










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias pecuarias
versión On-line ISSN 2448-6698versión impresa ISSN 2007-1124
Rev. mex. de cienc. pecuarias vol.11 no.4 Mérida oct./dic. 2020 Epub 02-Mar-2021
https://doi.org/10.22319/rmcp.v11i4.5295
Technical notes
QTL analysis associated to single nucleotide polymorphisms (SNP) involved in the dairy phenotype of Holstein cattle
a Universidad Autónoma de Chihuahua. Facultad de Zootecnia y Ecología. Periférico Francisco R. Almada Km 1, C.P. 31453. Cd. Chihuahua, Chih. México.
The aim was to identify QTLs associated with single nucleotide polymorphisms (SNPs) whose action contributes to the productive, reproductive and health phenotypic development of Holstein dairy cattle. 341 QTLs located in 120 genes of the Bos taurus UMD_3.1.1 genome and associated with 189 SNPs with effects on productive (FY, NM, MY, MTCAS, MBLF and PL), reproductive (CONCRATE, DPR, EMBSUR, DAYOPEN and CONCEPT) and health traits (SCC, BTBS and RESRATE) were identified. SNPs were verified in the dbSNP-NCBI database, according to which 42 % were located in introns. The Jvenn platform revealed that the SNPs rs135744058, rs110828053 and rs109503725 were common in all three traits. The network of correlations between traits and genes generated by MetScape (Cytoscape 3.4), showed a positive correlation between PL, DPR, DAYOPEN, CONCRATE and CONCEPT, and a negative correlation of FY with PL, NM, DPR and CONCRATE. The functionality of each gene was validated in the Gene-NCBI and UniProt databases, and ClueGo (Cytoscape 3.4) was used to select functional pathways with a significance value less than 0.05, which rendered an intertwining between the development of the mammary gland, the activation of the immune system and the response to steroid hormones evident, the GH gene being the one that directs this functionality. Although the genetic panorama shows that there is an antagonism between productive and reproductive traits, the functional genetic activity due to the 189 SNPs analyzed exhibits an interwoven action in ontological pathways that influence the production processes, as well as in reproductive and health pathways.
Key words QTL; SNP; Holstein
El objetivo fue identificar QTL asociados a polimorfismos de nucleótido único (SNP) cuya acción contribuya al desarrollo fenotípico productivo, reproductivo y de salud del ganado lechero de raza Holstein. Ubicados en 120 genes del genoma Bos taurus UMD_3.1.1, se identificaron 341 QTL asociados a 189 SNP con efectos sobre rasgos productivos (FY, NM, MY, MTCAS, MBLF y PL), reproductivos (CONCRATE, DPR, EMBSUR, DAYOPEN y CONCEPT) y de salud (SCC, BTBS y RESRATE). Los SNP fueron verificados en la base de datos dbSNP-NCBI donde 42 % se ubicaron en intrones. Con la plataforma Jvenn se supo que los SNP rs135744058, rs110828053 y rs109503725, fueron comunes en los tres rasgos. La red de correlaciones entre rasgos y genes generada por MetScape (Cytoscape 3.4), mostró una correlación positiva entre PL, DPR, DAYOPEN, CONCRATE y CONCEPT; y una negativa de FY hacia PL, NM, DPR y CONCRATE. La funcionalidad de cada gen fue validada en las bases Gene-NCBI y UniProt, y por ClueGo (Cytoscape 3.4) se seleccionaron vías funcionales con un valor de significancia menor a 0.05, lo que evidenció un entrelazamiento entre el desarrollo de la glándula mamaria, la activación del sistema inmune y la respuesta a hormonas esteroideas; siendo el gen GH quien dirige dicha funcionalidad. Aunque el panorama genético muestra que existe un antagonismo entre rasgos productivos y reproductivos, la actividad genética funcional debido a los 189 SNP analizados, muestra una acción entrelazada en vías ontológicas que influyen tanto en los procesos de producción, así como en vías de reproductivas y de salud.
Palabras clave QTL; SNP; Holstein
In Holstein cattle, several studies have identified quantitative trait loci (QTL) associated with productive, reproductive and health traits1-3. Thanks to the improvement of statistical methods and the development of molecular tools, it has been possible to carry out whole genome association studies (GWAS)4,5 to identify QTLs associated with a locus, whose influence on the phenotype can vary between individuals of the same species through the change of a single base in the genome; this is what is known as single nucleotide polymorphism (SNP). This type of QTL which correlates with a single SNP has an effect on the functionality of a specific gene and, therefore, an immediate action in the development of a phenotypic trait of interest6. Thus, identification of the biological pathways and genes that are associated with significant SNPs may provide a deeper biological understanding of the expression mechanism of a particular phenotypic trait7.
