











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
The ocean is a highly dynamic environment affected by a number of processes that take place at different spatial and temporal scales. The organisms that inhabit there are influenced by local processes, and these processes thus determine, to a great extent, the ecosystem’s productivity at a regional level (Daly and Smith 1993). The ocean surface layer is home to marine phytoplankton, the primary component of the food chain. Being autotrophic organisms, their largest abundance occurs in the well-illuminated ocean layer known as the euphotic zone. Given the importance of light in photosynthesis, irradiance is one of the main factors that modulate phytoplankton abundance (Strickland 1965). However, phytoplankton growth and biomass are limited by nutrient availability in the euphotic zone. In this regard, processes at different temporal and spatial scales become important because they determine the structure of the water column, which is directly related to productivity.
One of the major factors limiting the distribution of phytoplankton is water column stratification. Stratification is defined as the amount of energy required to mix water throughout the water column (Simpson and Bowers 1981); if the latter is strongly stratified, more energy will be required to mix it relative to a less stratified column. Stratification of the water column determines the availability of nutrients that are used for phytoplankton growth. The mixed layer depth (MLD) and mixing intensity are physical parameters related to stratification that strongly affect phytoplankton primary production by determining the exposure of phytoplankton to light and its losses by sedimentation (Jäger et al. 2008). A deep MLD leads to a decrease in phytoplankton abundance because the population is distributed throughout a larger volume of water, that is, dilution (Behrenfeld and Boss 2014). In addition, if light conditions change as a result of vertical movements associated with turbulence (deep MLD), phytoplankton will photosynthesize in accordance to the amount of incident light in the MLD (Marra 1978).
The ocean region off the coast of the Baja California peninsula has been affected by interannual processes. Some of these processes have led to changes in ocean circulation, which in turn have modified the structure of the water column. For example, El Niño 1997-1998 produced a deepening of the pycnocline as a result of the influence of tropical warm water transported by a coastal flow parallel to the coast with a poleward direction (Durazo and Baumgartner 2002). During La Niña 1998-1999, intensification of coastal upwelling events led to increased phytoplankton biomass (Lavaniegos et al. 2002). From 2002 to 2006, the weakening of the Alaska Gyre caused the anomalous intrusion of subarctic water off Baja California (Durazo 2009), leading to a decrease in phytoplankton production and biomass (Gaxiola-Castro et al. 2008, Espinosa-Carreón et al. 2015). The ocean warming caused by “The warm Blob” during 2013 and 2014 and El Niño 2015-2016 produced the highest reduction of phytoplankton biomass and production in the last 13 years at the southern boundary of the California Current (Gómez-Ocampo et al. 2017). However, although phytoplankton production and biomass in this region have responded to these interannual processes, the mechanisms through which these events cause variations in the structure of the water column remain unknown.
Since 1997, the Investigaciones Mexicanas de la Corriente de California (Mexican Investigations on the California Current; IMECOCAL for its acronym in Spanish) program has surveyed the region off Baja California. This has allowed for better knowledge and understanding of the many oceanic physical, biological, and chemical processes in this region (Durazo and Baumgartner 2002, Lavaniegos et al. 2002, 2015; Espinosa-Carreon et al. 2004, Gaxiola-Castro et al. 2008, Gaxiola-Castro 2010, Jeronimo and Gomez-Valdes 2010, Martínez-Gaxiola et al. 2010, Durazo 2015, Espinosa-Carreón et al. 2015, Martinez-Fuentes et al. 2016, Gómez-Ocampo et al. 2017, Lavaniegos 2017). However, although there is available information on physical, biological, and chemical variables, there is still insufficient understanding on how physical processes influence the interannual changes in phytoplankton biomass and production in the water column.
