











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkHighlights:
AGB production and ANPP increased across the Chronosequence.
The 30-years old stand produced 79 % of the AGB observed in the NMF.
In both, the 30-years old stand and the NMF, a high AGB corresponded with high ANPP, showing 87 % similarity.
Stems are the structural component with the highest AGB allocation.
Introduction
Forest biomass accumulation leads to timber production, the main ecosystem provisioning service, supporting the existence of other ecosystem services (support, regulating and cultural services) (Brockerhoff et al., 2017). The main objective of forest management is to increase its aboveground biomass (AGB), particularly towards tree stems. However, sustainable forest management aims not only to guarantee timber production but also to maximize the other ecosystem services, which allow self-regulation and long-term permanence of forests (Duncker et al., 2012). Currently, one of the most important research challenges is to know how to manage forests for timber production while enhancing the other ecosystem services (habitat and biodiversity conservation, natural hazard protection, recreation, water cycle regulation and carbon sequestration) (Häyhä, Franzese, Paletto, & Fath, 2015). Studies are still needed to determine whether a managed forest could provide ecosystem services similar to those of a natural area and at what point after harvesting occurs.
Sustainable forest management stimulates soil cover through silvicultural treatments such as release cutting, weeding, thinning, pruning; until the forest reaches the cutting cycle (Vignote, Martínez, & Villasante, 2011). Meanwhile, nutrient and water supplementation continues because of the maintenance of biogeochemical and water cycles, without external inputs. In addition, intermediate silvicultural practices, applied to the stand favors the accumulation of new AGB (Porter-Bolland et al., 2012; Vignote et al., 2011). Silvicultural practices are focused on directing biomass to the tree stems, and next to the other structural tree’s components (branches and foliage). However, this also depends on the stand density and age, species composition and site quality (Schulze, Körner, Law, Haberl, & Luyssaert, 2000).
Biomass allocation to the stem is essential in timber production, while tree leaves, and branches are more important to water and nutrients dynamics. Leaves and branches actively participate in ecosystem processes such as photosynthesis, C sequestration, litter decomposition, water and nutrients uptake, and evapotranspiration. Therefore, accumulated biomass result in enhanced C capture (Penman et al., 2003), thus the rate of nutrients and organic matter incorporation to the soil through litter decomposition is more efficient and economical. However, decomposition rates depend on the structural tree components, presenting greater lability (easy decomposition) the green foliage that remains on the ground after harvesting, followed by more lignified branches (Pérez-Suárez, Arredondo-Moreno, & Huber-Sannwald, 2012). In managed forests, nutrient cycling and C sequestration remain active because of the periodic inputs of cut residues to the forest floor (Mayer et al., 2020), hence contributing to the maintenance of soil fertility, and therefore the stand productivity. In this way, knowing and predicting the accumulation of tree biomass is one of the main ways to determine the forest management effect on C sequestration and its role in climate change mitigation (Montero, Ruiz-Peinado, & Muñoz, 2005). Deciding which silvicultural alternative enhances biomass accumulation to a particular structural tree component, based on growth models, is important to forest productivity (Barrero, Alvárez, Nepveu, García, & Guera, 2011).
Pinus patula Schiede ex Schltdl. & Cham. is endemic to Mexico and an important timber species in temperate forests. This species forms pure even-aged stands, as well as in mixed forest, where it is the dominant species (Ángeles-Pérez, 2009). Pinus patula is a fast-growing species found in highly productive sites where it can reach its maximum productive potential (Velázquez, Ángeles, Llanderal, Román, & Reyes, 2004). This species is used in forest recovery and conservation, with the aim of increasing C sequestration and storage in biomass through forest management programs (Secretaría de Medio Ambiente y Recursos Naturales [SEMARNAT], 2014). Quantifying AGB in P. patula managed stands and compared to natural stands under development allows observing the effect of forest management. This could enhance the benefits obtained from only timber production, by reversing environmental degradation, conserving natural resources, promoting participatory management, governance of natural resources, and promoting human development through sustainable development policies and programs. Based on above, the objective of the present study was to evaluate the AGB along a temperate forest chronosequence and to examine the role of forest management for timber production on patterns of AGB fixation, increment and allocation of stands dominated by P. patula in central Mexico.