The objective of this study was to detect, with the aid of bioinformatics tools, those QTLs associated with a SNP with a potential effect on the phenotypic traits of production, reproduction and health of Holstein dairy cattle. 15 phenotypical characters were used for the QTL search: duration of productive life (PL), net merit (NM), milk yield (MY), milk protein yield (PY), milk fat yield (FY) , casein content (MTCAS), β-lactoglobulin content (MBLG), susceptibility to bovine tuberculosis (BTBS), respiratory rate (RESRATE), somatic cell content (SCC), conception rate (CONCRATE), daughters’ pregnancy rate (DPR), early embryo survival (EMBSUR), parturition-conception interval (DAYOPEN) and services by conception (CONCEPT). The selection was made Out of the 114,685 QTL reported in the QTLdb database (Animal QTL Database)2, those QTL associated with one of the productive and health traits and whose peak was related to a single SNP with a significance value of less than 0.05 were selected, while the QTL selected for the reproductive SNPs were those reported by Cochran et al8 that did not have a negative effect on any productive trait. Based on the Bos taurus UMD_3.1.1 genome, each SNP was verified in the dbSNP database of the NCBI (National Center for Biotechnology Information, http://www.ncbi.nlm.nih.gov/snp/), where its chromosomal location was verified, and the affected gene was classified according to its location, as: intron (if it was in an intronic region), change of direction (if it caused a change in the amino acid sequence), synonym (if the change of base did not imply a change in the amino acid sequence), promoter (if it was located in the promoter of the affected gene), deletion / insertion (if the SNP resulted in a deletion or a base insertion), or UTR3 'and UTR5' (if the SNP was located in the 5'or 3 'untranslated region of mRNA).
The function of each of the genes involved in this study was validated in both the NCBI Gene database (http://www.ncbi.nlm.nih.gov/gene/) and UniProt (The Universal Protein Resource, http://www.uniprot.org). Common SNPs among phenotypic traits were identified through a Venn diagram using the Jvenn platform9. The Pearson correlation coefficients were calculated using the MetScape algorithm10, in order to establish associations between phenotypic traits with the presence of a gene. These values were visualized as a colorimetric matrix (heat map) composed of a color spectrum that went from green to red with correlation values from -1 to 1, respectively. These data were also analyzed as a hierarchical grouping and, with them, a network of correlations was generated using the MetScape application of the Cytoscape 3.4 software11. The ontological functional network of the genes was carried out with the ClueGo application of the Cytoscape 3.4 software, under the following criteria: the ontological data of each gene was taken from the “GO Biological Process-GOA; with a "GO Tree" range from 3 to 8; the selection terms for each pathway included at least 1 gene per cluster with a kappa score of 0.3; in addition, it was tested with a bilateral hypergeometric statistical test with Bonferroni correction, only those pathways with a significance value less than 0.05 were taken into account.
The results published to date on GWAS in dairy cattle have provided information on the influence of SNP on the expression of a QTL4,5,12. In the genome, SNP are the most abundant forms of DNA and due to their low mutation rate; they are excellent selection markers13,14, in addition to being easy and inexpensive to perform genotyping15. Taking the information included in the QTLdb database2, 341 QTLs associated with a SNP were found, of which 70 % were production QTL, 17 % were involved in health and 13 % in reproductive traits (Figure 1). Within the production phenotype, the most favored trait was PY, followed by MY, then FY, NM, PL, MBLG and lastly MTCAS. In health characters, it was BTBS followed by SCC and RESRATE. In reproductive traits, more QTL were associated with CONCRATE, followed by DPR, CONCEPT, and EMBSUR, which was the least favored. There were QTL that had the same SNP, as well as SNPs that were present in different regions of the same gene. Therefore, in the end, 341 QTL associated with 189 SNP located in 120 genes of the Bos taurus genome were identified. Each of the 189 SNP were verified in the NCBI dbSNP database, along with their location and the gene that they affected.
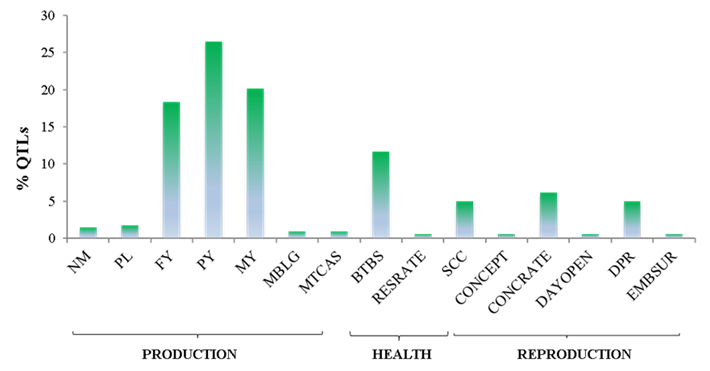
Figure 1 Distribution of 341 QTLs associated with a single SNP, according to the phenotype they favor
According to the information collected, 42 % of the SNP were intronic; 27 % SNP were located in coding regions, of which 17 % had an effect on the amino acid change of the protein and 5 % were synonymous mutations; 6 % were localized at the untranslated 3 'end (UTR3'), and 1 %, in UTR5 '. 6.7 % of the SNPs were located within the SAA2 gene, 4.1 % were located in the ATF3 gene, 3.8 % in the HSD17B7 gene, and 3.0 % were located in each of the AP3B1 and CARD15 genes. The BCHE and PDE9A genes had 2.6 %, while the PCC8, CDKN1A and CSN2 genes had 2 % each. 1.2 to 1.8 % of the SNP were located in the regions of the APP, GNAS, NRPL48, SERPINA5, SLC8A1, CACNA1D, COQ9, DGAT1, DSC2, FASN, IGF1R, LEP, PRLR, CSNK1E, BRINP3 and HSD17B13 genes (Figure two). With the exception of the Bta-9 and Bta-12 chromosomes, the 189 SNP were distributed in all autosomes and on the X chromosome of the Bos taurus UMD 3.1.1 genome (Figure 3). However, the distribution was uneven; there were chromosomes that exhibited more SNP than others, and in the same way, there were chromosomes that included few SNP, but these were associated with many QTL.


