










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias forestales
Print version ISSN 2007-1132
Rev. mex. de cienc. forestales vol.7 n.33 México Jan./Feb. 2016
Articles
Aboveground biomass of Zanthoxylum kellermanii P. Wilson in a tropical evergreen forest at the North of Oaxaca State
1Asesoría e Investigación para el Manejo de Ecosistemas Forestales Oaxaca, Oax., México.
2Postgrado Forestal, Colegio de Postgraduados. México.
3Posgrado Consultor independiente. Correo-e: elherrera2001@yahoo.com.mx
Zanthoxylum kellermanii is a species of ecological and economic value in southeast Mexico, in particular in the Chinantla region of Oaxaca State. In order to generate the equations that determine its aboveground biomass, two trees by diametric class (10, 15, 20 and 25 cm) were felled, dividing each of them into three sections: foliage, branches and stem. From each section samples were taken and weighed directly to determine fresh weight and later they were dried in kilns. With these data the relationship between fresh weight and dry weight was established to estimate values of biomass per tree compartments and the total for each category, adjusting allometric equations by nonlinear regression techniques. The values of biomass per tree ranged from 0.494 to 2.14 kg in foliage, 1.74 to 25.9 kg in branches and 7.6 to 206.72 kg in stems. The average proportion of the biomass was 83 % in stems, branches and 14 % in 3 % in foliage. Allometric equations generated for Z. kellermanii showed a good level of fit, determined by their statistical values, so that the model is reliable to make a good prediction of biomass through the normal diameter. The distribution pattern of biomass, relative to the total dry weight indicates that this species has the greatest potential to accumulate biomass in the stem.
Keywords: Aboveground biomass; allometric equations; foliage; stem; Oaxaca; branches
Zanthoxyllum kellermanii es una especie de importancia ecológica y económica del sureste mexicano, que se desarrolla en la región de La Chinantla, Oaxaca. Para generar las ecuaciones conducentes a determinar su biomasa aérea, se derribaron dos árboles por clase diamétrica (10, 15, 20 y 25 cm), de las que se separaron tres secciones: follaje, ramas y fuste. De cada una se obtuvieron muestras que se pesaron en fresco y que posteriormente fueron secadas en estufas. Con estos datos se estableció la relación entre peso fresco y peso seco para estimar valores de biomasa por componentes y del árbol total para cada clase, además se ajustaron ecuaciones alométricas mediante técnicas de regresión no lineal. Los valores de biomasa aérea por árbol variaron de 0.494 a 2.14 kg en follaje, de 1.74 a 25.9 kg en ramas y de 7.6 a 206.72 kg en fustes; la proporción promedio de la biomasa fue de 83 % en fustes, 14 % en ramas y 3 % en follaje. Las ecuaciones alométricas generadas para Z. kellermanii mostraron un buen nivel de ajuste, determinado por sus valores estadísticos, por lo que el modelo es confiable para realizar una buena predicción de la biomasa aérea a través del diámetro normal. El patrón de distribución de la biomasa, respecto al peso seco total indica que la especie tiene su mayor potencial para acumular biomasa en el fuste.
Palabras clave: Biomasa aérea; ecuaciones alométricas; follaje; fuste; Oaxaca; ramas
Introduction
Forest ecosystems have an important role in the carbon cycle because they store large amounts of this element in the vegetation and soil, to exchange it with the atmosphere through photosynthesis and respiration (Domínguez, 2011). These communities are real carbon sinks to form the structures of organisms and then partially converted to organic matter (Pardos, 2010), so that the distribution of biomass in the trees is crucial to the rate of accumulation of the latter and C flow (Chou and Gutiérrez, 2013).
The air component of trees is one of the main reservoirs of biomass; their production and their presence in the structures of the tree are two important characteristics to estimate it in forest species (Pardos, 2010). The ability of forests to store carbon in their biomass varies by species composition, age and density of the population in each stratum of the plant community (Laumonier et al., 2010).