The aim of this article is to describe and relate the interannual variations of phytoplankton biomass and production with changes in the structure of the water column. To this end, the 1997-2016 time series for chlorophyll a, primary production, pycnocline depth, and MLD were analyzed, as well as the stratification indices between the surface and 100 and 200 m (Ф100 and Ф200, respectively) along IMECOCAL line 100 (L100IMEC ). L100IMEC is one of the most intensively sampled lines and, due to its location, it has been found to be representative of the response of the marine ecosystem to interannual processes that occur off the peninsula (Linacre et al. 2010). In addition, in order to understand the relationship between interannual variations in the structure of the water column and phytoplankton production, the relationship between photosynthetic parameters (maximum photosynthetic rate at light saturation [
Material and methods
Data Sources
In order to estimate the average conditions in the study area, data derived from remote sensing were obtained for sea surface temperature (SST), chlorophyll (CHL), and sea surface wind in the southern end of the California Current, off Baja California (28ºN-33ºN, 120ºW-114ºW; Fig. 1). The monthly composite images (spatial resolution of 4 × 4 km) of SST and CHL concentrations derived from the MODIS-Aqua sensor were downloaded from the ERDDAP database (http://coastwatch.pfeg.noaa.gov/erddap/index.html) for the 2003-2015 period. The primary production (PP) data obtained with the Behrenfeld and Falkowski (1997) Vertically Generalized Production Model (VGPM) was obtained from Oregon State University (OSU, http://www.science.oregonstate.edu/ocean.productivity/). The climatology for the mixed layer MLD, with a spatial resolution of 0.5º × 0.5º, was obtained from the Monthly Isopycnal and Mixed-layer Ocean Climatology (MIMOC, http://www.pmel.noaa.gov/mimoc/) produced by the National Oceanic and Atmospheric Administration (NOAA). Wind data considered for this work were obtained from the National Centers for Environmental Prediction (NCEP). The zonal and southern components (0.25º spatial resolution and 1-day temporal resolution) are available on NOAA’s website at https://www.ncdc.noaa.gov. Wind stress curl was calculated from its components according to Trenberth et al. (1990). For all variables, long-period averages (hereafter average conditions) were calculated according to the length of the time period for which data was obtained. From the PP data obtained from OSU, average conditions were calculated for the northern zone off Baja California (28ºN-32ºN) from 1998 to 2002 for data derived from the SeaWiFS sensor and from 2003 to 2016 for data obtained from the MODIS-Aqua sensor (both with a monthly resolution of 18 × 18 km). This climatology was subsequently used to estimate the anomalies from 1998 to 2016, which were defined as the observed value minus the calculated average.
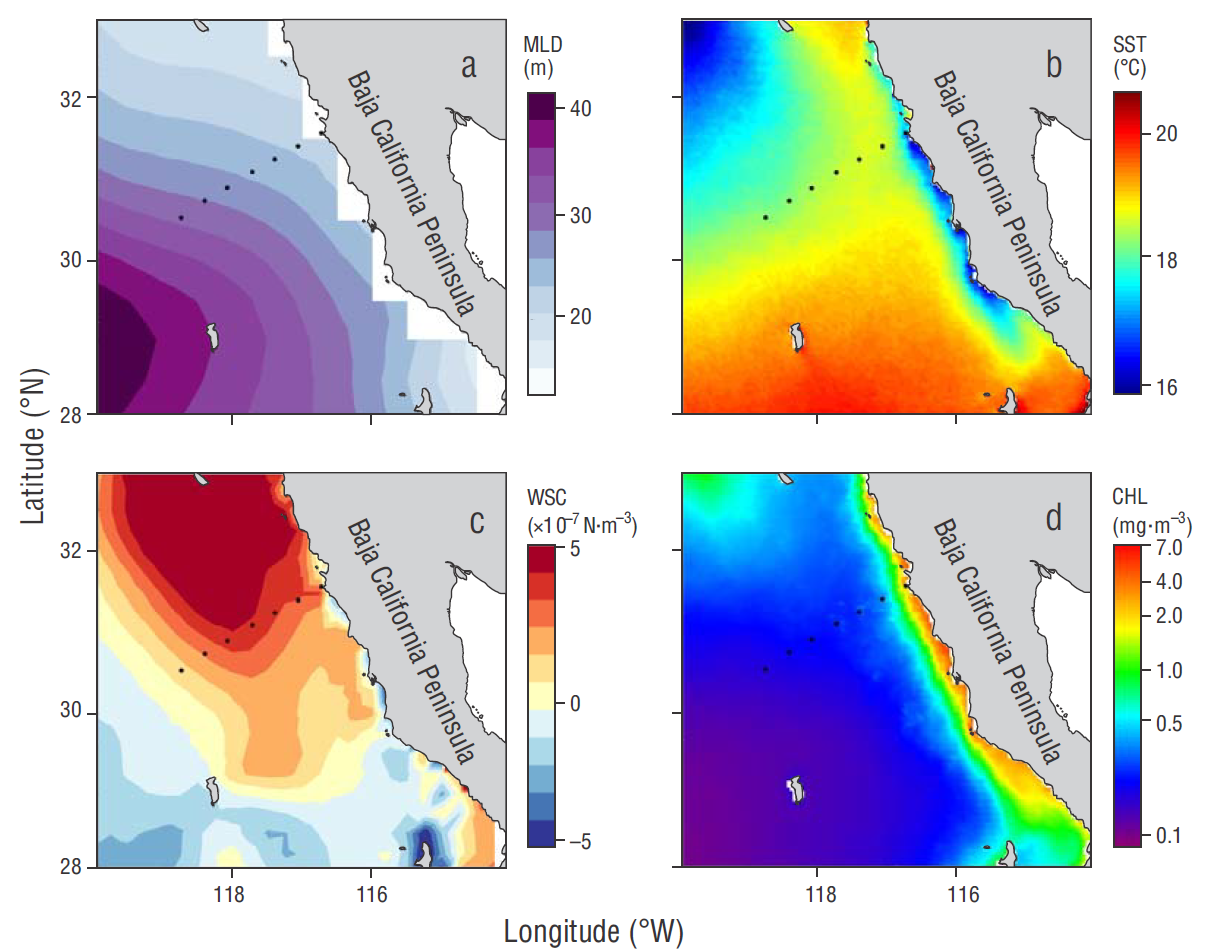
Figure 1 Horizontal distribution of the mean conditions for (a) mixed layer depth (MLD) (b) sea surface temperature (SST), (c) wind stress curl (WSC), and (d) satellite chlorophyll (CHL). Dots indicate the location of the hydrographic stations where in-situ data were collected along line 100 of the Investigaciones Mexicanas de la Corriente de California (IMECOCAL) program.
