado marca Felisa. La temperatura para deshidratar las de ramas y de hojas fue de 75 °C y la, de fustes a 80 °C, mismas que se mantuvieron hasta alcanzar un peso constante. Con estos datos se estableció la relación entre peso fresco y peso seco para estimar la proporción de biomasa por componentes, así como de la parte aérea total, mediante la ecuación 1 (Domínguez, 2011):
 1
1
Donde:
PS = Peso seco
PF = Peso fresco
PScom = Peso seco del componente arbóreo
PFcom = Peso fresco del componente arbóreo
Los valores de biomasa por componente de cada ejemplar muestreado a partir de los datos de peso fresco y peso seco se ajustaron con ecuaciones alométricas, con técnicas de regresión no lineal. Fue ajustado un modelo potencial mediante el procedimiento NLIN del paquete estadístico SAS versión 8.0 (SAS, 1999). Este modelo (ecuación 2) ha sido ampliamente utilizado en diferentes estudios de determinación de biomasa aérea en árboles (Acosta et al., 2002; Figueroa et al., 2005; Carrillo et al., 2014):
 2
2
Donde:
Y = Peso estimado del componente arbóreo en kg
b0 y b1 = Coeficientes de la regresión
X = Diámetro normal en cm
Con las ecuaciones alométricas se elaboraron gráficas en la hoja de cálculo Excel (Microsoft, 2003) para la biomasa aérea total y de los componentes del árbol.
La bondad de ajuste del modelo se evaluó a través de los valores estadísticos del análisis de varianza: cuadrado medio del error (CME), coeficiente de determinación (R2) y nivel de significancia (p < 0.05).
Resultados y Discusión
Ecuaciones alométricas
En el Cuadro 1 se presentan las ecuaciones alométricas generadas para estimar la biomasa aérea total y de componentes, así como sus estimadores de variación y confiabilidad estadística. Todas las ecuaciones mostraron un buen nivel de ajuste (p < 0.0001), y tanto los valores de R2 como los cuadrados medios del error (CME) indican que el modelo es confiable para estimar la biomasa aérea a partir del diámetro normal.
Cuadro 1 Ecuaciones alométricas para la estimar biomasa aérea total y de los componentes de árboles de Zanthoxylum kellermanii P. Wilson.

N = Tamaño de muestra; R2 = Coeficiente de determinación; CME = Cuadrado medio del error; Fc = F calculada, Y = Biomasa (kg), X = Diámetro normal (cm).
Las curvas de regresión ajustadas para estimar la biomasa aérea se ilustran en la Figura 2, para los componentes del árbol y para el individuo completo.

Distribución de biomasa aérea
La biomasa aérea en cada uno de los componentes de Zanthoxylum kellermanii se dispuso de la siguiente manera: en el fuste el intervalo tuvo una amplitud de valores entre 77.50 % a 87.52 % en las categorías diamétricas 10 y 25 cm; en las ramas fue de 1.00 % a 17.52 %, en los mismos diámetros, y la variación del follaje fue de 1.47 % a 4.98 % en los de 25 y 10 cm. La proporción promedio de biomasa por sección fue de 83.21 % en fuste, de 13.57 % en ramas y de 3.22 % en follaje, lo que incluye a todas las categorías diamétricas (Cuadro 2 y Figura 3).
Cuadro 2 Peso seco (kg) y distribución de biomasa aérea (%) en los componentes arbóreos para las diferentes categorías diamétricas de Zanthoxylum kellermanii P. Wilson.