Materials and methods
Study area
The study was conducted in a temperate forest dominated by P. patula in the ejido La Mojonera, in the Zacualtipán region of Hidalgo (Figure 1) in central part of Mexico (Sierra Alta Hidalguense) along the Trans-Mexican Volcanic Belt. The site belongs to the Network Intensive Carbon Monitoring Sites (MEX-SMIC network) (Ángeles-Pérez et al., 2012) that is part of the MexFlux network (Vargas et al., 2013). The topography of the site is characterized by a moderate slope with a mean altitude of 2 100 m. The dominant climate is humid temperate (C(fm)) with mean annual temperature of 13.5 °C and mean annual precipitation of 2 050 mm (García, 2004). The typical soil is phaeozem haplic, with high organic matter and nutrient content. The main vegetation cover types are oak forest, pine-oak, and mountain cloud forests (Ángeles-Pérez, 2009; Ponce-Vargas, Luna-Vega, Alcántara-Ayala, & Ruiz-Jiménez, 2006).
Figure 1 Stands area in the ejido La Mojonera, forest region of Zacualtipán, Hidalgo, central Mexico.
In this study a natural forest area (NMF: non managed) of mixed pine-oak forest was selected and it was used as reference to compare its productive growth to that of managed stands. The NMF is a more species diverse forest than the stands regenerated after the harvesting process, since in the latter, species of higher economic value, such as P. patula are favored. In the tree and shrub strata were identified approximately 15 species. The dominant species were Quercus laurina Bonpl., P. patula, Cornus disciflora Moc. & Sess. ex D.C., Vaccinium leucanthum Schltdl., Ternstroemia sylvatica Schltdl. & Cham., and Clethra mexicana D. C. In less proportion species such as Rhamnus capraeifolia Schltdl., Viburnum tiliaefolium Oerst., Q. excelsa Liebm., Crataegus pubescens (Kunth) Steud, Ilex discolor Hemsl., Prunus serotina Ehrh., Arbutus xalapensis Kunth., and Cleyera theaoides Sw, were also recorded in smaller proportions. In the NMF we recorded 896 trees∙ha-1 (71 % of total density) with diameter at breast height (dbh) range of 5 to 20 cm, while 317 trees∙ha-1 had a dbh > 20 cm. The maximum DBH size observed was 61 cm. The average basal area in the NMF was 36 m2∙ha-1, of which 63 % (22 m2∙ha-1) corresponded to P. patula.
Forest management history in Zacualtipán region
Forest management in the study area dates back to 1980 when landowners implemented programs for timber production and habitat conservation (Musálem, 2006). To generate even-aged stands of P. patula, the seed-tree method has been used based on natural regeneration (Castelán-Lorenzo & Arteaga-Martínez, 2009; Velázquez et al., 2004). When natural regeneration is insufficient, it is also combined with enrichment planting. All stands have the same forest management history because they receive the same silvicultural treatments such as site preparation, release cutting, weeding, thinning, pruning, improvement cuttings, and salvage cuttings. However, thinning is the most common silvicultural practice applied to produce commercial timber; moreover, it reduces competition among remaining trees, and helps to control tree species composition. These forests are part of a network of Measurement, Reporting and Verification (MRV) sites to evaluate the growth and mortality of stands under forest management supervised by the regional communities. The ejido La Mojonera have been recognized for its collective work in forest care, receiving the National Forestry Merit Award. Therefore, forest management and conservation projects contribute to the conservation and recovery of the country's vegetation cover, being an example of success in community forest management indicators.