The management of forest ecosystems with a focus on sustainability has led to the generation of allometric equations that estimate the biomass in different sections of the tree. This potential to interpret dimensional and volumetric variations has already been tested and used in plants for decades (Niklas, 1994; Enquist and Niklas, 2002) and its application has been useful in agriculture, and in the study of how some ecosystems and forests work (Soto et al., 2010; Chave et al., 2014).
These equations are useful in estimating carbon storage and the partial calculation of primary productivity of many types of ecosystems (Rodríguez et al., 2008; Návar, 2009; Acosta et al., 2011; Carrillo et al., 2014). Primary productivity is an indicator of carbon balance between vegetation and the atmosphere, as well as a variable of interest in research on global climate change (Schlesinger, 1997; Clark et al., 2001).
In the Mexican tropics this issue has been addressed in plantations of Hevea brasiliensis Muell. Arg (Monroy and Návar, 2004) and Bambusa oldhamii Munro (Castañeda et al., 2005), in agroforestry systems (Callo et al., 2004; Roncal et al., 2008; Casanova et al., 2011; Soto et al., 2010), in deciduous forests (Rodríguez et al., 2008; Návar et al., 2013), semi- deciduous (Aquino et al., 2015) and evergreen (Montero et al., 2008; Douterlungne et al., 2013). It highlights the fact that even though the forests have a high concentration of biomass, many species have not been fully explored yet.
The third largest forest area in the Mexican humid tropics is located at the north of Oaxaca State (De Teresa et al., 2009); it has been subjected to intense processes of disturbance that affect carbon dynamics such as deforestation and land use change. In this situation, it is necessary to quantify or to make an inventory of the current biomass and carbon reserves of these ecosystems. Zanthoxyllum kellermanii P. Wilson is an integral species of the secondary forests of La Chinantla, it is a valuable component of the ecological succession and it is prized for its timber (Manzano et al., 2010).
From the aforementioned, the aim of the actual study was to make an estimation of the total aboveground biomass, its location in the stem, branches and foliage in Zanthoxyllum kellermanii trees of a secondary tropical forest at the north of Oaxaca.
Materials and Methods
Study area
Field work was carried out in San Martín Soyolapan, Comaltepec municipality, Oaxaca State which is physiographically placed in the Papaloapan river basin and in the surroundings of the Sierra Norte de Oaxaca (17°41'53.5'' N y 96°16'57.3'' W) (Figure 1), at 160 m high.
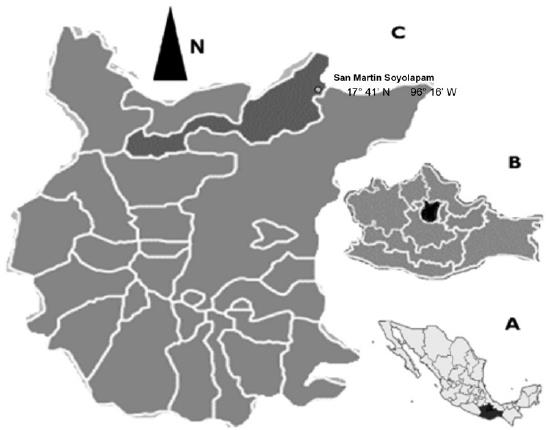
Figure 1 Study area. Location of Oaxaca State in Mexico (A), Sierra Norte Region (B), Santiago Comaltepec (C).
Climate is classified as Am(w)g, which is moist warm with summer rains, an average annual temperature, 24 °C and annual rainfall up to 3 590 mm (García, 1987). Geologically there can be found extrusive igneous rocky and limestone outcrops with the following soil types: yellow-reddish lateritic, yellow-reddish latosolic, karstic lythosoils, red loams and laterites (Inegi, 2011). The prevailing vegetation is the evergreen tropical forest where the following tree species are abundant: Andira galeotiana Standley, Brosimum alicastrum Sw., Dialium guianense (Aub) Sandw., Terminalia amazonia (J. F. Gmel.) Exell y Vochysia guatemalensis Donn. Sm. (Miranda and Hernández, 1963).