CD = Categoría diamétrica.
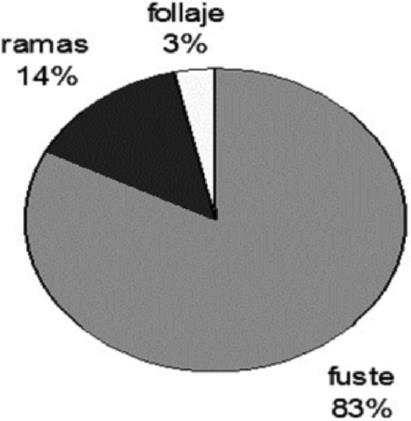
Figura 3 Distribución proporcional de la biomasa aérea en cada uno de los componentes arbóreos de Zanthoxylum kellermanii P. Wilson.
El peso seco en las diferentes secciones de los árboles se distribuyó en los intervalos: follaje, de 0.494 kg a 2.136 kg; ramas de 1.738 kg a 8.055 kg y fuste 7.690 a 206.725 kg, para las categorías diamétricas de 10 y 25 cm en todos los casos; la mayor proporción de la biomasa aérea se concentró en el fuste y la menor en el follaje. Los valores de biomasa total del árbol van de 9.923 a 236.727 kg en categorías diamétricas de 10 y 25 cm (Cuadro 2).
Los resultados del presente estudio revelan un comportamiento semejante de la distribución de biomasa en relación con otras investigaciones en el trópico húmedo mexicano, ya que concentra gran parte en el fuste (83.21 %). Al respecto, Monroy y Návar (2004) calcularon 73.88 % en el fuste total, y 27.09 % en ramas para individuos de una plantación de Hevea brasiliensis en Uxpanapa, Veracruz; y Castañeda et al. (2005) registran valores de 83.7 % en tallos, 12.3 % en follaje y 4 % en ramas de Bambusa oldhamii Munro en Huatusco, Veracruz.
Otros estudios de biomasa en especies de selvas altas en Tabasco (Montero et al., 2008) y Costa Rica (Fonseca et al., 2009) confirman que una mayor proporción de biomasa en el fuste se incrementa con la edad y el tamaño del árbol, por lo que algunas investigaciones únicamente basan su metodología en el volumen del tronco (Návar et al., 2013).
La biomasa de ramas registró un porcentaje intermedio (13.57 %) en contraste con la biomasa de fuste y de las hojas. Goodman et al. (2014) señalan que la primera está determinada por las dimensiones y la arquitectura de la copa, así como la calidad de sitio y características del rodal, entre las que destaca el espacio de crecimiento (Foli et al., 2003).
La biomasa del follaje tuvo el valor más bajo en la distribución de los componentes arbóreos (3.22 %); al igual que en las ramas, esta biomasa fue proporcionalmente superior en los árboles con diámetros pequeños y disminuyó, progresivamente, con el aumento del mismo. Los individuos en esta condición son dominados y disponen de poca luz (López et al., 2006), lo que sugiere una condición limitante a modo de la Ley del Mínimo en la que las cantidades de recurso disponible están muy cercanas a lo indispensable (Odum, 1972); y en este caso particular, se relaciona con el supuesto de deficiencia de crecimiento, en el que se manifiesta que la producción de madera del tallo tiene menor prioridad, comparada con la producción de follaje y raíces.
Enquist et al. (1999) demostraron que la tasa de crecimiento en árboles tropicales tiene una relación potencial con la biomasa. Para el presente estudio, la especie presenta incrementos de diámetro superiores a 1.5 cm año-1 favorecidos por copas con buena calidad (simétrica, vigorosa) y expuestas, tanto vertical como lateralmente a la luz solar (Manzano et al., 2010). Los mismos autores señalan que siendo una especie de sucesión temprana, puede ser utilizada en programas de recuperación y enriquecimiento de áreas con disturbio en la zona de estudio.
Conclusiones
Las ecuaciones alométricas generadas para Zanthoxylum kellermanii mostraron un buen nivel de ajuste, determinado por sus valores estadísticos, por lo que el modelo es confiable para realizar una predicción atinada de la biomasa aérea a través del diámetro normal.
El patrón de distribución de la biomasa, respecto al peso seco total, indica que la especie tiene su mayor potencial para acumular biomasa en el fuste.
Conflicto de intereses
Los autores declaran no tener conflicto de intereses.
Contribución por autor
Filemón Manzano Méndez: trabajo de campo y de gabinete y elaboración del manuscrito; Juan Ignacio Valdez Hernández: trabajo de gabinete y elaboración del manuscrito; Miguel Ángel López López: trabajo de campo y de gabinete y elaboración del manuscrito y Elizandro Pineda Herrera: trabajo de gabinete, preparación y revisión del manuscrito.