Field sampling
A seven-stand chronosequence corresponding to the harvesting years 1998, 1995, 1990, 1988, 1986, 1984, and 1982 were selected for this study (Figure 1). Each year corresponds to the application of the seed-tree regeneration method. The chronosequence was compared with the NMF of approximately 80-years old. Three permanent sampling units (PSUs) of 0.04 ha (20 × 20 m) were randomly established in stands and the NMF. All live trees with a DBH ≥ 5 cm, at 1.30 m above the ground level were tagged and the total height of all tree individuals was recorded. In the analyses, chronosequence measurements were compared in three subsequent years (2005, 2008 and 2012), while NMF measurements were only recorded in 2005 and 2012.
Aboveground biomass, its increment and allocation
AGB was estimated with local allometric equations previously parameterized for
the dominant tree species (conifer and broadleaved species) in the study site
(Soriano-Luna, Ángeles-Pérez,
Martínez-Trinidad, Plascencia-Escalante, & Razo-Zárate, 2015).
First, AGB was calculated for individual tree and each tree structural component
(stems, bark, branches, and foliage). The biomass from each tree was summed-up
to estimate the biomass in the corresponding stand. The aboveground biomass
increments (AGBI) were calculated as the difference between the AGB of trees in
each stand of the chronosequence compared to measurements from later years (2008
or 2012), with the equation:
Modeling aboveground biomass
The stands AGB production was analyzed with the parameterization of the following growth models: Chapman-Richards (Zhao-gang & Feng-ri, 2003), Holling type III, and Schumacher (Peng, Zhang, & Liu, 2001). AGB was estimated from the data collected in the stands of a chronosequence aged 2 to 30 years old, including all the tree species. The best model was selected according to the goodness of fit parameters, i.e., adjusted coefficient of determination (R2); mean square error (MSE); and sum of squares errors (SSE). Also, the current annual increment (CAI) and mean annual increment (MAI) were calculated in AGB with the model with the best goodness-of-fit parameters.
Aboveground net primary productivity
The aboveground net primary productivity (ANPP) of each stand was estimated with the equation ANPP = ∆B + M; where ∆B is the aboveground annual biomass increment and, M is the annual litterfall production (Clark et al., 2001). Herbivory was not considered in this calculation.
The percentage ANPP of each stand was calculated and compared to that of the NMF. To estimate annual litterfall production, four circular litter traps were installed in each PSUs (Rocha-Loredo & Ramírez-Marcial, 2009). The traps were equipped with a highly flexible nylon sheet (medium mesh size) with a surface area of 0.5 m2. The litterfall was collected each month from October 2005 to September 2008. On each sampling date, the litter was separated as follows: leaves, twigs, and other material, including bark and reproductive structures. Leaves were sorted according to tree species. All vegetative components were oven-dried at 65 °C for 72 h and then weighed.
Statistical analysis
AGB data were subjected to normality tests such as Shapiro-Wilk and Kolmogorov-Smirnov before further analysis. Then, one-way analysis of variance (ANOVA) was used to detect the effects of stand age on the evaluated variables: AGB production and its allocation to structural tree components, AGBI, ANPP, stand density, and tree mortality. Subsequently, a Tukey HSD test (α ≤ 0.05) was applied for means comparison. All statistical analyses were performed in the SAS/ETS® SAS Inc. software (SAS Institute Inc., 2009).
Results
AGB production, increment and allocation by stand age
AGB production showed statistically significant differences (P ≤ 0.05) among stands (Table 1), in all measurements. In 2005, the 1998 stand (7-years old) reached 32.27 Mg∙ha⁻1 of AGB, representing 16.6 % of the AGB recorded in the NMF (Table 1). In 2012, this same stand (now 14-years old) increased its AGB up to 71.10 Mg∙ha-1, only three times lower than in the NMF (Table 1). In comparison the 1982 stand (30-years old) recorded in 2012 more than 78 % of the AGB reported in the NMF. Some stands showed a reduction in AGB as a result of silvicultural practices, specifically in the 1998 stand (Table 1). By 2012, in that stand, tree density decreased drastically (56 % in a seven-year period) as a consequence of a 50 % thinning intensity during the previous year (2011) (Table 2).