Species description
Zanthoxyllum kellermanii is a tree up to 10 to 25 m high and 80 cm in diameter. It has a brownish-grey bark. Pinnate leaves 20 to 50 cm long, 4 x 2 a 1.5 x 5.5 cm orbicular to elliptic shape leaflets displayed in a spiral. Dioecious species, the male inflorescences up to 30 cm long and the female inflorescences up to 10 cm, both in subterminal and axillary panicles. Capsular, triangle shape, brownish-black fruits 5 to 7 mm. It grows in the humid forests, at 50 to 700 m high, from southern Mexico up to Peru. It is part of the medium stratum of the evergreen tropical rainforest, particularly of Terminalia amazonia, over deep lateritic soils from ancient alluvial runoffs. Its wood has a beautiful grain which makes it attractive for sawing and it is used, as well, for rural building (Pennington and Sarukhán, 2005).
Construction of allometric equations
Two trees from each of the following categories were removed: 10, 15, 20 y 25 cm, as this sampling size has been used in different studies carried out in the tropics with good results (Brown et al., 1995). Before these trees were felled, their normal diameter (DN) was measured with a 5 m long Ben Meadows diametric tape. Once on the ground, they were divided into three sections: stem, branches and foliage.
Stems as well as branches were cut in 2 m lots and foliage was taken from the branches and packaged in 0.5 kg portions in plastic bags (Domínguez, 2011). Each part was placed in a separate way in a 500 kg Torrey balance to determine fresh weight in situ.
Five samples were obtained from each tree section of the diameter categories 10 and 15 cm, and 10 samples per section of each tree categories 20 and 25 cm. Samples of branches and stems were cross sections (slices) of 3 and 5 cm thick, respectively. The fresh weight of the samples was recorded in situ for all the sections with a pan scale (Oken) five kilograms of capacity and an approximation of five grams. All the samples were placed in plastic bags for transportation to the laboratory (Laboratorios Generales del Colegio de Postgraduados Campus Montecillo) in which control data were recorded (Figueroa et al., 2005; Domínguez, 2011).
In the laboratory, samples of branches and foliage were put into paper bags and then into drying kilns. Branch and leaves samples were dried at 75 °C and stems, at 80 °C., until constant weight was achieved. With these data the ratio of fresh and dry weight was determined to estimate the proportion of biomass components as well as the total aboveground, by using Equation 1 (Domínguez, 2011):
 1
1
Where:
PS = Dry weight
PF= Fresh weight
PScom = Dry wight of the tree section
PFcom = Fresh weight of the tree section
The biomass values by section of each tree that was sampled from the dry and fresh weight data were fitted with allometric equations by non -linear regression techniques. A potential model was fitted by the NLIN procedure of SAS 8.0 version (SAS, 1999). This model (Equation 2) has been widely used in different studies focused on tree aboveground biomass (Acosta et al., 2002; Figueroa et al., 2005; Carrillo et al., 2014):
 2
2
Where:
Y = Estimated weight of the tree section in kg
b0 and b1 = Regression coefficients
X = Normal diameter in cm
With the allometric equations graphics were made with an Excel sheet (Microsoft, 2003) for total aboveground biomass and of the tree sections.
The goodness of fit of this model was assessed through the statistical values of the analysis of variance: mean squared error (CME, for its acronym in Spanish), determination coefficient (R2) and significance level (p < 0.05).
Results and Discussion
Allometric equations
In Table 1 are shown the allometric equations formulated to estimate total aboveground biomass and of the tree sections, as well as their variation estimators and statistical reliability. All the equations showed a good fit (p < 0.0001) and both, the R2 and the mean squared error (CME) values point out that the model is reliable to estimate aboveground biomass from normal diameter.
