










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias forestales
versión impresa ISSN 2007-1132
Rev. mex. de cienc. forestales vol.15 no.81 México ene./feb. 2024 Epub 13-Mayo-2024
https://doi.org/10.29298/rmcf.v15i81.1435
Scientific article
Differences in the response of dendrocronological indicators to climatic and topographic conditions
1Universidad Autónoma de Nuevo León, Facultad de Ciencias Forestales. México.
Vegetation dynamics in forest ecosystems are commonly attributed to climatic factors. However, there is still limited knowledge on how topography influences the climatic sensitivity of forest species. The response of dendroecological indicators of Pinus arizonica to exposure and climatic variability was determined. Indices of ring width (RWI) and Basal area increments (BAI) were obtained. The correlation between RWI and monthly records of accumulated precipitation (PP) and mean temperatures (Tmax and Tmin) was calculated for three periods: 1990-2000 (B1), 2001-2011 (B2), and 2012-2021 (B3). Mann-Whitney and Kruskall-Wallis tests were performed to test for differences in BAI by exposure (northwest, NW and northeast, NE) and by time period, respectively. Tmax limited growth in all three periods, Tmin restricted it in B2 and PP favored it in B1 and B2. A higher BAI was detected in the NW exposure (224.7 mm2 yr-1) compared to the NE (143.9 mm2 yr-1). There was a significant increase in BAI in periods B2 (62.1 mm2 yr-1) and B3 (56.9 mm2 yr-1) compared to B1 (51.2 mm2 yr-1). This highlights the importance of considering topography when analyzing climate-growth associations of forest species. Considering the interaction of climatic factors and microclimates associated with site-specific topography generates an integrated view of ecological processes. This allows proposing better forest management strategies to mitigate the effects of Climate Change.
Keywords Basal area increment; dendroecology; dplR; growth rings; topographic exposure; Pinus arizonica Engelm
La dinámica de la vegetación en ecosistemas forestales, comúnmente, se atribuye a factores climáticos. Sin embargo, aún existe un conocimiento limitado sobre cómo la topografía influye en la sensibilidad climática de especies forestales. Se determinó la respuesta de indicadores dendroecológicos de Pinus arizonica a la exposición y la variabilidad climática. Se obtuvieron índices de ancho de anillo (IAA) e incrementos de área basal (IAB). Se calculó la correlación entre los IAA, registros mensuales de precipitación acumulada (PP) y temperaturas medias (Tmáx y Tmín) en tres periodos: 1990-2000 (B1), 2001-2011 (B2) y 2012-2021 (B3). Se realizaron las pruebas de Mann-Whitney y de Kruskall-Wallis para comprobar diferencias en el IAB por exposición (noroeste, NO y noreste, NE) y por bloque temporal, respectivamente. La Tmáx limitó el crecimiento en los tres periodos, la Tmín lo restringió en el B2 y la PP lo favoreció en los B1 y B2. Se detectó un mayor IAB en la exposición NO (224.7 mm2 año-1) en comparación con la NE (143.9 mm2 año-1). Existió un aumento significativo en el IAB en los periodos B2 (62.1 mm2 año-1) y B3 (56.9 mm2 año-1) en comparación con el B1 (51.2 mm2 año-1). Esto resalta la importancia de considerar la topografía al analizar las asociaciones clima-crecimiento de especies forestales. Considerar la interacción de factores climáticos y microclimas asociados a la topografía específica del sitio, genera una visión integral de los procesos ecológicos, lo que permite proponer mejores estrategias de manejo forestal para la mitigación de los efectos del Cambio Climático.
Palabras claves Incremento de área basal; dendroecología; dplR; anillos de crecimiento; exposición topográfica; Pinus arizonica Engelm
Introduction
Climate variability plays a fundamental role in the distribution and development of forest species (Diao et al., 2023). In addition, Climate Change effects have been observed that have led to the deterioration of growth and even the decline of certain species in various forest ecosystems (Allen et al., 2010). The increase in the severity and intensity of droughts, related to the increase in global average temperature (IPCC, 2023), is a significant phenomenon. Therefore, understanding the impact of climatic conditions on the development of plant communities, is essential to unravel the mechanisms of vulnerability and adaptation of species.
