











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink1. Introduction
Agriculture has been the main source of NH3 emissions in Europe for a long time (Asman, 1998). Several studies have reported increasing atmospheric concentration of NH3 and ammonium, especially in the regions of concentrated animal feeding operations (Theobald et al., 2006; Walker et al., 2008; Sutton et al., 2011; Jones et al., 2013). Emissions of atmospheric NH3 from agriculture and its subsequent entry into sensitive ecosystems is an important environmental problem (Bobbink et al., 2002, 2010, 2011). NH3 deposition may cause soil acidification through nitrification processes (van Breemen et al., 1982). More importantly, NH3 plays a significant role in eutrophication of sensitive, mainly terrestrial ecosystems (Sutton et al., 2009; Theobald et al., 2009; Paoli et al., 2010). Atmospheric inputs of NH3 may cause a decrease of biodiversity in sensitive ecosystems (Stevens et al., 2004; Emmett, 2007; Jones et al., 2011). NH3 significantly affects the Earth´s climate, mainly because the volatilization potential of NH3 nearly doubles for every 5 ºC increase in temperature (Sutton et al., 2013). Measurement of NH3 emissions from naturally ventilated animal houses is technically challenging (Phillips et al., 2001; Scholtens et al., 2004; Welch et al., 2005a, b) due to the difficulties in determining the ventilation rate. Ventilation rate depends on the building design, animal occupancy, temperature, wind speed, wind direction and accuracy of the NH3 measurement (Phillips et al., 2001; Welch et al., 2005b; Zhang et al., 2005). However, methods such as the internal or external tracer-ratio techniques or the flux-sampling technique have been successfully tested under real conditions (Demmers et al., 1999; Dore et al., 2004). The conservation of mass is an important concept that underlies environmental analysis of buildings (Albright, 1990). The concept of mass conservation is applied to latent heat (humidity) and gaseous contaminants (Samer et al., 2012). Several studies have investigated moisture balance and tracer gas technique for ventilation rates measurements and NH3 emissions quantification in naturally ventilated buildings (Samer et al., 2011). Pedersen et al. (1998), Teye and Hautala (2007) and Samer et al. (2012) investigated the H2O balance for ventilation rate measurements. The H2O mass balance method largely depends on animal production of H2O. Factors that influence H2O production include flooring system, stocking, density, watering, moisture content of the forages, animal activity and relative humidity.
In the vicinity of intense agricultural activities, deposition of atmospheric NH3 may totally dominate the overall load of reactive nitrogen (N) from the atmosphere (Hertel et al., 2006), and NH3 dry deposition velocities in the areas of intensive animal production may exceed critical loads (Kuylenstierna et al., 1998).
NH3 dry deposition velocities for grassland, crops and forest are different (Smith et al., 2000; Theobald et al., 2006). Several field studies have shown that the deposition velocity of NH3 for forests is relatively high and variable (Wyers et al., 1992; Duyzer et al., 1994; Andersen et al., 1999). Due to its high deposition velocity and its reactivity in the atmosphere, gaseous NH3 has a relatively short atmospheric lifetime (few days or less; Warneck, 1999). Dry deposition fluxes of NH3 to grassland, crops and forest are different (Zapletal 1998, 2001; Phillips et al., 2004; Walker et al., 2006). The difference in fluxes is caused by the differences in emission strenght, and by differences between vegetation of low and high N status (Sutton et al., 1997; Flechard and Fowler, 1998; Milford et al., 2001).
The objective of this study was to compare the inferred emission rates in a naturally ventilated dairy farm during a three months period with measured concentrations of NH3 to obtain data for derivation of the emission-deposition relationship around a dairy farm.