Table 1 Allometric equations formulated to estimate total aboveground biomass and of the tree sections of Zanthoxylum kellermanii P. Wilson.
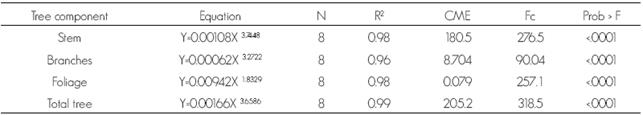
N = Sample size; R2 = Determination coefficient; CME = Mean squared error; Fc = calculated F; Y = Biomass (kg), X = normal diameter (cm).
The regression fitted curves to estimate aboveground biomass are shown in Figure 2 for tree components and the full tree.

Distribution of aboveground biomass
Aboveground biomass in each of the sections of Zanthoxylum kellermanii was organized as follows: in the shaft the range of values was between 77.50 % and 87.52 % in the diametric categories 10 and 25 cm; in the branches, it was 1.00 % to 17.52 % which referred to 25 and 10 cm, and the variation in foliage was from 1.47 % to 4.98 % in the same classes. The average proportion of biomass per section was 83.21 % in stem, from 13.57 % in branches and foliage of 3.22 % in, which includes all diameter categories (Table 2 and Figure 3).
Table 2 Dry weight (kg) and aboveground biomass (%) distribution in tree components for the different diametric categories of Zanthoxylum kellermanii P. Wilson.
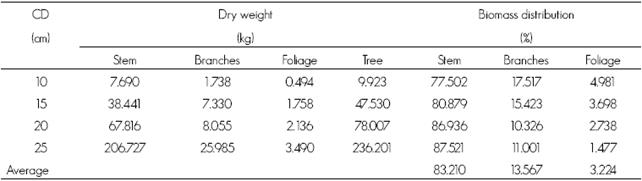
CD = Diametric category.
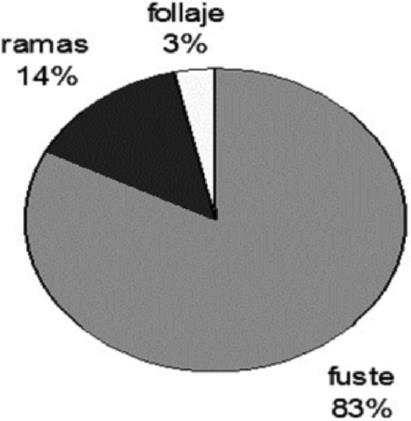
Figure 3 Proportional distribution of the aboveground biomass in each of the tree components of Zanthoxylum kellermanii P. Wilson
The dry weight in the different tree sections was distributed within the following ranges: foliage, 0.494 kg to 2.136 of kg; branches, 1.738 kg to 8.055 kg and stem 7.690-206.725 kg, for the 10 and 25 cm diameter categories in all cases, being the highest proportion of aboveground biomass in the stem and the lowest in the foliage. The values of total tree biomass range from 9.923 to 236.727 kg in the diameter categories of 10 and 25 cm (Table 2).
The results of the actual study reveal a similar behavior of the biomass to others from research make in the Mexican humid tropic, as it concentrates the greatest part in the stem (83.21 %). In this regard, Monroy and Návar (2004) determined 73.88 % in the whole stem and 27.09 % in branches in a Hevea brasiliensis plantation in Uxpanapa, Veracruz State; and Castañeda et al. (2005) of 83.7 % in stems, 12.3 % in foliage and 4 % in branches of Bambusa oldhamii Munro in Huatusco, Veracruz State.
Other studies about biomass of tropical rainforests species in Tabasco (Montero et al., 2008) and Costa Rica (Fonseca et al., 2009) confirm that a greater amount of biomass in the stem increases with age and the size of the tree, thus, some research studies only base their methodology in the volume of the trunk (Návar et al., 2013).
The biomass of branches recorded an intermediate (13.57 %) value relative to the biomass of stem and leaves. Goodman et al. (2014) mention that the first is determined by the size and architecture of the crown, and the quality of site and stand characteristics, among which the growing space is outstanding (Foli et al., 2003).