The QTLs most often identified were mainly productive, which supports the fact that the quantity and quality of milk produced go hand in hand with economic gains16. In Bta-14, the SNP rs109421300 located in the DGAT1 gene has more influence on the MY, FY and PY traits14,15,17-19. Bta-5 associates with MY, while FY and PY associate with the SNP rs41591907, rs41256890, rs41592943, rs41592948, rs137408198, and rs1334491667,17,20, located in the BCATI, MGP, GUCY2C, CDEABAR1, and CDEABAR1 genes. Bta-18 has been associated with both MY and PY, due to the influence of the SNP rs4158169414,17, which is located in the FOXA3 gene. In Bta-20, the SNP rs385640152 causes a change in the GHR protein that affects MY and PY (7). In Bta-21, the SNP rs41644615 located in the SERPINE5 gene is associated with MY and PY19. Finally, the SNP rs110475419 is associated with PY14, which is in the ADAM12 gene in Bta-26.
Milk is made up of the proteins α-lactalbumin, β-lactoglobulin, and the α, β and κ caseins21. Bta-11 has been associated with β-lactoglobulin (MBLG) concentrations in milk, due to the effect of the PAEP gene, which has the SNP rs4125567916,21,22, rs11006622921, and rs11018046321. On the other hand, Bta-6 has been related to MTCAS through the SNP rs10929940123, which, when expressed in the CSN2 gene, causes a functional change in β-casein. MTCAS has also been associated with Bta-14 and Bta-19 due to the SNPs rs110757796 and rs41923484, respectively23. In dairy cattle, the most common disease and the one that causes the most economic losses is mastitis, a disease with which the somatic cell count (SCC) in milk is associated as a predictor12. The chromosomes that have the greatest influence on SCC are Bta-6, Bta-13, Bta-14, Bta-19 and Bta-2024. There are 6 SNPs in Bta-6, of which rs43703013, rs43703011and rs109299401 are in the coding region of the CSN2 gene, where each one of them causes an amino acid change in the β-casein protein, while rs110239379 and rs110118210 are located in the same intron of the KIT gene, and rs109757609 is located in the promoter of the CSNIS1 gene. The association with SCC in Bta-13 was related to two SNPs in the SRC gene, rs41703851 and rs41602996. Bta-14 contributed to SCC due to SNPs rs109162116 and rs109234250 in the DGAT1 gene. Other SNPs associated with SCC are rs109149276 and rs109149276, of the FASN gene of Bta-19 and of the promoter of the PRLR gene in Bta-20, respectively24, as well as rs43315150, located in Bta-2, in an intron of the CYP27A1 gene20.
Tuberculosis is another disease that causes economic losses in the livestock industry; bovines are not only one of the animal species that most coexists with humans but have also become one of the main sources of spread of this disease worldwide25. Richardson et al1 found BTBS associated to Bta-1 with 9 SNP (rs42294486, rs29020933, rs29020933, rs42294431, rs42294441, rs110098599, rs132953892, rs41665131 and rs43741780 gene, located in the BCHE region), and with 2 SNPs (rs109186526 and rs110679397) located in the same intron of the KALRN gene. In the chromosomes Bta-3, Bta-8, Bta-10 and Bta-23, the same research identified the SNP rs110622046, rs135916795, rs109277058 and rs41642913, located in intronic regions of the MAEL, PTPRD, AP3B1 and FKBP5 genes, respectively. On the other hand, Finlay et al26 reported that in Bta-22 the SNP rs42286978, rs42287005 and rs42724727, which are located in the promoter region of the SLC6A6 gene, are also associated with susceptibility to tuberculosis. Wang et al27 reported that in Bta-18 the CARD15 gene had 3 SNPs within the coding region of exon 4, as well as one SNP located in the intron prior to exon 4.
As for reproductive traits, these were located very close to productive traits in chromosomes Bta-1, Bta-3, Bta-5, Bta-10, Bta-15, Bta-16, Bta-18, Bta-22, Bta-23 and Bta-24 (Figure 3). Notably, although these SNPs represent only a small portion of the genes involved in the reproductive process, they were selected from the works of Cochran et al8 and Ortega et al28, who proved that these SNP have a positive effect on the reproductive traits associated with fertility and, at the same time, they exert no negative influence on productive traits. Of the total SNPs only rs135744058, rs110828053 and rs109503725 were common to productive, health and reproductive traits alike. The SNP rs135744058 is associated with the MY8, CONCRATE8,28, DPR8, and RESRATE characters29. In the genetic language, it is composed of the A/G variants 8 and is located in the exonic region of the CACNAID gene where it brings about the change in the α1D subunit of the calcium voltage channel30. In cattle, its presence has been detected at the level of the hypothalamus during the development of the central nervous system31. The SNP rs110828053 has been reported to be associated with NM8, DPR28, CONCRATE28, CONCEPT28 and RESTATE29 and it generates an A/G swap in the HSD17B7 gene that causes a change that substitutes alanine for threonine at position 308 of the protein. The HSD17B7 gene encodes for dehydrogenase 7 hydrosteroid 17-β, which participates in the biosynthesis of sex steroids and cholesterol28; in bovines, it has been located in the oviduct epithelial cells32, being essential for ovarian function and regulation of fertility33. The SNP rs109503725, composed of the variables T/C in the DSC2 gene, is associated with the PL8, DPR8,28, CONCRATE28 and SCC8 characters. This SNP generates a change in amino acid 535, substituting arginine for tryptophan in the protein Desmocholine 2, which is a protein involved in cell-to-cell junctions, forming desmosomes in epithelial cells34.