2. Materials and methods
2.1 Site and building description
The dairy barn was located in a rural area in the east of the Czech Republic at an altitude of 300-350 masl. About 240 dairy cows in loose housing were the main source of NH3 emissions from the barn (Fig. 1), which was 88 m long and 27 m wide. The height of the sheet metal roof was 9 m at the gable peak. The internal room volume of the barn was 21 100 m3 (87.9 m3 per animal). The dairy barn was naturally ventilated by a draft introduced into the building through adjustable curtains on the long sidewalls (protected by nets), which were open in summer, and through five doors with a size of 3 × 4 m in the gable wall of the northern and southern sides. The prevailing summer winds are north and south winds. Ambient meteorological parameters were measured from May to July at a height of 2 m above ground and 100 m from the dairy barn (Fig. 2, site B). Temperature and humidity were measured by a Young 41372LC/LF sensor (Young Company,Traverse City, Michigan, USA). Wind speed and direction were measured by a Gill 2D ultrasonic anemometer (Hampshire, UK), intensity of solar radiation was measured by an RS 81 pyranometer (Envitech Bohemia, Prague, Czech Republic). Data was stored in an UJED datalogger (Baghirra, Prague, Czech Republic) at 1-min intervals. The distribution of sensors is displayed in Fig. 1.

2.2 Estimation of air exchange rates
Moisture from animal respiration, evaporation from manure, and forages can be used as a natural gas tracers. The ventilation rate throughout the building can be determined by the mass balance of H2O. The moisture balance was based on several studies (Pedersen et al., 1998; Teye and Hautala, 2007; Samer et al., 2012). The following mathematical model describes the ventilation rate:
where Q H2O (m3 s-1) represents the ventilation rate subject to the H2O-balance; v (m3 kg-1 dry air) is the specific volume; W i (g H2O kg-1 dry air) is the humidity ratio inside the building, and W o (g H2O kg-1 dry air) is the humidity ratio outside the building. Humidity ratios were determined by the psychometrics software EZAir Properties v.1.3.5 (R.M. Parks, Gradyville, PA, USA). M w (g H2O s-1) represents the moisture produced by the cows housed in the building, and can be calculated as follows:
where n represents the number of cows housed in the building, M w (g H2O s-1) is the moisture produced by one dairy cow, M avg (kg) is the average mass of the cows, and P H2O (g H2O h-1 kg-1) is the moisture produced by a dairy cow per mass unit, which is equal to 1.8 (Lindley and Whitaker, 1996).
2.3 Estimation of NH3 emissions
NH3 concentrations, air temperature and relative humidity were measured inside and outside the barn (Fig. 1) according to Samer et al. (2012). Multiwarn II gas detectors with infrared sensors (Dräger Safety, Germany) were used to measure NH3 concentrations at eight points (G2, G3, G5, G6, G7, G8, G10 and G11) inside the barn and four points (G1, G4, G9 and G12) outside the barn at a height of 2.8 m during one week during the summer (July) (Fig. 1). The outdoor gas sampling point (G4) and temperature-humidity sensor (TH3) were placed 30 m from the eastern side of the building. The outdoor gas sampling point (G9) and temperature-humidity sensor (TH4) were placed 30 m from the western side of the building. The outdoor gas sampling points (G12) and G(1) were placed 30 m from the northern and southern sides of the building.
The emission was calculated according to the following equation (Zhang et al., 2005; Wang et al., 2006):
where E g;animal is the gas emission per animal in g s-1, V is the ventilation rate in m3 s-1, ρ g is the gas density in g m-3, and N a is the total number of animals. Indoor gaseous concentrations (µg m-3) G c;in were calculated as the average of values measured by the eight inside gas sampling points. The outdoor gaseous concentrations G c;out were calculated as the average of values measured by the four outside gas sampling points.
2.4 Ambient NH3 concentrations measurements
Monthly integrated NH3 concentrations were measured using a passive diffusion-tube sampler (Gradko International) described by Sutton et al. (2001), which was composed of polyacrylate tube approximately 4 cm long, with an internal diameter of about 1 cm and ends fitted with a polyethylene cover. Replacement of the sampler was performed every four weeks. Before and after exposure, the samplers were placed in a cool and dark place. The concentration of NH3 was measured (Fig. 2) at 12 locations (A to L), positioned as follows: locations A to E, 5 to 400 m to the north; locations F to G, 50 to 150 m to the south; locations H to I, 300 to 400 m to the west, and locations J to L, 150 to 400 m to the east. Passive samplers were situated at a height of 1.4 m.