Table 1 Aboveground biomass (AGB) and aboveground biomass increment (AGBI) of even-aged stands dominated by Pinus patula in a seven-year period (2005-2012) in a temperate forest Chronosequence in Zacualtipán, Hidalgo, Mexico.
| Stand (harvesting year) | Stand age (years) | AGB (Mg∙ha-1) | AGBI (Mg∙ha-1∙año-1) | ||||
|---|---|---|---|---|---|---|---|
| 2005 | 2008 | 2012 | 2005-2008 | 2008-2012 | 2005-2012 | ||
| 1998 | 7 | 32.27 ± 0.84 d | 80.51 ± 1.25 cd | 71.10 ± 4.27 d | 16.12 ± 0.13 a | 6.05 ± 0.94 a | 7.68 ± 0.61 a |
| 1995 | 10 | 30.31 ± 2.81 d | 69.67 ± 3.00 d | 99.95 ± 1.11 d | 13.12 ± 0.19 ab | 8.25 ± 0.39 a | 10.17 ± 0.21 a |
| 1990 | 15 | 69.38 ± 6.89 cd | 91.57 ± 13.29 bcd | 111.12 ± 19.51 cd | 7.42 ± 2.04 c | 5.76 ± 1.44 a | 6.31 ± 1.73 a |
| 1988 | 17 | 80.81 ± 7.73 bcd | 110.71 ± 8.69 bcd | 130.36 ± 6.52 bcd | 10.10 ± 0.67 bc | 8.73 ± 0.84 a | 8.76 ± 0.68 a |
| 1986 | 19 | 92.90 ± 7.81 bcd | 129.41 ± 10.84 abc | 125.92 ± 5.84 bcd | 12.02 ± 1.01 abc | 6.05 ± 0.88 a | 7.49 ± 0.61 a |
| 1984 | 21 | 129.13 ± 17.36 abc | 141.13 ± 21.04 ab | 166.74 ± 23.44 abc | 10.97 ± 1.37 abc | 6.34 ± 0.52 a | 8.08 ± 0.85 a |
| 1982 | 23 | 136.87 ± 2.81 ab | 161.67 ± 0.92 a | 173.96 ± 4.01 ab | 9.42 ± 0.95 bc | 5.64 ± 0.36 a | 7.09 ± 0.32 a |
| NMF | 80 | 194.36 ± 33.56 a | 223.44 ± 10.88 a | 10.53 ± 2.91 a | |||
(Standard error of the mean. Different letters indicate statistically significant differences between stands (Tukey, P ≤ 0.05 significance level). Empty cells indicate that AGB was not measured during that year. Stand age corresponds to the age at the 2005 measurement. NMF: non-managed forest.
Table 2 Density and mortality of even-aged stands dominated by Pinus patula in a seven-year period (2005-2012) in a temperate forest chronosequence in Zacualtipán, Hidalgo, Mexico.