During the last two decades in dairy cattle, selection strategies have focused on developing highly dairy producing animals, which has resulted in a decline in reproductive capacity. This has generated a negative genetic association, i.e. an antagonist, between the productive and reproductive traits20,35-37. Now, in this study, reproductive SNP were previously reported as non-antagonists with productive traits8,28; in fact, these SNP are located in the same regions as SNPs associated with productive traits (Figure 3). In order to locate those traits that are positively related, a correlation analysis of phenotypical characters was performed, based on the genes that they shared (Figure 4A). This analysis managed to classify the characters in two groups, grouped into two clades. Clade 1 comprised all the reproductive traits together with the PL and NM characters of the productive traits. The rest of the productive traits (MY, PY, FY, MTCAS and MBLG) were grouped together with the health traits in clade 2 >(Figure 4B).
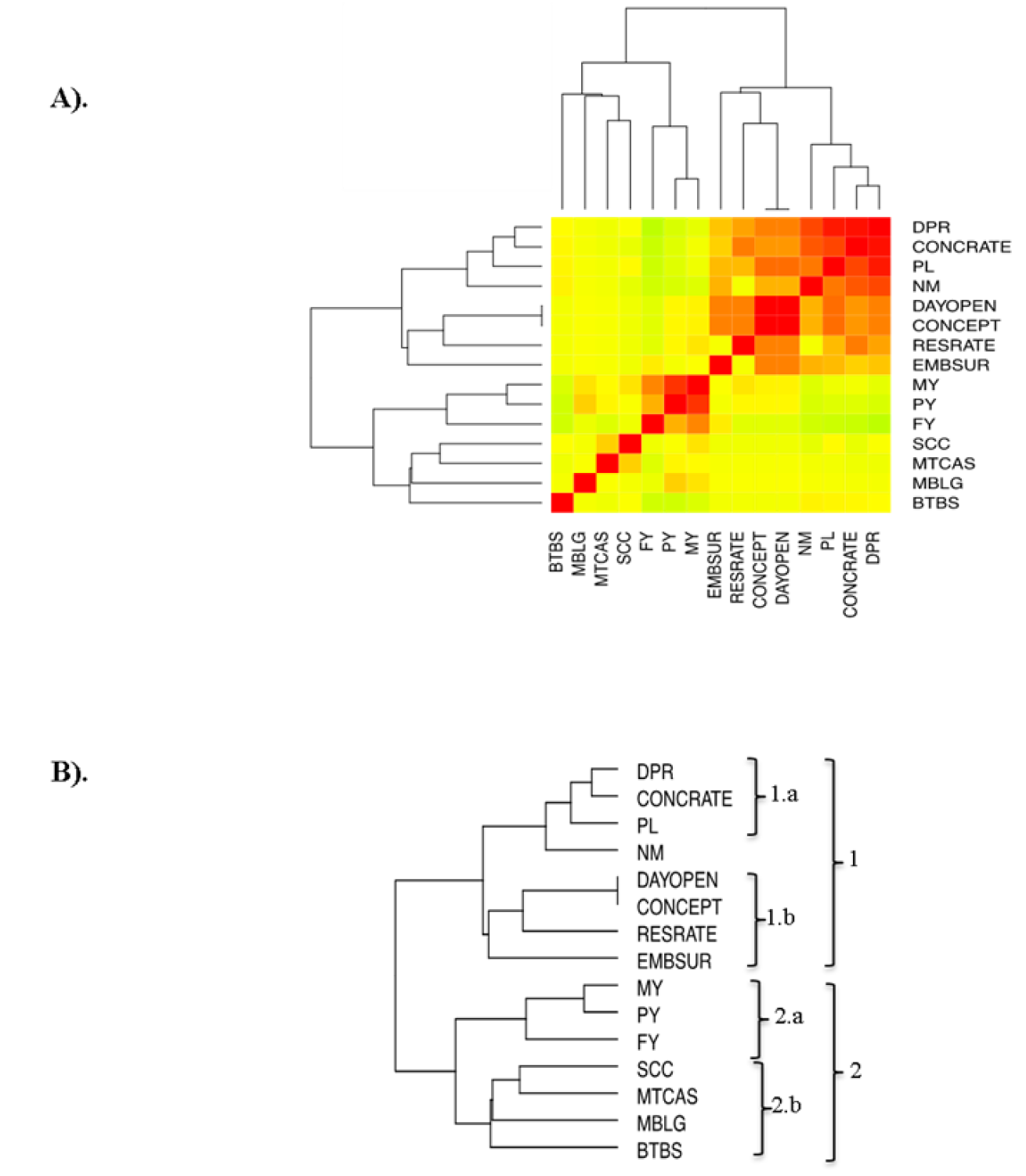
(A) Pearson's correlation matrix (Heat map), the color spectrum goes from green to red, with values from -1 to 1, respectively. (B) Hierarchica grouping of A. DPR daughters’ = pregnancy rates; CONCRATE = conception rate; PL = duration of reproductive life; NM = Net merit; DAYOPEN = birth-conception interval; CONCEPT = conception inseminations; RESRATE = respiratory rate; EMSUR = early embryo survival; MY = milk yield; PY = protein yield in milk; FY = fat yield in milk; SCC = somatic cell count in milk; MTCAS = casein content in milk; MBLG = content of β-lactoglobuin in milk; BTBS = susceptibility to bovine tuberculosis.