Fig. 2 Schematic view of the study area, including the barn and sampling points for gas concentrations along upwind transects.
NH3 concentrations were continuously measured in location B, 100 m north of the dairy barn using on-line instrumentation during five days in July, when the emission rate of this gas was measured in parallel. NH3 was detected using a well known fluorimetric method (Genfa, 1989) and continuously collected in a cylindrical wet effluent diffusion denuder (Mikuška et al., 2008). The limit of detection of NH3 was 0.102 ppb (Kapoun, 2007) and the calibration curve was linear over 3 orders of magnitude (0.102-102 ppb NH3).
2.5 Modelling of dry NH3 deposition
Dry deposition fluxes were estimated from NH3 concentrations in air multiplied by the corresponding deposition velocities:
where F is the deposition flux of the NH3 to a unit area, V d (cm s-1) the deposition velocity of the NH3, and C(z) the concentration of NH3 at a height z above surface. Deposition velocity for NH3 (V d ) was calculated using the resistance analogy:
where R a (s cm-1) is the aerodynamic resistance for the turbulent layer, R b (s cm-1) is the laminar layer resistance for the quasi-laminar layer, and R c (s cm-1) is the surface or canopy resistance of the receptor. R a was calculated from micrometeorological relations as suggested by Voldner et al. (1986) and Hicks et al. (1987). R b was calculated from micrometeorological relations as suggested by Hicks et al. (1987). The surface roughness z0 = 90 cm was chosen for deciduous and coniferous forest, the surface roughness z0 = 2.7 cm was chosen for crop (Voldner et al., 1986).
Surface resistance was calculated using the following equation (Erisman and Draaijers, 1995):
R c was expressed based on the known global radiation, surface temperature, relative humidity, and land cover according to Eq. (7), using the results and assumptions obtained from literature for computing and parameterization of the canopy stomatal resistence, R sto (Wesely, 1989; Baldocchi et al., 1987); the mesophyll resistence, R m (Wesely, 1989; Erisman and Draaijers, 1995); the canopy cuticle or external leaf resistance, R ext (Baldocchi et al., 1987); the soil resistance, R soil (Meyers and Baldocchi, 1988), and incanopy resistance, R inc (van Pul and Jacobs, 1993). The stomatal resistance (R sto ) includes dependence upon global radiation and surface air temperature. Here we use the following generalized function to estimate the canopy stomatal resistance (Wesely, 1989):
where R i is the input resistance (s m-1), G is the global radiation (W m-2), and T s is the surface air temperature (ºC). The appropriate value of R i was chosen from Wesely (1989). The incanopy resistance (R inc ) for vegetation was modelled according to van Pul and Jacobs (1993):
where LAI is the leaf area index (m2 m-2), h the vegetation height (m), b an empirical constant taken as 14 m-1, and u * the friction velocity (cm s-1).
3. Results and discussion
3.1 Estimation of air exchange rate and NH3 emission
Table I shows the air exchange rates in the dairy barn estimated by the H2O-balance method, ventilation rate, relative humidity and temperature inside and outside the buildings during the experiment. Furthermore, mass flow emission rates per animal unit and per cow are presented. To obtain reliable data for emission rates, continuous measurements over longer periods are recommended (Joo et al., 2014). Five days was the minimum measurement period in this experiment. The mean NH3 emissions were 43.2 g cow-1 d-1, 126% of what was obtained using estimated emission factors from the Czech national inventory (34.2 g cow-1 d-1; MZP, 2013). For examle Joo et al. (2014) reported 14 to 35 g cow-1 d-1 for a naturally ventilated freestall barns, and Bluteau et al. (2009) reported 11-19 g cow-1 d-1 for a naturally ventilated barn. The highest value was reported by Rumburg et al. (2008) also in a naturally ventilated barn (110 g cow-1 d-1). In this study, the mass flow emission rate of NH3 was of 1.4 g AU-1 h-1, a lower value than reported by Samer et al. (2012) and Fiedler and Müller (2011) (from 2 to 5 g AU-1 h-1 during summer seasons in a naturally ventilated dairy building) but with a higher number of dairy cows (364 dairy cows).