Foliage biomass recorded the lowest value in the distribution of magazines (3.22 %); as in branches, this sort of biomass was proportionately greater in trees with small diameters and gradually decreased with their increment. Small trees are dominated and have little light (López et al., 2006), suggesting a limiting condition by way of the Law of Least in which the amounts of resources available are very close to what is indispensable (Odum, 1972); and in this particular case it is related to the concept of growth deficiency, in which it is stated that the production of stem wood has a lower priority, compared to the production of foliage and roots.
Enquist et al. (1999) proved that the growth rate in tropical trees has a potential relationship with biomass. For this study, the species has diameter increases higher than 1.5 cm yr-1 favored by the glass with good quality (symmetrical, vigorous) and exposed both vertically and laterally to sunlight (Manzano et al., 2010). The same authors mention that being a species of early succession, it can be used in recovery programs and enrichment of disturbed areas in the study area.
Conclusions
The allometric equations from Zanthoxylum kellermanii showed a right goodness of fit, determined by their statistical values, which make the model a reliable option to make a correct prediction of the aboveground biomass through normal diameter.
The distribution pattern of biomass in regard to total dry weight shows that this species has its greatest potential to accumulate biomass in the stem.
Conflict of interests
The authors declare no conflict of interests.
Contribución by author
Filemón Manzano Méndez: field and office work and writing of the manuscript; Juan Ignacio Valdez Hernández: office work and writing of the manuscript; Miguel Ángel López López: field and office work and writing of the manuscript and Elizandro Pineda Herrera: office work, writing and review of the manuscript.
Acknowledgements
The authors would like to thank the community authority of Santiago Comaltepec for his support during this research, as well as to the authorities and people of San Martín Soyolapan. Also to the Ford Foundation for having sponsored the graduate studies of the first author. To the specialists that reviewed the document by assignment of the Revista Mexicana de Ciencias Forestales for their comments and suggestions to the original version.
REFERENCES
Acosta M., M., F. Carrillo A. y R. Gómez V. 2011. Estimación de biomasa y carbono en dos especies de bosque mesófilo de montaña. Revista Mexicana de Ciencias Agrícolas 2(4):529-543. [ Links ]
Acosta M., M., J. Vargas H., A. Velázquez M. y J. D. Etchevers B. 2002. Estimación de la biomasa aérea mediante el uso de relaciones alométricas en seis especies arbóreas en Oaxaca, México. Agrociencia 36(6):725-736. [ Links ]
Aquino R., M., A. Velázquez M., J. Castellanos B., H. de los Santos P. y J. Etchevers B. 2015. Partición de la biomasa aérea en tres especies arbóreas tropicales. Agrociencia 49(3):299-314. [ Links ]
Brown, I. F., L. Martinelli, W. W. Thomas, M. Moreira, C. Ferreira and R. Victoria. 1995. Uncertainty in the biomass of Amazonian forests: an example from Rondonia, Brazil. Forest Ecology and Management 75(1):175-189. [ Links ]
Callo C., D. A., I. B. Rajagophal y L. C. Krishnamurthy. 2004. Secuestro de carbono por sistemas agroforestales en Veracruz. Ciencia UANL 2(2):60-65. [ Links ]