Figure 4 Correlation of phenotypic traits based on a SNP associated with a QTL
Likewise, when analyzing the correlation network of genes based on the trait, two groups of genetic networks could be observed (Figure 5): Group A, corresponding to those genes present in the characters FY, MY and PY (productive traits), and Group B, with genes present in BTBS, SCC, CONCEPT, CONCRATE, DAYOPEN, DPR, EMBSUR, NM, PL, RESRATE, encompassing the health, reproduction and production phenotypes. These results indicate that at the genetic level there is an antagonistic scenario between the production and reproduction QTL.
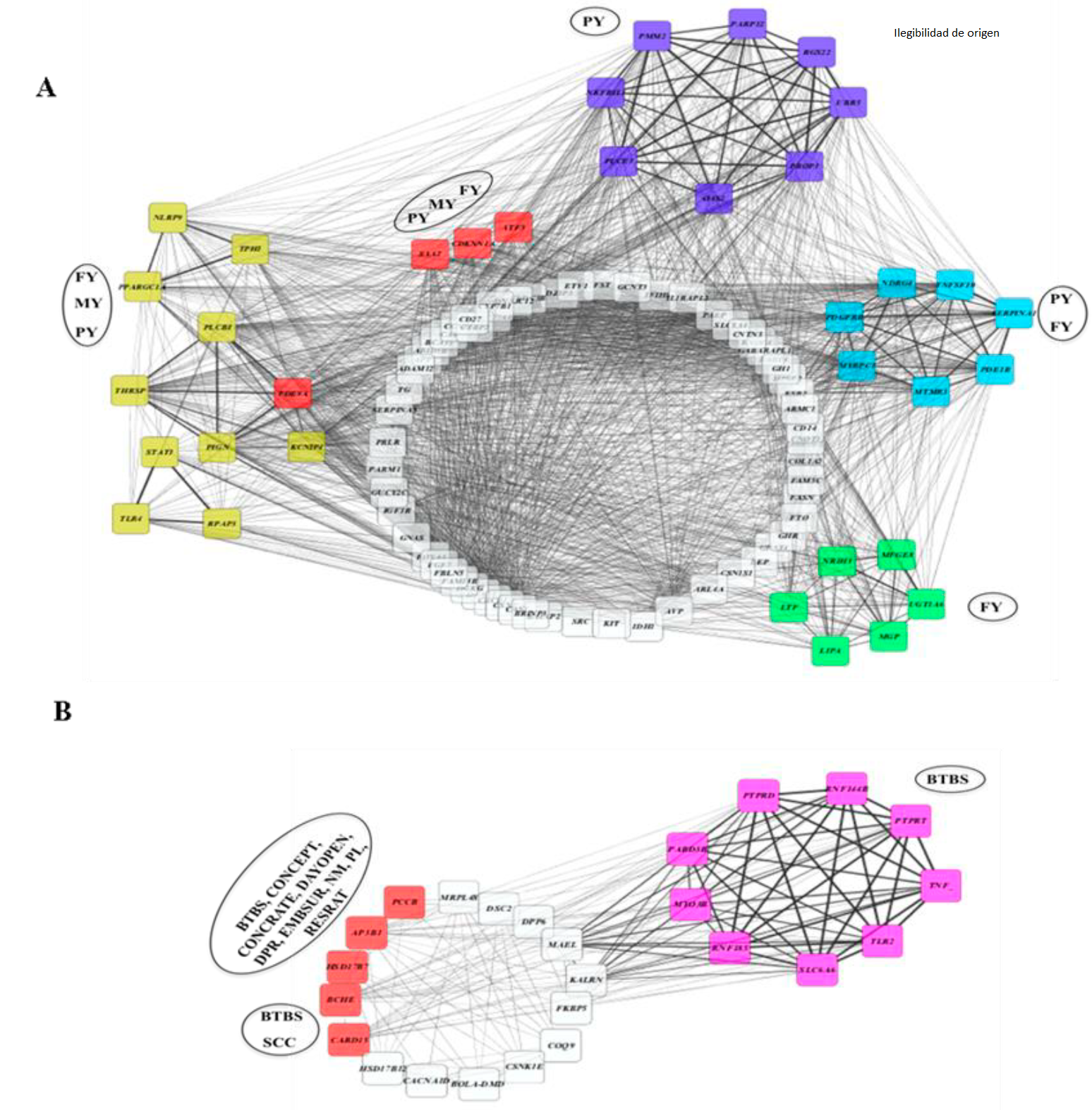
The nodes represent the gene, and the black lines represent the correlations between the nodes. The analysis was carried out with the MetScape application of the Cytoscape software, considering the positive ones from 0.5 to +1. The thicker the line, the higher the correlation. The nodes in yellow, purple, blue, green and pink; represent nodes with high correlation generating groups, and the nodes in red represent the genes with the most SNP involved in the panel. The legends within the black ovals represent the phenotypical character associated with the genes in each group of nodes.
Figure 5 Pearson's correlation network of genes associated with a phenotypic trait
In order to encompass the functional information of the genes involved in this study, an ontological network was designed in which those genes whose functions were related to each other were located. (Figure 6) shows that, starting from the development of the mammary gland where the genes CSN2, FASN, GH, GPAT4, SRC and TPH1 participate, the GH gene is linked to the pathways that correspond to lipid development, where the DGAT1, GPAT4 and THRSP genes are found. Hence, GH and GPAT4 are linked to cell growth pathways such as the development of adipose tissue, in which the APP, ARID5B, FTO, GNAS, GPAT4, LEP, PPARGC1A genes participate; of these, ARID5B is the one that leads to the differentiation of fat cells, while FTO, LEP and PPARBC1A participate in the differentiation into brown fat cells. Likewise, GH, LEP and PPARGC1A are linked, together with CSN1S1, with the steroid hormone response pathways and are related to the regulation of the immune system.