The study of the relationship between climate and growth has been addressed through indicators based on growth rings (Acosta-Hernández et al., 2017). Disparities in the response of ring width indices (RWI) have been identified both among species coexisting in the same habitat (González-Cásares et al., 2017) and among those established in different geographic locations (Pompa-García et al., 2017). These differentiated responses are attributed to microenvironmental conditions, where topography, slope, and exposure emerge as important variables (Urrutia-Jalabert et al., 2021). Thus, heterogeneous landforms generate microhabitats characterized by specific environmental conditions, even in areas with a similar local climate. Elements such as elevation, slope and exposure play a crucial role in generating conditions that influence the establishment and growth of forest species, which affect the moisture, sunlight availability, and wind exposure, among other factors (Resler et al., 2005; Sundqvist et al., 2013; Pyatt et al., 2016).
In recent years, there has been an increased interest in the ecology of forest ecosystems in Mexico, and dendroecological approaches have been adopted as a means of study (Acosta-Hernández et al., 2017). Much research has inquired into the impact of climatic conditions and droughts on radial growth in various species (González-Cásares et al., 2017). However, within the national context, there is still a gap in the study of the influence of topographic variables on dendroecological indicators, such as ring width indices or the increase in basal area derived from the appearance of growth rings, and how they respond to climatic variables.
Therefore, this study analyzes the response of dendrochronological indicators such as ring width index and annual basal area increment to climatic and topographic conditions. Two objectives are established: (1) To evaluate the response of the ring width index to monthly precipitation and temperature variables, exploring potential variations in this response in 10-year intervals, and (2) To assess the impact of topographic exposure on basal area increment in trees of Pinus arizonica Engelm. var. stormiae Martínez, a species with limited distribution in an arid area of the Eastern Sierra Madre. Variations in dendrochronological indicators are related to monthly precipitation and temperature variables. In addition, it is hypothesized that topographic exposure will have an impact on the increase in the basal area of Pinus arizonica trees.
Materials and Methods
Study area and species
The study site known as Río de San José is located in the Eastern Sierra Madre, specifically in the municipality of Galeana, Nuevo León (Figure 1).
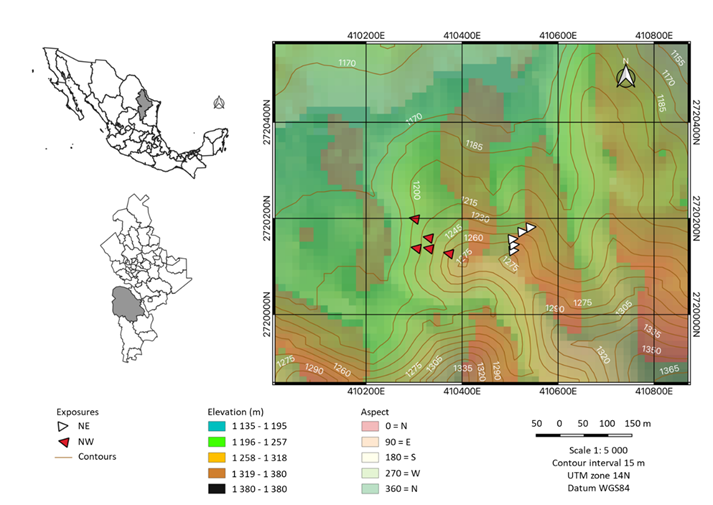
Figure 1 Location of the study area in the ejido Río de San José, in the municipality of Galeana, Nuevo León, Mexico.
The predominant climate is semi-dry, semi-warm, characterized by cool winters. The mean annual temperature is 21.3 °C, the accumulated precipitation reaches 663 mm per year, with 58 % of this total recorded between the months of June and September (Figure 2) (Harris et al., 2020). The predominant soil type in this region is Lithosol, characterized by its stony and poorly developed nature with a large amount of calcareous material (Schad, 2023).
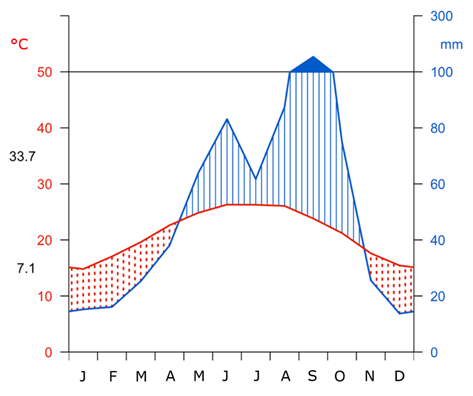
Río de San José (1 240 m). 1990 to 2022. 21.3 °C, 660 mm.
Figure 2 Walter-Lieth climate graph for precipitation and mean monthly temperature for the period 1990-2022.