| Stand (harvesting year) | Stand age (years) | Density (trees∙ha-1) | Mortality (trees∙ha-1) | ||||
|---|---|---|---|---|---|---|---|
| 2005 | 2008 | 2012 | 2005-2008 | 2008-2012 | 2005-2012 | ||
| 1998 | 7 | 3 917 ± 180.47 a | 3 950 ± 212.62 a | 1 708 ± 22.04 ab | -33 (0 %) | 2242 (57 %) | 2 209 (56 %) |
| 1995 | 10 | 2 158 ± 209.33 b | 2 167 ± 173.40 b | 1 917 ± 152.97 ab | -9 (0 %) | 250 (11 %) | 241 (11 %) |
| 1990 | 15 | 1 375 ± 156.12 c | 1 400 ± 125.83 bc | 1 292 ± 136.42 abc | -25 (0 %) | 108 (8 %) | 83 (6 %) |
| 1988 | 17 | 1 117 ± 181.62 c | 1 100 ± 175.00 c | 867 ± 110.23 c | 17 (2 %) | 233 (21 %) | 250 (22 %) |
| 1986 | 19 | 1 000 ± 94.64 c | 1 058 ± 283.70 c | 775 ± 90.13 c | -58 (0 %) | 283 (27 %) | 225 (22 %) |
| 1984 | 21 | 958 ± 174.60 c | 1 058 ± 283.70 c | 1 208 ± 284.80 bc | -100 (0 %) | -150 (0 %) | -250 (0 %) |
| 1982 | 23 | 1 600 ± 150.00 bc | 1 792 ± 134.11 bc | 1 942 ± 79.49 a | -192 (0 %) | -180 (0 %) | -342 (0 %) |
| NMF | 80 | 1 233 ± 22.04 c | 1 300 ± 75.00 abc | -67 (0 %) | |||
( Standard error of the mean. Different letters indicate statistically significant differences between stands (Tukey, P ≤ 0.05). For mortality, negative numbers indicate that no dead trees were reported during the measurement period. The percentage of dead trees are shown in parentheses. Empty cells indicate that density was not measured during that year. Stand age corresponds to the stand age at the 2005 measurement. NMF: non-managed forest.
The AGB increment (AGBI), which represents the magnitude of AGB variation with respect to time, decreased as stand age increased, with statistically significant differences (P ≤ 0.05) among stands (Table 1), specifically in the measurement of 2005 to 2008. Thus, in a three-year period (2005-2008) AGBI were 2.1 times higher than those recorded in an overlapping four or seven-year period (2008-2012 and 2005-2012, respectively), in which not statistically significant differences (P > 0.05) between stands were detected.
AGB of all structural components increased significantly with stand age (Figure 2). As expected, the highest proportion of AGB was allocated to stems, the main reservoir of trees in all stands. In particular, trees in the 1998 stand (7-years old) allocated 59.9 Mg∙ha⁻1 of AGB to stems and bark, three times more than the observed allocation to stems in the NMF. In contrast, AGB allocation to branches decreased as stand age increased. Thus, 18 % AGB was allocated to branches for the 1998 stand and by 14 % for the 1982 stand. AGB allocated to foliage corresponded to 4 to 9 % of the total and was higher in younger stands (7 to 17-years old, 1998 and 1995 stands, respectively).
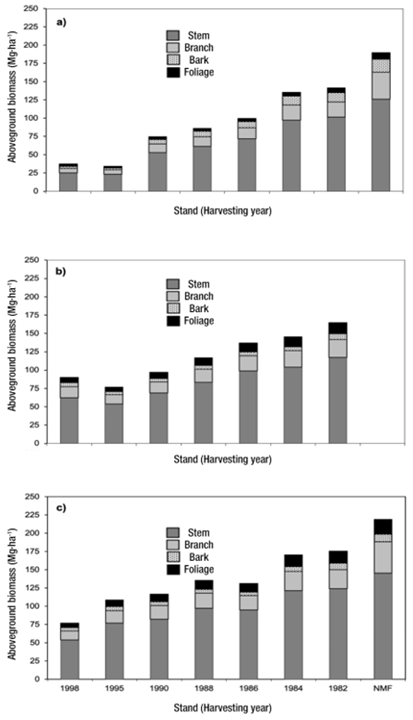
Figure 2 Allocation of aboveground biomass (AGB) to tree structural components in even-aged stands dominated by Pinus patula in a seven-year period: 2005 (a), 2008 (b), and 2012 (c) for a temperate forest chronosequence in Zacualtipán, Hidalgo, Mexico. In 2008 (b) the AGB was not measured in non-managed forest (NMF) stand.
Aboveground biomass growth model
The Chapman-Richards model showed the best goodness-of-fit parameters for predicting AGB stand growth (Figure 3), where all parameters were significantly different from zero (P < 0.0001) at the 95 % confidence level (Table 3). Specifically, this model had the highest R2 value (0.76), the lowest MSE (834.9) and SSE (85 990.9). According to the model, the maximum CAI occurred at 15-years, and the maximum MAI at 5-years of stand age. CAI and MAI reached the same value of 5.9 Mg∙ha⁻1∙year⁻1 after 12-years of stand regeneration (Figure 4).