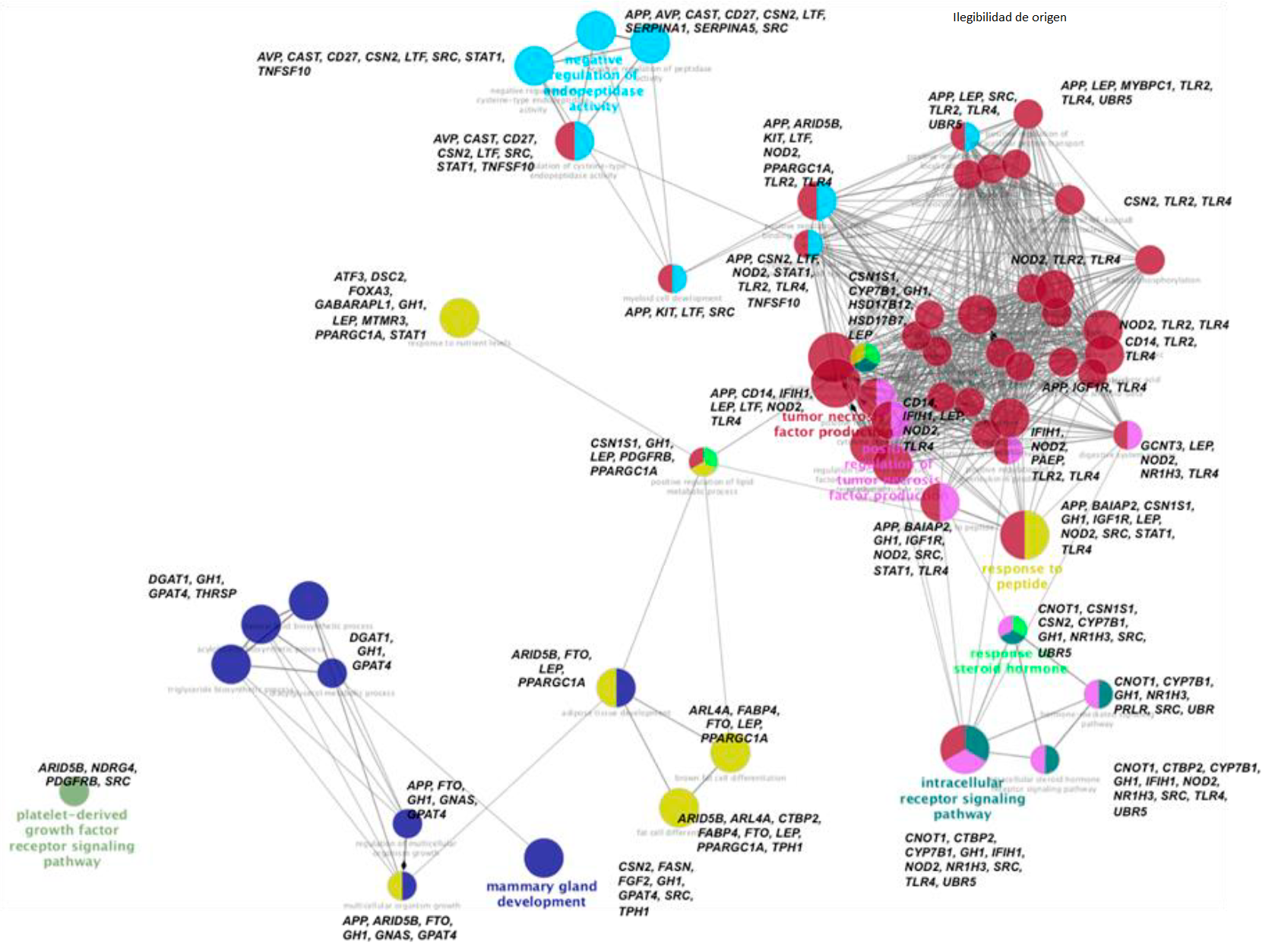
Each GO is represented as a circular node, whose size is related to the significance (P <0.05) and the number of genes associated with a particular biological process; they are grouped by color, each color representing one of these processes. The genes involved in each GO are indicated in bold letters.