Carrillo A., F., M. Acosta M., E. Flores A., J. Juárez B. y E. Bonilla P.2014. Estimación de biomasa y carbono en dos especies arbóreas en La Sierra Nevada, México. Revista Mexicana de Ciencias Agrícolas 5(5):779-793. [ Links ]
Casanova L., F., J. Petit A. y J. Solorio S. 2011. Los sistemas agroforestales como alternativa a la captura de carbono en el trópico mexicano. Revista Chapingo Serie Ciencias Forestales y del Ambiente 17(1):133-143. [ Links ]
Castañeda M., A., J. Vargas H. , A. Gómez G., J. I. Valdez H. y H. Vaquera H. 2005. Acumulación de carbono en la biomasa aérea de una plantación de Bambusa oldhamii. Agrociencia 39:107-16. [ Links ]
Chave, J., M. Réjou M., A. Búrquez, E. Chidumayo, M. Colgan, W. Delitti B.C., A. Duque, T. Eid, P.M. Fearnside, R. C. Goodman, M. Henry, A. Martínez-Yrízar, W. A. Mugasha, H. Muller-Landau, M. Mencuccini, B. W. Nelson, A. Ngomanda, E. M. Nogueira, E. Ortiz M., R. Pélissier, P. Ploton, C. M. Ryan, J. G. Saldarriaga and G. Vieilledent. 2014. Improved allometric models to estimate the aboveground biomass of tropical trees. Global Change Biology 20:3177-3190. [ Links ]
Chou, S. W. y E. Gutiérrez E. 2013. Ecuación para estimar la biomasa arbórea en los bosques tropicales de Costa Rica. Tecnología en Marcha 26(2):41-54. [ Links ]
Clark, D. A., S. Brown, D. W. Kickligghter, J. Q. Chambers, J. R. Thomlinson, J. Ni and E. A. Holland. 2001. NPP in tropical forest: an evaluation and synthesis of existing field data. Ecological Applications 1:371-384. [ Links ]
De Teresa, A. P., G. Hernández C., P. Legarreta H. y M. Orozco R. 2009. Quia-na. La selva chinanteca y sus pobladores. Ed. UAM-I-Juan Pablos editores, México, D. F., México. 13 p. [ Links ]
Domínguez C., C. 2011. Manual interdisciplinario de investigación forestal. Instituto Internacional de Dasonomía Tropical. USDA Forest Service. Río Piedras, Puerto Rico.168 p. [ Links ]
Douterlungne, D., A. M. Herrera G., B. G. Ferguson, I. Siddique y L. Soto P. 2013. Ecuaciones alométricas para estimar biomasa y carbono de cuatro especies leñosas neotropicales con potencial para restauración. Agrociencia 47(4):385-397. [ Links ]
Enquist, B. J., and K. Niklas. 2002. Global allocation rules for patterns of biomass partitioning in seed plants. Science 295(5559):1517-1520. [ Links ]
Enquist, B. J., G. V. West, E. L. Charnov and J. H. Brown. 1999. Allometric scaling of production and life-history variation vascular plants. Nature 401:907-91. [ Links ]
Figueroa N., C., J. Etchevers B., A. Velásquez M. y M. Acosta M. 2005. Concentración de carbono en diferentes tipos de vegetación de la Sierra Norte de Oaxaca. Terra Latinoamericana 23(1):57-64. [ Links ]
Foli, E. G., D. Alder, H. G. Miller and M. D. Swaine. 2003. Modelling growing space requirements for some tropical forest tree species. Forest Ecology and Management 173:79-88. [ Links ]
Fonseca, G. W., F. Alice G. y J. M. Rey B. 2009. Modelos para estimar la biomasa de especies nativas en plantaciones y bosques secundarios en la zona Caribe de Costa Rica. Bosque 30(1):36-47. [ Links ]
García, M. E. 1987. Modificaciones al sistema de clasificación climática de Köppen. Instituto de Geografía. Universidad Nacional Autónoma de México. México, D. F., México. 217 p. [ Links ]
Goodman, R. C., O. Phillips L. and T. R. Baker. 2014. The importance of crown dimensions to improve tropical tree biomass estimates. Ecological Applications 24(4):680-698. [ Links ]
Instituto Nacional de Estadística, Geografía e Informática (Inegi). 2011. Conjunto de datos vectoriales de la carta de uso del suelo y vegetación: escala 1:250 000. Serie III (continuo nacional). Aguascalientes, Ags., México. s/p. [ Links ]