The study species was Pinus arizonica var. stormiae. This consists of medium-sized trees, 15 to 20 m tall. It forms pure stands or is commonly found mixed with Quercus L. species, Pinus engelmannii Carrière and P. strobiformis Engelm. It grows in moderately dry to mesic forests, with chalky soils and at altitudes ranging between 1 500 and 2 800 masl (Farjon et al., 1997). It is mainly distributed in southern Nuevo León, as well as in some localities of southern Coahuila, Zacatecas, and San Luis Potosí (Farjon et al., 1997).
Sampling and sample preparation
Two sites were selected with different northeast (NE) and northwest (NW) exposures. Five trees were selected from each site (Figure 1) with a normal diameter >20 cm (caliper, Haglöff® Mantax Blue), good vigor, a long-standing appearance, and a straight, cylindrical stem without bifurcations, with the least possible disturbance. The coordinates (GPS, Garmin eTrex® 25x) of each tree were recorded, and two growth cores were obtained for each tree with a Pressler increment borer (Haglöff®, 5 mm) in opposite directions, always perpendicularly to the slope of the terrain (González-Tagle et al., 2023). Radial increment cores were attached to wooden guides with white glue and allowed to dry at room temperature. Subsequently, they were sanded with sandpaper of successively finer grain (100:1 000) (González-Tagle et al., 2023).
Chronology of ring width and basal area increment
The samples were visually dated using a stereomicroscope (AmScopeTM SM-4TPZ-B). Growth patterns were identified, and each ring was assigned its year of formation (Fritts, 2001). The ring width was measured using the VELMEX® measuring system (0.001 mm) and the Measure J2X software V5.03 (VoorTech Consoulting, 2021). The quality of the visual dating was statistically evaluated by comparing the individual tree series with the master chronology of the species. This procedure was carried out with the COFECHA software (Grissino-Mayer, 2001). A residual ring width chronology of Pinus arizonica was constructed. The ring width series were standardized to remove non-climate associated growth trends such as tree geometry and age to obtain ring width indices (RWI). An autoregressive model was applied to the generated indexes (Bunn, 2008). Subsequently, the residual ring width chronology was calculated, using a robust average to determine the averages of the individual indices. Residual ring width indices were used to remove most of the temporal autocorrelation related to the previous year's growth (Cook and Holmes, 1986).
The basal area increment (BAI) of Pinus arizonica was calculated using the following formula (Bunn, 2008):
Where:
R = Radius of the tree without bark at the height where the core was extracted (1.3 m)
t = Year of ring formation
First, the BAI was obtained at the sample (core) level, and then the annual average BAI was calculated for each tree. The construction of the chronology and the calculation of the BAI was done with the dplR (Dendrochronology Program Library in R) package (Bunn, 2008) of the R statistical software (R Core Team, 2021).
Climatic data
Monthly climate data were obtained: mean, maximum, and minimum temperature, and total precipitation. The data were downloaded from the Climate Research Unit at http://climexp.knmi.nl/ (Harris et al., 2020). Correlation coefficients were calculated between climate data and the residual chronology of the species for the 1920-2021 period, in order to evaluate the variation of the species radial growth sensitivity as a function of time for 10-year time windows: 1990-2000 (B1), 2001-2011 (B2), and 2012-2021 (B3).
Statistical analysis
The Mann-Whitney U test was utilized to determine whether there were significant differences in basal area increment (BAI) between the northeast and northwest exposures (Rubio-Cuadrado et al., 2021). The Kruskall-Wallis test and a Dunn’s multiple comparison test (Hood et al., 2018) were applied to assess the existence of significant differences between time periods. The choice of these analyses was based on the nature of the data, which did not show a normal distribution. Both were performed with the R-package ggstatsplot (Patil, 2021).
Results
Climate-radial growth association
The ring width chronology (Figure 3) shows the growth pattern of the 10 Pinus arizonica trees. The correlation between segments (0.38) was higher than the 0.328 (p>0.01) required by COFECHA, and the expressed population signal (EPS) obtained was 0.88. The first order self-correlation was 0.49, and the signal to noise ratio was 7.85. A low RWI is observed in 1996, 1998-1999, 2008-2009, 2019, and 2021, while high RWI is observed in 1993, 2007, 2010, and 2016.
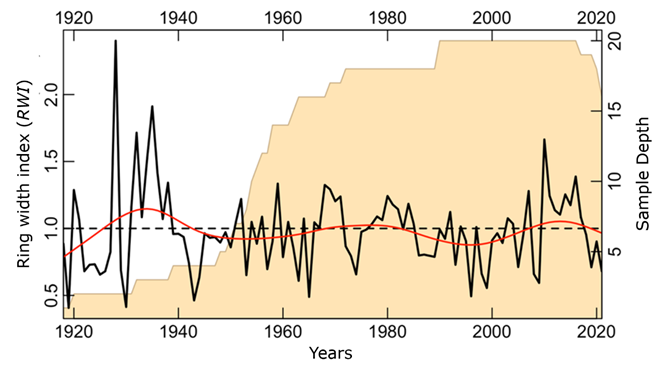
The solid line represents the ring width index.