Figure 6 Genetic ontology (GO) of the genes that make up the Panel, depicted within a network of biological functions generated with the ClueGo application of the Cytoscape 3.4 software
The genes involved in the response to steroid hormones are CSN1S1, CYPP7B1, GH, HSD17B12, HSD17B7, and LEP; thus, again, GH and CSN1S1 lead to the hormone signaling pathways in which CNOT1, NRH13, SRC, UBR, CTBP2, CYPB7B1, NOD2 and UBR5 participate. GH is also linked to the regulation of the immune system, specifically to the production of the tumor necrosis factor involving the genes APP, BAIAP2, GH, IGF1R, NOD2, SRC, STAT1 and TLR4. Hence, NO2 and TLR4 are the bridge to other pathways of the immune system, such as the activation of cytosines and interleukins and that of the innate immune system. Interestingly, at this point we find LEP once more, which leads back to the lipid metabolic process. When it is looked at this functional ontological pathway from a global perspective, it seems that GH acts as a master gene that directs, like an orchestra conductor, the functional activity of the other genes involved in the development of dairy cattle. The GH gene is located in Bta-19 and codes for the protein somatotropin. This gene is associated with PY and MTCAS due to the effect of the SNP rs41923484, which causes a change in amino acid 153 by substituting valine for leucine in somatotropin23. Somatotropin, also known as growth hormone, participates in multiple activities, which range from its effect on cell growth to the differentiation of various tissues, such as the development of follicles38 and of the mammary gland39; therefore, it is not surprising that its action is crucial for the entire body of Holstein cattle to function properly. The functional activity of the 120 genes associated with 189 SNPs within 341 QTL is intertwined within the ontological pathways that influence both the production processes, such as the development of the udders, and the reproductive and health pathways. Awareness of this fact opens the door to improving selection in order to generate animals that will gradually acquire both productive and reproductive traits.
Acknowledgements
The authors thank CONACYT for the support granted through project No. 216179
REFERENCES
1. Richardson IW, Berry DP, Wiencko HL, Higgins IM, More SJ, McClure J, et al. A genome-wide association study for genetic susceptibility to Mycobacterium bovis infection in dairy cattle identifies a susceptibility QTL on chromosome 23. Genet Sel Evol 2016;48:19-31. [ Links ]
2. Hu ZL, Park CA, Reecy JM. Developmental progress and current status of the Animal QTLdb. Nucleic Acids Res 2016;44(D1):D827-D833. [ Links ]
3. Yudin NS, Voevoda MI. Molecular genetic markers of economically important traits in dairy cattle. Russian J Genetics 2015;51(5):506-517. [ Links ]
4. Abo-Ismail MK, Brito LF, Miller SP, Sargolzaei M, Grossi DA, Moore SS, et al. Genome-wide association studies and genomic prediction of breeding values for calving performance and body conformation traits in Holstein cattle. Genet Sel Evol 2017;49(1):82-110. [ Links ]
5. Pegolo S, Mach N, Ramayo-Caldas Y, Schiavon S, Bittante G, Cecchinato A. Integration of GWAS, pathway and network analyses reveals novel mechanistic insights into the synthesis of milk proteins in dairy cows. Sci Rep 2018;8(1):566-580. [ Links ]
6. Wang M, Hancock TP, Chamberlain AJ, Vander Jagt CJ, Pryce JE, Cocks BG, et al. Putative bovine topological association domains and CTCF binding motifs can reduce the search space for causative regulatory variants of complex traits. BMC Genomics 2018;19(1):395-411. [ Links ]
7. Nayeri S, Sargolzaei M, Abo-Ismail MK, May N, Miller SP, Schenkel F, et al. Genome-wide association for milk production and female fertility traits in Canadian dairy Holstein cattle. BMC Genet 2016;17(1):75-85. [ Links ]
8. Cochran SD, Cole JB, Null DJ, Hansen PJ. Discovery of single nucleotide polymorphisms in candidate genes associated with fertility and production traits in Holstein cattle. BMC genetics 2013;14(1):49-71. [ Links ]
9. Bardou P, Mariette J, Escudié F, Djemiel C, Klopp C. jvenn: an interactive Venn diagram viewer. BMC bioinformatics 2014;15(1):293-299. [ Links ]
10. Basu S, Duren W, Evans CR, Burant CF, Michailidis G, Karnovsky A. Sparse network modeling and metscape-based visualization methods for the analysis of large-scale metabolomics data. Bioinformatics 2017;33(10):1545-1553. [ Links ]
11. Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome research 2003;13(11):2498-2504. [ Links ]
12. Duran Aguilar M, Roman Ponce SI, Ruiz Lopez FJ, Gonzalez Padilla E, Vasquez Pelaez CG, Bagnato A, et al. Genome-wide association study for milk somatic cell score in Holstein cattle using copy number variation as markers. J Anim Breed Genet 2017;134(1):49-59. [ Links ]
13. Karim L, Takeda H, Lin L, Druet T, Arias JA, Baurain D, et al. Variants modulating the expression of a chromosome domain encompassing PLAG1 influence bovine stature. Nat Genet 2011;43(5):405-413. [ Links ]
14. Kolbehdari D, Wang Z, Grant JR, Murdoch B, Prasad A, Xiu Z, et al. A whole genome scan to map QTL for milk production traits and somatic cell score in Canadian Holstein bulls. J Anim Breed Genet 2009;126(3):216-227. [ Links ]