Table 3 Goodness-of-fit statistics and mathematical model parameters for estimating aboveground biomass (AGB) production according to stand age (t) in a temperate forest chronosequence in Zacualtipán, Hidalgo, Mexico.
| Model | Equation | Estimate parameters | SSE | MSE | R2 | ||
|---|---|---|---|---|---|---|---|
|
|
|
|
|||||
| Chapman-Richards |
|
244.42 | 0.04 | 1.29 | 85 990.90 | 834.90 | 0.76 |
|
|
(26.80) | (0.01) | (0.18) | ||||
| Holling type III |
|
213.12 | 17.53 | 93 506.50 | 899.10 | 0.74 | |
|
|
(15.52) | (1.59) | |||||
| Schumacher |
|
251.37 | 14.74 | 97 800.70 | 940.40 | 0.72 | |
|
|
(17.73) | (1.36) | |||||
Standard error shown in brackets. SSE = sum of squares errors; MSE = mean square errors.

Figure 3 Relationship between observe and predicted aboveground biomass (AGB) production accumulation according to the stand age in a temperate forest chronosequence in Zacualtipán, Hidalgo, Mexico.

Aboveground net primary productivity
As observed for AGB, ANPP increased as stand age increased (Table 4). ANPP in the oldest stand after harvesting (30-years old) was 23 % lower than that observed in the NMF. The highest ANPP was recorded in the 1982 stand (30-years old) and was similar to the 1995 stand (17-years old). The same pattern was recorded in the 1998 and 1986 stands, which had an ANPP of 14.7 and 14.3 Mg∙ha⁻1∙year⁻1, respectively. Even though the 1998 stand had the highest tree mortality (Table 2) as a result of thinning, ANPP was not affected. Overall, the ANPP of the managed stands was only 27 % lower than that observed in the NMF.
Table 4 Aboveground net primary productivity (ANPP) in even-aged stands dominated by Pinus patula in a temperate forest chronosequence in Zacualtipán, Hidalgo, Mexico.
| Stand | Age in 2012 (years) | ANPP (Mg∙ha-1∙year-1) | ANPP(%) |
|---|---|---|---|
| 1998 | 14 | 14.7 | 77 |
| 1995 | 17 | 16.1 | 84 |
| 1990 | 22 | 13.9 | 73 |
| 1986 | 26 | 14.3 | 75 |
| 1982 | 30 | 16.5 | 87 |
| Non-managed forest | 87 | 19.1 | 100 |
Discussion
Production, increment and AGB allocation by stand age
Sustainable forest management aims to promote timber production while maintaining other ecological services. In this research we evaluated the role of forest management on increasing AGB production along a P. patula chronosequence. We found that a managed forest reached the maximum AGB accumulation in a shorter time than a natural forest. The AGB allocation and AGBI allowed estimating time in which a stand, as a result of forest management, may function similar to a natural unmanaged forest.
The results on AGB were similar to findings in other studies carried out in the same region (Chávez-Aguilar et al., 2016; Figueroa-Navarro, Ángeles-Pérez, Velázquez-Martínez, & de los Santos-Posadas, 2010; Soriano-Luna et al., 2018). Even with studies in other sites and species, for example, Peichl and Arain (2006) studied Pinus strobus L. stands of different ages (2, 15, 30, and 65-years old) and reported significant increments in AGB as stand age increased. In the same way, Jagodziński, Dyderski, Gęsikiewicz, and Horodecki (2019) reported that total AGB in 3- to 117-year-old Pinus sylvestris L. even-aged stands increased with increasing stand age. On the other hand, Mendoza-Ponce and Galicia (2010) found AGB production of 74.0 and 127.0 Mg∙ha-1 in 28- and 30-year-old Pinus stands, respectively, in a temperate forest landscape in central Mexico; these amounts are lower than those recorded for even-aged stands in the present study (Table 1). However, these differences in AGB storage capacity and production depend on stand characteristics (species composition, stage of development, land use, and stand density) and forest regeneration. The AGB results in the present study are higher (Table 1) compared to those reported in other studies with even-aged stands, which could be a good indicator that the forest management scheme applied in the study site contributes significantly to the increase in AGB.