Figure 3 Ring width chronology of Pinus arizonica Engelm. var. stormiae Martínez for the 1920-to-2021 period.
During the 1990-2021 period, significant correlations were found between climate variables and RWI. Precipitation in December prior to the growing year and in January and in April in the growing year were positively and significantly correlated with the RWI. The maximum temperature in December prior to the growing year and in January, February, and April in the growing year were negatively and significantly correlated with the RWI (Figure 4).
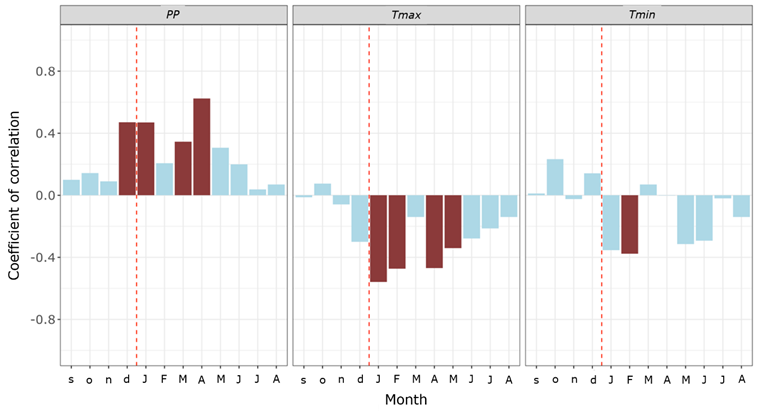
PP = Accumulated monthly precipitation; Tmax = Average maximum temperature; Tmin = Average minimum temperature. Significant correlations (95 %) are represented by red bars. Months in lower case correspond to the previous year of growth, and those in uppercase, to the current year of growth.
Figure 4 Growth-climate associations for the 1990-2021 period.
Correlation analyses reveal significant temporal variations (Figure 5). Between 1990-2000, precipitation in January of the year of growth had a positive and significant influence on the RWI. In the following ten years, the RWI correlated positively and significantly with the precipitation of June in the year of growth.
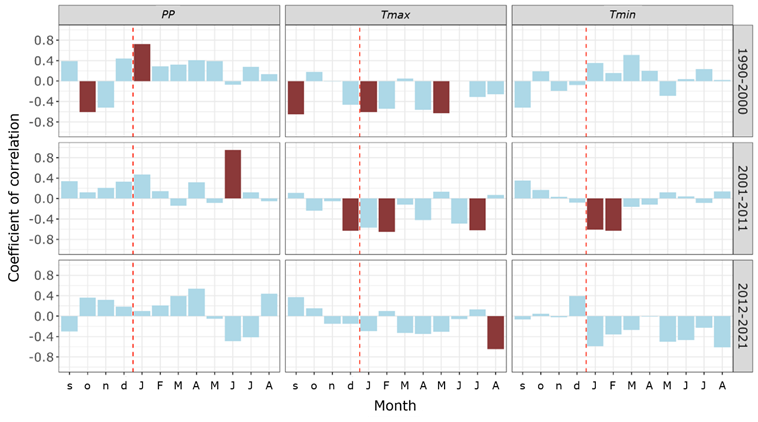
PP = Accumulated monthly precipitation; Tmax = Average maximum temperature; Tmin = Average minimum temperature. Significant correlations (95 %) are represented by red bars. Months in lower case correspond to the previous year of growth, and those in uppercase, to the current year of growth.
Figure 5 Growth-climate associations between 1990-2000 (B1), 2001-2011 (B2), and 2012-2021 (B3).
The RWI responds negatively and significantly to the Tmax of different months between periods. For the 1990-2000 period, it was to the Tmax for January and May of the year of growth. Between 2001-2011, to the Tmax of the previous December, as well as to the Tmax of February and July of the year of growth. In the period 2012-2021, to the Tmax of August of the year of growth. Tmin only negatively and significantly affected the RWI in the 2001-2011 period, particularly, the Tmin for January and February of the year of growth.
Effect of exposure on basal area increment
The Mann-Whitney U analysis revealed statistically significant differences in basal area increment between exposures (p<0.001) for the 1990-2021 period (Figure 6). Pinus arizonica trees with a northwest exposure showed a significantly higher mean BAI (224.7 mm2 yr-1) compared to trees exposed in the northeast direction (143.9 mm2 yr-1).