15. Fang M, Fu W, Jiang D, Zhang Q, Sun D, Ding X, et al. A multiple-SNP approach for genome-wide association study of milk production traits in Chinese Holstein cattle. PLoS One 2014;9(8):e99544-e99551. [ Links ]
16. Gambra R, Penagaricano F, Kropp J, Khateeb K, Weigel KA, Lucey J, et al. Genomic architecture of bovine kappa-casein and beta-lactoglobulin. J Dairy Sci 2013;96(8):5333-5343. [ Links ]
17. Gervais O, Pong-Wong R, Navarro P, Haley CS, Nagamine Y. Antagonistic genetic correlations for milking traits within the genome of dairy cattle. PLoS One 2017;12(4):e0175105-e0175117. [ Links ]
18. Cole JB, Wiggans GR, Ma L, Sonstegard TS, Lawlor TJ, Crooker BA, et al. Genome-wide association analysis of thirty one production, health, reproduction and body conformation traits in contemporary US Holstein cows. BMC genomics 2011;12(1):408. [ Links ]
19. Pesek P, Pribyl J, Vostry L. Genetic variances of SNP loci for milk yield in dairy cattle. J Appl Genet 2015;56(3):339-347. [ Links ]
20. Pimentel EC, Bauersachs S, Tietze M, Simianer H, Tetens J, Thaller G, et al. Exploration of relationships between production and fertility traits in dairy cattle via association studies of SNPs within candidate genes derived by expression profiling. Anim Genet 2011;42(3):251-262. [ Links ]
21. Huang W, Penagaricano F, Ahmad KR, Lucey JA, Weigel KA, Khatib H. Association between milk protein gene variants and protein composition traits in dairy cattle. J Dairy Sci 2012;95(1):440-449. [ Links ]
22. Schopen GC, Visker MH, Koks PD, Mullaart E, van Arendonk JA, Bovenhuis H. Whole-genome association study for milk protein composition in dairy cattle. J Dairy Sci 2011;94(6):3148-158. [ Links ]
23. Viale E, Tiezzi F, Maretto F, De Marchi M, Penasa M, Cassandro M. Association of candidate gene polymorphisms with milk technological traits, yield, composition, and somatic cell score in Italian Holstein-Friesian sires. J Dairy Sci 2017;100(9):7271-7281. [ Links ]
24. Fontanesi L, Calo DG, Galimberti G, Negrini R, Marino R, Nardone A, et al. A candidate gene association study for nine economically important traits in Italian Holstein cattle. Anim Genet 2014;45(4):576-580. [ Links ]
25. Wang W, Cheng L, Yi J, Gan J, Tang H, Fu MZ, et al. Health and production traits in bovine are associated with single nucleotide polymorphisms in the NOD2 gene. Genet Mol Res 2015;14(2):3570-3578. [ Links ]
26. Finlay EK, Berry DP, Wickham B, Gormley EP, Bradley DG. A genome wide association scan of bovine tuberculosis susceptibility in Holstein-Friesian dairy cattle. PLoS One 2012;7(2):e30545-e135094. [ Links ]
27. Wang Y, Wang S, Liu T, Tu W, Li W, Dong G, et al. CARD15 Gene Polymorphisms Are Associated with Tuberculosis Susceptibility in Chinese Holstein Cows. PLoS One 2015;10(8):e0135085. [ Links ]
28. Ortega MS, Denicol AC, Cole JB, Null DJ, Hansen PJ. Use of single nucleotide polymorphisms in candidate genes associated with daughter pregnancy rate for prediction of genetic merit for reproduction in Holstein cows. Anim Genetics 2016;47(3):288-297. [ Links ]
29. Dikmen S, Wang XZ, Ortega MS, Cole JB, Null DJ, Hansen PJ. Single nucleotide polymorphisms associated with thermoregulation in lactating dairy cows exposed to heat stress. J Anim Breed Genet 2015;132(6):409-419. [ Links ]
30. Srivastava U, Aromolaran AS, Fabris F, Lazaro D, Kassotis J, Qu Y, et al. Novel function of alpha1D L-type calcium channel in the atria. Biochem Biophys Res Commun 2017;482(4):771-776. [ Links ]
31. Peruffo A, Giacomello M, Montelli S, Panin M, Cozzi B. Expression profile of the pore-forming subunits alpha1A and alpha1D in the foetal bovine hypothalamus: a mammal with a long gestation. Neurosci Lett 2013;556:124-128. [ Links ]
32. Cerny KL, Garrett E, Walton AJ, Anderson LH, Bridges PJ. A transcriptomal analysis of bovine oviductal epithelial cells collected during the follicular phase versus the luteal phase of the estrous cycle. Reprod Biol Endocrinol 2015;13:84-96. [ Links ]
33. Kemilainen H, Adam M, Maki-Jouppila J, Damdimopoulou P, Damdimopoulos AE, Kere J, et al. The hydroxysteroid (17beta) dehydrogenase family gene HSD17B12 is involved in the prostaglandin synthesis pathway, the ovarian function, and regulation of fertility. Endocrinology 2016;157(10):3719-3730. [ Links ]
34. Gehmlich K, Lambiase PD, Asimaki A, Ciaccio EJ, Ehler E, Syrris P, et al. A novel desmocollin-2 mutation reveals insights into the molecular link between desmosomes and gap junctions. Heart Rhythm 2011;8(5):711-718. [ Links ]
35. Pryce J, Royal M, Garnsworthy P, Mao IL. Fertility in the high-producing dairy cow. Livestock Prod Sci 2004;86(1-3):125-135. [ Links ]
36. Pryce J, Veerkamp R. The incorporation of fertility indices in genetic improvement programmes. BSAP Occasional Publication 2001;26(1):237-249. [ Links ]
37. Peñagaricano F, Khatib H. Association of milk protein genes with fertilization rate and early embryonic development in Holstein dairy cattle. J Dairy Res 2011;79(01):47-52. [ Links ]
38. Cushman R, DeSouza J, Hedgpeth V, Britt J. Effect of long-term treatment with recombinant bovine somatotropin and estradiol on hormone concentrations and ovulatory response of superovulated cattle. Theriogenology 2001;55(7):1533-1547. [ Links ]
39. Akers RM. A 100-Year Review: Mammary development and lactation. J Dairy Sci 2017;100(12):10332-10352. [ Links ]
Received: March 19, 2019; Accepted: September 17, 2019
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