The most applied silvicultural practice to promote tree growth in the study site is thinning, which serves to decrease tree competition and promote the importance of the forest harvest by increasing the volume, size and commercial quality of the timber. Thinning changes tree growth and increases AGB at both the individual tree and stand levels. Several studies have indicated that thinning is able to increase AGB in stands (Porter-Bolland et al., 2012; Rodríguez-Ortíz et al., 2011; Wang, Chang, Lee, Yang, & Lin, 2005) and have demonstrated the impact of such a practice on stand structure, especially regarding the C cycle, soil biodiversity, and improved ecological dynamics (Marchi, Paletto, Cantiani, Bianchetto, & De Meo, 2018).
The AGB production also depends on the allocation to the structural components of the tree, such as stems, branches, and leaves. In the present study, the highest AGB production was allocated to stems, which represented 70 % of total mean tree biomass, favored by the thinning practices. Rodríguez-Ortíz et al. (2011) reported in P. patula forest that AGB allocation is directed toward stems regardless of thinning intensity. The implementation of thinning has also been recommended as one of the main tools to promote AGB allocation into stems, since the removal of a fraction of the population in the stands changes the AGB allocation. Moreover, a higher AGB allocation to stems in a shorter period than that observed in the NMF, generates stand stability and high-quality timber production (Marchi et al., 2018), in this way, the economic profitability of forests with forest management increases at the same time that some provision ecosystem services (timber, through forest biomass accumulation) are recover. In this study, AGB allocation to trees structural components varied with stand age and was similar to a natural forest. Therefore, AGB allocation may decline as managed stands aged because net primary productivity (NPP) and biomass accumulation rates decrease as respiration rates increase (Ryan, Binkley, Fownes, Giardina, & Senock, 2004). Forest management is based on the general productivity theory that predicts that forest NNP steadily increases to a maximum as forests age and then gradually declines (Chávez-Aguilar et al., 2016; Rodríguez-Ortíz et al., 2011).
In this research, ANPP as result of the harvesting process did not show a behavior according to stand age (unlike AGB), because trees in early age use their resources to increase dimensions and their efficiency in nutrient assimilation, which is higher compared to older trees. Therefore, this study showed that the 1995 stand (the youngest stand) had an ANPP only 2.5 % lower than that observed in the oldest stand (1982), which had an ANPP only 12 % lower than the NMF (19.1 Mg∙ha⁻1∙year⁻1) having the highest ANPP (Table 4). Older stands did not undergo thinning during the study period, although similar thinning schedules are prescribed for all stands during the rotation period. Some studies have shown that thinning initially reduces stand NPP while increasing the NPP of residual trees (Law, Riitters, & Ohmann, 1992). Hu, Lehtonen, Minunno, and Mäkelä (2020) estimated the AGB allocation in P. sylvestris and Picea abies (L.) H. Karst forests of various ages and found that AGB allocation in all components increased with age in both forest species; furthermore, they observed that AGB allocation stabilized after trees reached the age of maturity (67 to 100 years). In this research, AGB and ANPP were higher in older stands than in younger stands, confirming that age influences the distribution not only of AGB, but also of ANPP.
Aboveground biomass growth model
Growth models applied in managed forest are a useful tool to anticipate (modeling) stand development and predict yields, which is essential for decision making in the application of timely silvicultural practices that direct forest health and growth (Schöngart, 2008).