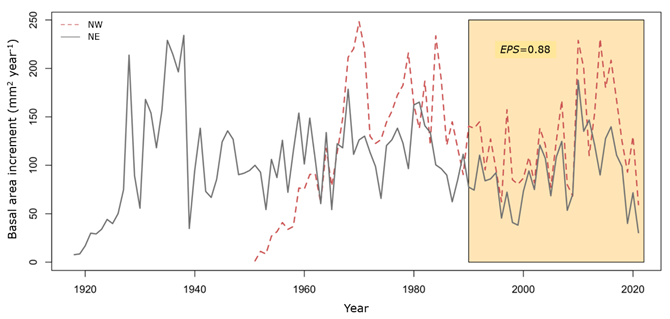
The yellow box shows the analyzed 1990-2021 period.
Figure 6 Comparison of the basal area increment (BAI) of Pinus arizonica Engelm. var. stormiae Martínez trees between the northeast (NE) and northwest (NW) exposures.
The Kruskall-Wallis test showed significant differences in the basal area increment between growth time periods (p<0.001) (Figure 7). Dunn's multiple comparison test indicated that the BAI in the 2001-2011 (B2, 62.1 mm2 yr-1) and 2012-2021 periods (B3, 56.9 mm2 yr-1) did not show a significant difference between each other (p>0.05). However, both B2 and B3 exhibited significant differences (p<0.05) in BAI compared to the 1990-2000 period (B1, 51.2 mm2 yr-1).
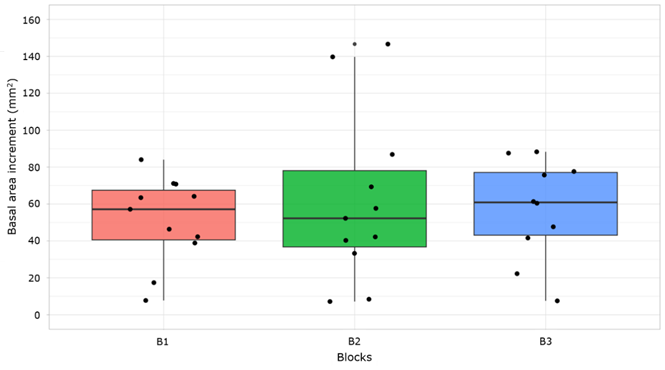
B1 = 1990-2000; B2 = 2001-2011; B3 = 2012-2021.
Figure 7 Comparison between basal area increment (BAI) of Pinus arizonica Engelm. var. stormiae Martínez trees between growth time periods.
Years that registered low RWI (Figure 3) agree with low site records of the Palmer drought index (PDSI) (Stahle et al., 2016). They also coincided with the lowest records of the standardized precipitation-evapotranspiration index (SPEI) (Vicente-Serrano et al., 2010) and the superposed epoch analysis (SEA) (Figure 8).

Discussion
This study researched, the influence of monthly climatic variables such as temperature and precipitation on the radial growth of Pinus arizonica in a semi-arid region of the Eastern Sierra Madre. We also evaluated whether this response varied at different 10-year intervals. The basal area increment (BAI) was examined in order to understand forest productivity in specific topographic exposures (northeast and northwest). These analyses enrich our understanding of the sensitivity and adaptability of this species to climatic conditions.
The importance of these findings lies in their potential to improve our understanding of the ecology of Pinus arizonica, especially within the context of mitigating the projected effects of Climate Change and their potential implications, such as an increase in the mean global temperature (IPCC, 2023).
Climate-radial growth association
In this study, a significant correlation was found between the ring width index (RWI) of Pinus arizonica and climatic conditions, specifically precipitation and maximum temperature. A larger radial growth of the species was related to higher levels of precipitation in the months of December prior to the growth year and January and April of the growth year, while the maximum temperature in those same months and February reduced the RWI (González-Cásares et al., 2017).
In the Western Sierra Madre, Pinus arizonica was found to be highly sensitive to climatic variables, particularly to the maximum temperature over a longer period, from the previous October to June of growth. Also, the minimum temperature in April was shown to have a significant limiting effect on the growth of this species (González-Cásares et al., 2017). These differences in sensitivity are attributed to both the specific characteristics of the variety studied and the conditions of the site, which is characterized as a dry area throughout the year (Harris et al., 2020).