Laumonier, Y., A. Edin, M. Kanninen and A. W. Munandar. 2010. Landscape- scale variation in the structure and biomass of the hill dipterocarp forest of Sumatra: implications for carbon stock assessments. Forest Ecology and Management 259:505-513. [ Links ]
López L., M. A., A. Velásquez M., J. Acosta M. and E. Estañol B. 2006. Biomass distribution in declining Sacred-fir seedlings. Interciencia 31(6):451-455. [ Links ]
Manzano, F., J. I. Valdez H., M. A. López L. y H. Vaquera H. 2010. Crecimiento en diámetro de Zanthoxylum kellermanii P. Wilson en una selva perennifolia del norte de Oaxaca, México. Madera y Bosques 16(2):19-33. [ Links ]
Microsoft, 2003. Microsoft Excell for Windows. (Version 2003). Redmond, WA, USA. n/p. [ Links ]
Miranda, F. y E. Hernández X. 1963. Los tipos de vegetación de México. Boletín de la Sociedad Botánica de México 28:29-179. [ Links ]
Monroy, R. C. y J. J. Návar Ch. 2004. Ecuaciones de aditividad para estimar componentes de biomasa de Hevea brasiliensis Muell. Arg. en Veracruz, México. Madera y Bosques 10(2):29-43. [ Links ]
Montero G., N., O. Castillo A. y J. L. Martínez S. 2008. Captura de carbono en un remanente de Selva Alta Perennifolia en el Ejido Niños Héroes, Tenosique, Tabasco. Kuxulkab 14(26):45-50. [ Links ]
Návar Ch., J. J. 2009. Allometric equations and expansion factors for tropical dry trees of eastern Sinaloa, Mexico. Tropical and Subtropical Agroecosystems. 10:45-52. [ Links ]
Návar Ch., J., F. J. Rodríguez F. y P. A. Domínguez C. 2013. Ecuaciones alométricas para árboles tropicales: Aplicación al inventario forestal de Sinaloa, México. Agronomía Mesoamericana. 24(2):347-356. [ Links ]
Niklas, K. J. 1994. Plant allometry: the scaling of form and process. University of Chicago Press. Chicago, IL, USA. 412 p. [ Links ]
Odum, E. P. 1972. Ecología. Nueva editorial Interamericana. México, D.F., México. 653 p. [ Links ]
Pardos, J. A. 2010. Los ecosistemas forestales y el secuestro de carbono ante el calentamiento global. Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (INIA). Ministerio de Ciencia e Innovación Madrid, España. 253 p. [ Links ]
Pennington, T. D. y J. Sarukhán K. 2005. Árboles tropicales de México: manual para la identificación de las principales especies. Fondo de Cultura Económica. Universidad Nacional Autónoma de México. México, D. F., México. 523 p. [ Links ]
Rodríguez L., R., J. Jiménez P., J. Meza R., O. Aguirre C. y R. Razo Z. 2008. Carbono contenido en un bosque tropical subcaducifolio en la reserva de la biosfera el cielo, Tamaulipas, México. Revista Latinoamericana de Recursos Naturales 4 (2):215-222 [ Links ]
Roncal G., S., L. Soto P., J. Castellanos A., N. Ramírez M. y B. de Jong. 2008. Sistemas agroforestales y almacenamiento de carbono en comunidades indígenas de Chiapas, México. Interciencia 33(3):200-206. [ Links ]
Schlesinger, W. H. 1997. Biogeochemistry: an analysis of global change. Academic Press, San Diego, CA, USA. 588 p. [ Links ]
Soto Pinto, L., M. Anzueto, J. Mendoza, G. Ferrer and B. de Jong. 2010. Carbon sequestration through agroforestry in indigenous communities of Chiapas, Mexico. Agroforestry Systems 78:39-51. [ Links ]
Statistical Analytical System (SAS). 1999. User's guide, version 8.0 for Windows SAS Institute Inc. Cary, NC, USA. 595 p. [ Links ]
Received: December 30, 2014; Accepted: January 01, 2016
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