Table I Air exchange rates and average NH3 emission rates from a dairy barn during a five-day measurement period in June.
| n | Mavg | RHi | RHo | ti | to | V | AERH2O | ENH3AU |
| 240 | 632 | 71.1 | 74.8 | 16.7 | 14.8 | 34.2 | 4.8 | 1.4 |
n: Number of cows housed in the building; Mavg: Average weight of the cows (kg); RHi: Relative humidity inside the building (%); RHo: Relative humidity outside the building (%); ti: Air temperature inside the building (ºC); to: Air temperature outside the building (ºC); V: Ventilation rate (m3 s-1); AERH2O: Air exchange rate subject to H2O-balance (h-1); ENH3AU: Specific mass flow emission rate of NH3 (g AU-1 h-1).
3.2 NH3 concentrations
Monthly mean NH3 concentrations (µg m-3) vs. distance from the source in different wind directions, frequencies and speed during the period May-July are depicted in Fig. 3. Monthly mean NH3 concentrations in different wind directions and wind speed at 50, 100, 150, 300 and 400 m during the period May-July for 12 sampling sites are depicted in Table II. NH3 concentrations decreased rapidly with distance from the dairy barn, with highest concentrations observed in the eastern direction 200 m away from the source, and lowest concentrations observed in the southern direction. One possible reason for the smaller NH3 concentrations at 50 and 150 m from the dairy farm in the latter direction could be attributed to the highest wind speed at our site (with a three-monthly mean of 2 m s−1 at a 2 m height). High wind speed usually causes fast dispersion and dilution of the NH3 plume, and thus low NH3 concentration (Shen et al., 2016). The highest NH3 concentration at 300 m and 400 m from dairy farm in eastern direction could be attributed to the lowest wind speed (with a three-monthly mean of 0.7 m s−1 at a 2 m height) and the lowest wind direction frequency. We recommend to locate tree belts around the source as a strategy to reduce the effects of emission hotspots on forest ecosystems located 300 m northeast and southwest from a dairy farm.

Fig. 3 Mean NH3 concentrations (µg m-3) vs. distance from the source in different wind directions, wind direction frequencies and wind speed during the period (a) May-July, (b) May, (c) June, and (d) July.
Table II Monthly mean NH3 concentrations in different wind directions and wind speeds during the period May-July for 12 sampling sites.
| Month | May | Juny | July | |||||||
| Wind direction | Distance (m) |
Wind speed (m s−1) |
NH3 concentration (µg m-3) |
Wind speed (m s−1) |
NH3 concentration (µg m-3) |
Wind speed (m s−1) |
NH3 concentration (µg m-3) |
|||
| North | 50 | 2.0 | 47.1 | 1.4 | 76.2 | 1.6 | 68.2 | |||
| 100 | 36.6 | 27.9 | 40.6 | |||||||
| 150 | 16.4 | 19.8 | 20.4 | |||||||
| 300 | 8.0 | 8.3 | 8.4 | |||||||
| 400 | 6.0 | 6.1 | 6.4 | |||||||
| East | 150 | 0.9 | 13.9 | 0.6 | 23.9 | 0.7 | 22.9 | |||
| 300 | 10.4 | 17.2 | 11.8 | |||||||
| 400 | 9.1 | 14.7 | 10.1 | |||||||
| South | 50 | 2.0 | 12.0 | 1.7 | 14.7 | 2.3 | 13.2 | |||
| 150 | 5.0 | 5.3 | 6.2 | |||||||
| West | 300 | 0.8 | 6.2 | 1.1 | 3.4 | 1.0 | 5.4 | |||
| 400 | 5.0 | 3.3 | 3.3 | |||||||
Several studies have dealt with the measurement of atmospheric NH3 profiles downwind of a known source. Our results were similar to those of Pitcairn et al. (1998), Theobald et al. (2006), Walker et al. (2008), Sutton et al. (1998, 2011) and Jones et al. (2013), who reported a rapid decrease of NH3 concentrations with increasing distance to livestock farms. Our results show the short-range dispersion of NH3 emitted from a point source. However, short-range dispersion of NH3 emitted from a point depends not only on the surrounding land-cover (Vogt et al., 2013) and the number of animals in the barn (Adrizal et al., 2008; Verhagen and van Diggelen, 2006), but also on the strategies for reducing the effect of emission hotspots on ecosystems by locating tree belts around the sources (Theobald et al., 2001; Dragosits et al., 2006).