In addition, the ring width indices of this species in northwestern Mexico has been found to exhibit a close connection with large-scale circulatory phenomena such as El Niño-Southern Oscillation (ENSO), the North Pacific Oscillation (NPO), and El Niño in region 3.4 (González-Cásares et al., 2017).
Furthermore, differences were detected in the response of the RWI to climatic variables by time periods. During the 1990-2000 period, the January precipitation significantly influenced radial growth, while in the 2001-2011 period, it was the June precipitation that had a significant impact due to the occurrence of hurricanes and tropical storms in those years (NHC and CPH, 2023).
As for maximum temperatures, a significant decrease in the radial growth of Pinus arizonica was observed during the 1990-2000 and 2001-2011 periods in response to three-month maximum temperatures. However, during the last study period (2012-2021), only the maximum temperature in August had a limiting effect on the RWI, indicating that the increase in temperature is affecting the growth of the species. On the other hand, the minimum temperature exerted a significant influence only during the period 2001-2011, exhibiting a negative association between the minimum temperature in January and February and the RWI of Pinus arizonica. These variations in radial growth sensitivity were related to changes in local temperatures over the three periods (Harris et al., 2020).
Effect of exposure and time periods on the BAI
A greater basal area increment (BAI) occurs in response to topographic exposure; such behavior is documented in several studies as a consequence of the complex interaction of such factors as solar radiation and microclimate, which together generate diverse microhabitats (Vitali et al., 2017). These microsites harbor conditions that result in differences in the basal area increment of the species. Elements such as temperature and the availability of water and nutrients (Lévesque et al., 2016) impact dendrochronological indicators. However, the genus Pinus can develop well in poor and dry sites. It is important to consider the soil composition of the Regosol, the stoniness, and the low water storage capacity (Eilmann et al., 2013). These conditions are accentuated by the topographic variation.
Different exposures result in specific conditions in limited geographical spaces. Although elevation and slope were not addressed in this research, they are topographic variables that influence radial climate-growth relationships (Pompa-García et al., 2017). Moreover, another study conducted in a dry temperate forest with conifers documented that the heterogeneity and sparse vegetation in this type of forest increases soil temperature, favoring a high rate of evapotranspiration (Breshears et al., 1998). Comparatively, the area in the Río de San José ejido region is sparsely vegetated and has a high spatial heterogeneity of individuals. These factors can lead to variations in tree growth and BAI across different topographic exposures within the study area.
A significant increase in the BAI of Pinus arizonica trees was observed in time periods B2 and B3 compared to period B1. This increase in BAI could be related to changes in environmental conditions during these periods. This is reflected in the climatic variables, especially during the 1990-2000 period, when less precipitation was documented in contrast to the other two time segments. In that decade, a mean annual precipitation of 556 mm was recorded, i. e., 159 and 167 mm less than during the periods B2 and B3, respectively. Furthermore, it was the only period that presented a dry summer month (Figure 7) (Harris et al., 2020).
Limitations and prospects
The limited distribution of a vulnerable species in a drought-prone area reduced the availability of trees for sampling. Consequently, the limited number of trees analyzed could compromise the adequate representation of the intrinsic variability of the population. This implies that the small sample may not fully encompass the diversity of individual responses to climatic and topographic conditions. Moreover, the ability to identify temporal patterns and trends across time periods may be compromised with a reduced number of trees. This could affect the ability of the study to detect short-term responses to climatic events or disturbances. In order to strengthen the robustness of future analyses, it is recommended to increase the number of trees sampled, considering various exposures, as well as to evaluate other environmental variables that may influence the growth of the species, such as drought indices. However, it is important to note that, despite the limitations in the number of trees sampled, this research contributes significantly to the general knowledge of the species.
Conclusions
This study confirms the premise of the significant impact of climatic conditions and exposure on the ring width index and basal area increment, respectively. A difference in the increase in the basal area between time periods is evident. The growth of Pinus arizonica showed differences in the response to climatic variations over three decades. This allows us to discern how the basal area increment responds to changing temperature and precipitation conditions. These findings reinforce the importance of considering the topography when assessing the growth dynamics within the context of Climate Change.
The use of these dendrochronological techniques is essential to understand the behavior of the species under changing climatic conditions and, therefore, to provide valuable information for forest management and evaluation of forest resources in the study area. We suggest exploring the applicability of this approach in different geographic regions and with different tree species, in order to improve our understanding of the response of dendrochronological indicators to the influences of climate and topography in an arid environment. Furthermore, this study enriches our knowledge of the ecology of this species in relation to topographic exposure and climatic conditions. These two factors must be taken into account in forest species management strategies, especially within the context of adaptation to the effects of climate change.