The Chapman-Richards model predicted that a 30-years old stand reaches 85 % of the AGB observed in the NMF, reflecting high timber production in a younger stand compared to a natural forest, where natural succession occurs, but with the same quality and quantity of timber produced. Several studies have described the growth patterns in managed forests; however, most were parameterized in terms of diameter growth, basal area, height, and volume (Calvillo García, Cornejo Oviedo, Valencia Mazo, & Flores López, 2005) without considering AGB production. Méndez-González, Luckie-Navarrete, Capó-Arteaga, and Nájera-Luna (2011) reported that maximum AGB production (79.2 Mg∙ha-1) occurred at 20-years old, with a CAI-MAI intersection point of 3.96 Mg∙ha⁻1∙year⁻1 in Pinus plantation. Although the predicted AGB production was similar in both studies, the managed forest studied herein reached its maximum AGB at a lower age (12-years old; Figure 3). The AGBI reached in managed stands (Table 1) showed that P. patula is a highly productive species (Table 4). Pinus patula is a fast-growing species with biological characteristics that enable its establishment in highly productive sites, where it can reach its maximum productive potential in a short time (Velázquez et al., 2004). However, it is important to consider the biological and economic rotations of stands, since a technical biomass rotation of 12 years (Figure 3) could be insufficient for the self-regulation and recovery of ecosystem functions after the harvesting process (Restrepo & Alviar, 2010), in order to make it sustainable. The rotation period that has been used in P. patula forests with forest management in Mexico is between 30 (Guillespie, 1992) and 40 years (Castelán-Lorenzo & Arteaga-Martínez, 2009).
It is important to know the pattern of biomass production during the ontogenetic development of the forest, because it not only provides information on the amount of wood produced, but also on the ecosystem services that can be recovered, such as the sequestration of CO2 implicit in the construction of forest biomass, which in parallel allows the regulation of climate, water cycle and nutrients. To some extent, these services may be governed by the intermediate silvicultural treatments applied in the stands, which also define whether the forest could be a potential C sink. Some studies have evaluated AGB in several temperate forests with forest harvesting in Europe (Kowalski et al., 2004; Valentini et al., 2000) and have revealed that the application of silvicultural treatments can change the forest from a C sink to a source after harvesting. However, this process is temporary and as soon as the vegetation of the stand is established, the process is reversed, although the time in which this process occurs has not been documented. The results of this study affirmed that a stand at the age of 7 years (1998 stand) stores approximately 8 % of C in its biomass respect to that stores NMF (Table 2), considering that 50 % of the biomass corresponds to the C fixed by photosynthesis (Penman et al., 2003). On this matter, Strukelj, Brais, and Paré (2015) reported that nine years after harvesting Populus tremuloides Michx. stands, clear-cut areas were a net C source, while those partially harvested were a net C sink, which demonstrates that a well-managed secondary forest can have higher production than primary forests and, therefore, sequester more atmospheric CO2. Active forest management in the forests of the study region allows for better control of AGB production, C stocks and diversification of harvesting prescriptions could favor C sequestration at the ecosystem and landscape levels (Mayer et al., 2020). In this way, both AGB production and ANPP could be considered as indicators of the managed forest capacity to C storage, as well as the intermediate and complementary silvicultural treatments can ensure timber productivity and C fixation by vegetation.
Conclusions
P. patula forests under forest management accelerate aboveground biomass (AGB) accumulation in temperate forest stands throughout the chronosequence, reaching similar levels to a non-managed forest (NMF) in a shorter period. The oldest stand (30-years old) after harvesting showed an AGB only 22 % lower than that in NMF. Silvicultural practices strongly influenced AGB production and allocation, which increased with stand age. These accumulated more than 70 % of AGB in stems, as a result of thinning, which aims to produce stems with characteristics suitable for long-lived wood products. As AGB, ANNP (aboveground net primary productivity) increased with stand age; however, the highest values were observed in NMF. Based on the results, it can be considered that sustainable forest management represents an alternative for climate change mitigation, through the conservation and increase of C reserves in forest ecosystems.