3.3 Comparison of NH3 emissions and concentrations
The diurnal variation of NH3 emissions and concentrations through the measurement period at location B, with a maximum in the morning and minima in the afternoon and night, are shown in Figs. 4 and 5. Higher concentrations of NH3 on June 16 and 17 were caused by different meteorological conditions during those days. The diurnal variations of NH3 emission were similar to those reported by Zhang et al. (2005) and Wang et al. (2006). Averaging the inferred hourly emission rates over all of the available data and converting them to daily emission rates lead to the average emission of 10.3 kg NH3 d−1 from dairy barn with a standard error of 0.16 kg NH3 d−1 (n = 91). Hensen et al. (2009) reported a similar emission and concentration pattern and estimated NH3 emissions between 6.4 ± 0.18 kg d−1 (Huang 3-D model) and 9.2 ± 0.7 kg d−1 (Gaussian 3-D model) from a naturally ventilated livestock farm. Figure 6 shows the average NH3 emission rate as an exponential function of the indoor temperature in the cattle building. The emission level increased approximately from 1 to 7 g NH3 h−1 as the temperature increased from 9 to 25 ºC. Exponential dependence of NH3 emissions to indoor temperature was demonstrated by Zhang et al. (2005) over a temperature range of 5-23 ºC and Hensen et al. (2009) over a temperature range of 14-23 ºC.

Fig. 4 Time course of NH3 emissions in the north wind direction and hourly average of NH3 concentrations measured at a location (B) located 100 m from the building.
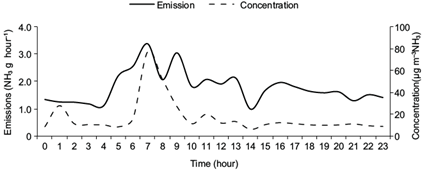
Fig. 5 Mean hourly values of NH3 emissions in the north wind direction and mean hourly values of NH3 concentrations measured at a location (B) located 100 m from the building.

3.4 Total and stomatal NH3 flux
Our mean deposition velocities computed according to Eq. (6) were 0.5 cm s−1 for coniferous forests and 0.4 cm s−1 for crops. Zhang et al. (2009) reported a deposition velocity of 0.5 cm s−1 for coniferous forests from June to July and 0.3 cm s−1 for the crops during spring. Deposition velocities were estimated from big-leaf models using on-site meteorological measurements (Hicks et al., 1987). Mean deposition velocities from 0.2 to 0.4 cm s−1 for agricultural sites were reported by several authors: 0.3 cm s−1 in spring by Cui et al. (2010); 0.2 cm s−1 in summer by Cui et al. (2010) and Zhou et al. (2010); 0.3 cm s−1 by Hayashi et al. (2012); an annual mean of 0.3 cm s−1 by both Cui et al. (2011) and Delon et al. (2012), and an annual mean of 0.4 cm s−1 by Loubet et al. (2011). Mean fluxes to the forest vs. distance from the source in different wind directions, frequencies and wind speeds during the period May-July are depicted in Fig. 7. Mean fluxes to crops vs. distance from the source in different wind directions, frequencies and wind speeds during the period May-July are depicted in Fig. 8. The total flux of NH3 (F total ) to the forest canopy ranged from -0.32 to -0.03 µg NH3 m-2 s-1 at 50 and 400 m, respectively, from the source to the north, which is the dominant downwind direction (Fig. 7a). The stomatal flux of NH3 to the forest canopy ranged from -0.092 to -0.009 µg NH3 m-2 s-1 at distances of 50 and 400 m from the source to the north, which is the dominant downwind direction (Fig. 7b). The total flux of NH3 (F total ) to the crops ranged from - 0.24 to - 0.02 µg NH3 m-2 s-1 at 50 and 400 m, respectively, from the source to the north (Fig. 8a). The stomatal flux of NH3 to the crops ranged from -0.094 to -0.008 µg NH3 m-2 s-1 at 50 and 