Acknowledgments
The authors are grateful to Azeneth García for her support in the collection and extraction of the increment cores in the field, as well as in the measurement of the rings in the Laboratory of Dendrochronology of the Graduate School Forest Sciences of the Autonomous University of Nuevo León. Marcos González Cásares thanks Conacyt for the grant awarded by the Postdoctoral Fellowships for Mexico 2022 program.
REFERENCES
Acosta-Hernández, A. C., M. Pompa-García and J. J. Camarero. 2017. An updated review of dendrochronological investigations in Mexico, a megadiverse country with a high potential for Tree-Ring Sciences. Forests 8(5):160. Doi: 10.3390/f8050160. [ Links ]
Allen, C. D., A. K. Macalady, H. Chenchouni, D. Bachelet, … and N. Cobb. 2010. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecology and Management 259(4):660-684. Doi: 10.1016/j.foreco.2009.09.001. [ Links ]
Breshears, D. D., J. W. Nyhan, C. E. Heil and B. P. Wilcox. 1998. Effects of woody plants on microclimate in a semiarid woodland: Soil temperature and evaporation in canopy and intercanopy patches. International Journal of Plant Sciences 159(6):1010-1017. Doi: 10.1086/314083. [ Links ]
Bunn, A. G. 2008. A dendrochronology program library in R (dplR). Dendrochronologia 26(2):115-124. Doi: 10.1016/j.dendro.2008.01.002. [ Links ]
Cook, E. R. and R. L. Holmes. 1986. Appendix 2: Users manual for program ARSTAN. In: Holmes, R. L., R. K. Adams and H. C. Fritts. Tree-ring chronologies of western North America: California, eastern Oregon and northern Great Basin with procedures used in the chronology development work including users manuals for computer programs COFECHA and ARSTAN. Laboratory of Tree-Ring Research, University of Arizona. Tucson, AZ, United States of America. pp. 50-65. [ Links ]
Diao, Y., S. Zhang, Y. Liu, G. Jin, S. Tian and Y. Liu. 2023. Effects of topography on radial growth of tree species with different mycorrhizal types. Forests 14(3):546. Doi: 10.3390/f14030546. [ Links ]
Eilmann, B., M. Dobbertin and A. Rigling. 2013. Growth response of Scots pine with different crown transparency status to drought release. Annals of Forest Science 70:685-693. Doi: 10.1007/s13595-013-0310-z. [ Links ]
Farjon, A., J. A. Pérez de la Rosa and B. T. Styles. 1997. A field guide to the pines of Mexico and Central America. Royal Botanic Gardens. Richmond, LDN, England. 147 p. [ Links ]
Fritts, H. C. 2001. Tree rings and climate. Blackburn Press. Caldwell, NJ, United States of America. 567 p. [ Links ]
González-Cásares, M., M. Pompa-García and J. J. Camarero. 2017. Differences in climate-growth relationship indicate diverse drought tolerances among five pine species coexisting in Northwestern Mexico. Trees 31:531-544. Doi: 10.1007/s00468-016-1488-0. [ Links ]
González-Tagle, M. A., J. Cerano-Paredes, W. Himmelsbach, E. Alanís-Rodríguez and Á. A. Colazo-Ayala. 2023. Fire records based on dendrochronological techniques for a coniferous forest in the southeastern region of Jalisco, Mexico. Revista Chapingo Serie Ciencias Forestales y del Ambiente 29(1):35-50. Doi: 10.5154/r.rchscfa.2022.03.018. [ Links ]
Grissino-Mayer, H. D. 2001. Evaluating crossdating accuracy: A manual and tutorial for the computer program COFECHA. Tree-Ring Research 57(2):205-221. https://www.ltrr.arizona.edu/~ellisqm/outgoing/dendroecology2014/readings/Grissino_mayer_COFECHA_2001.pdf . (20 de septiembre de 2023). [ Links ]
Harris, I., T. J. Osborn, P. Jones and D. Lister. 2020. Version 4 of the CRU TS monthly high-resolution gridded multivariate climate dataset. Scientific Data 7:1-18. Doi: 10.1038/s41597-020-0453-3. [ Links ]
Hood, S. M., D. R. Cluck, B. E. Jones and S. Pinnell. 2018. Radial and stand-level thinning treatments: 15-year growth response of legacy ponderosa and Jeffrey pine trees. Restoration Ecology 26(5):813-819. Doi: 10.1111/rec.12638. [ Links ]
Intergovernmental Panel on Climate Change (IPCC). 2023. Climate Change 2023: Synthesis Report, Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. IPCC, World Meteorological Organization (WMO) and United Nations Environment Programme (UNEP). Geneva, GE, Switzerland. 184 p. [ Links ]