400 m, respectively, from the source to the north (Fig. 8b). Our estimated range is consistent with studies conducted in Europe and the USA, where deposition fluxes were estimated by Erisman and Wyers (1993), Duyzer et al. (1987), and Phillips et al. (2004). Erisman and Wyers (1993) observed deposition fluxes of 0.0 to 0.45 NH3 m-2 s-1 over heathland near a livestock production facility. Duyzer et al. (1987) reported NH3 fluxes in the range of -0.19 to -0.03 NH3 m-2 s-1 over dry heathland. Phillips et al. (2004) observed the summer season-averaged deposition flux of -0.11 μg NH3 m-2 s-1 over a site with grass or short vegetation near a small swine production facility.

Fig. 7 (a) Total flux of NH3 and (b) stomatal flux of NH3 to the forest vs. distance from the source in different wind directions, frequencies and speeds during the period from May to July.

Fig. 8 Total flux of NH3 (a) and stomatal flux of NH3 (b) to crops vs. distance from the source in different wind directions, frequencies and speed during the period from May to July.
The total flux to forest and the total flux to crops displayed similar spatial patterns, decreasing exponentially with distance from the cow-barn (Figs. 7 and 8), which is in accordance with values reported previously (Fowler et al., 1998; Sutton et al., 1998; Verhagen and van Diggelen, 2006).
The mean F total and F sto from 0 to 400 m from the source respect to the surface type is shown in Fig. 9. F total fluxes range from 0.06 to 0.08 µg N-NH3 m-2 s-1 for crops and forest, respectively. F sto to the forest canopy and mean F sto to crops represent 37 and 41% of F total , respectively. Walker et al. (2008) estimated higher F total fluxes of 0.08 to 0.16 for crops and forest, respectively. Mean F sto to the forest canopy represented 26% of F total in the vicinity of a swine production facility (Walker et al., 2008). Differences between our results and those reported by Walker et al. (2008) are partially attributed to the differences in spatial distribution of forest and crops around the source and to the number of animals (4900 pigs). Assuming an emission factor of 43.2 g NH3 cow-1 d-1 and a mean population of 240 cows, the NH3 dry deposition over the nearest 400 m from the source accounted for approximately 11.5% (1192.3 g NH3) of daily emissions (10368 g NH3). Walker et al. (2008) found that 7.8-13.3% of emissions were deposited within 500 m of the swine facility surrounded by forest and crops. According to Fowler et al. (1998), 3-10% of emissions were deposited within 300 m of a poultry facility surrounded by forest. Asman (1998) and Sutton et al.(1998) found that 2-50% of emissions were deposited within 300 m

4. Conclusion
Inferred emissions were compared with measured concentrations of NH3 to obtain data for the derivation of emission-deposition relationship around a dairy farm. The daily pattern of a source was correlated with the temperature inside a dairy farm. The daily pattern resulted from a combination of the temperature effect on a source concentration and the effect of the building’s ventilation rate. Both NH3 concentration and NH3 total flux displayed similar spatial patterns, decreasing exponentially with distance from the dairy barn. The results show the short-range dispersion of NH3 emitted from a point source, but it may not be the same in other situations, since dispersion of NH3 is dependent on the surrounding land-cover and on the number of animals in a barn. NH3 deposition around a dairy barn is a significant nitrogen input to surrounding crops and forest.