Lévesque, M., L. Walthert and P. Weber. 2016. Soil nutrients influence growth response of temperate tree species to drought. Journal of Ecology 104(2):377-387. Doi: 10.1111/1365-2745.12519. [ Links ]
National Hurricane Center (NHC) and Central Pacific Hurricane Center (CPHC). 2023. NHC Data Archive, Tropical Cyclone Reports (TCRs). National Oceanic and Atmospheric Administration (NOOA). https://www.nhc.noaa.gov/data/#tcr . (20 de septiembre de 2023). [ Links ]
Patil, I. 2021. statsExpressions: R Package for Tidy Dataframes and Expressions with Statistical Details. Journal of Open Source Software 6(61):3236. Doi: 10.21105/joss.03236. [ Links ]
Pompa-García, M., M. González-Cásares, A. C. Acosta-Hernández, J. J. Camarero and M. Rodríguez-Catón. 2017. Drought influence over radial growth of Mexican conifers inhabiting mesic and xeric sites. Forests 8(5):175. Doi: 10.3390/f8050175. [ Links ]
Pyatt, J. C., D. F. Tomback, S. C. Blakeslee, M. B. Wunder, … and H. D. Bevency. 2016. The importance of conifers for facilitation at treeline: Comparing biophysical characteristics of leeward microsites in Whitebark Pine Communities. Arctic, Antarctic, and Alpine Research 48(2):427-444. Doi: 10.1657/AAAR0015-055. [ Links ]
R Core Team. 2021. R: A language and environment for statistical computing (version 4.0.3). Vienna, W, Austria. R Foundation for Statistical Computing. [ Links ]
Resler, L. M., D. R. Butler and G. P. Malanson. 2005. Topographic shelter and conifer establishment and mortality in an alpine environment, Glacier National Park, Montana. Physical Geography 26(2):112-125. Doi: 10.2747/0272-3646.26.2.112. [ Links ]
Rubio-Cuadrado, Á., C. Gómez, J. Rodríguez-Calcerrada, R. Perea, … and L. Gil. 2021. Differential response of oak and beech to late frost damage: an integrated analysis from organ to forest. Agricultural and Forest Meteorology 297:108243. Doi: 10.1016/j.agrformet.2020.108243. [ Links ]
Schad, P. 2023. World Reference Base for Soil Resources-Its fourth edition and its history. Journal of Plant Nutrition and Soil Science 186(2):151-163. Doi: 10.1002/jpln.202200417. [ Links ]
Stahle, D. W., E. R. Cook, D. J. Burnette, J. Villanueva, … and I. M. Howard. 2016. The Mexican drought atlas: Tree-ring reconstructions of the soil moisture balance during the late pre-Hispanic, colonial, and modern eras. Quaternary Science Reviews 149:34-60. Doi: 10.1016/j.quascirev.2016.06.018. [ Links ]
Sundqvist, M. K., N. J. Sanders and D. A. Wardle. 2013. Community and ecosystem responses to elevational gradients: Processes, mechanisms, and insights for global change. Annual Review of Ecology, Evolution, and Systematics 44:261-280. Doi: 10.1146/annurev-ecolsys-110512-135750. [ Links ]
Urrutia-Jalabert, R., J. Barichivich, V. Rozas, A. Lara, … and E. Cuq. 2021. Climate response and drought resilience of Nothofagus obliqua secondary forests across a latitudinal gradient in south-central Chile. Forest Ecology and Management 485:118962. Doi: 10.1016/j.foreco.2021.118962. [ Links ]
Vicente-Serrano, S. M., S. Beguería and J. I. López-Moreno. 2010. A multiscalar drought index sensitive to global warming: the Standardized Precipitation Evapotranspiration Index. Journal of Climate 23(7):1696-1718. Doi: 10.1175/2009JCLI2909.1. [ Links ]
Vitali, A., J. J. Camarero, M. Garbarino, A. Piermattei and C. Urbinati. 2017. Deconstructing human-shaped treelines: Microsite topography and distance to seed source control Pinus nigra colonization of treeless areas in the Italian Apennines. Forest Ecology and Management 406:37-45. Doi: 10.1016/j.foreco.2017.10.004. [ Links ]
VoorTech Consoulting. 2021. The tree ring measuring program project J2X. (V5.03). Holderness, NH, United States of America. VoorTech. http://www.voortech.com/projectj2x/ . (22 de septiembre de 2023). [ Links ]
Received: September 05, 2023; Accepted: December 02, 2023
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