The interannual variations in the structure of the water column and phytoplankton biomass in oceanic and coastal areas were analyzed using data from the stations located along L100IMEC (Fig. 1). This line was selected because it was the IMECOCAL line representative of the northern zone off Baja California with the largest number of temperature, salinity, and chlorophyll a (Chla) records from 1998 to 2016. Cruise averages were calculated for the variables analyzed at the L100IMEC stations representative of the coastal zone (stations 30 and 35) and the oceanic zone (stations 45, 50, 40, 55, and 60) (Gaxiola-Castro et al. 2010, Durazo 2015). Phytoplankton Chla samples were obtained from water samples collected with Niskin bottles at standard depths (0, 10, 20, 50, and 100 m) and analyzed with the fluorimetric method (Yentsch and Menzel 1963, Holm-Hansen et al. 1965). Temperature and salinity CTD (Sea- Bird) data were used to estimate MLD following the method proposed by Kara et al. (2000) and modified by Jeronimo and Gomez-Valdes (2010) for the IMECOCAL zone. Pycnocline depth ( Z Pyc ) was calculated according to Fiedler et al. (2013) . The stratification parameter (Ф) between the surface and 100 m and the surface and 200 m depth (Ф100 and Ф200, respectively) was calculated according to Simpson and Bowers (1981). Chla was integrated into the water column from the surface to 100 m depth using the trapezoidal rule to estimate integrated Chla (Chla int, mg m-2).
The photosynthesis-irradiance experiments (P-I curves) were carried out onboard during some oceanographic campaigns at oceanic stations, with water samples collected between 10:00 and 14:00, local time, at a depth corresponding to 30% surface irradiance. Water was collected with 5-L Niskin bottles to fill 27 flat 250-mL polystyrene bottles (Nucleon), which were spiked with 100 μL NaH14CO3 (~5 μCi). Bottles were placed in a Morel incubator (Marcel et al. 1994) for ~2 h under a light gradient of 1 to 900 mol quanta m-2 s-1 generated by a 500 W tungsten halogen lamp. Carbon assimilation rate (P, mg C m-3 h-1) was estimated from the incorporation of 14C during incubation, by subtracting the baseline values (time = 0), and was normalized with respect to Chla values measured at a depth of 30% surface irradiance in each experiment (
Statistical analysis
The relationship of trends PP and water column structure was explored using GAMs (Hastie and Tibshirani 1986). This method fits non-linear relationships between variables, which are expressed as a curve from a smoothed function. In contrast with linear models, GAMs allow assigning an exponential distribution (Poisson, binomial, gamma, or normal) to the dependent variable.
In order to explain the variation in PP in terms of phytoplankton physiology, the
Results
Average conditions in the study area
In order to characterize the horizontal distribution of some variables related to water column structure and productivity off Baja California, average conditions in the study area were obtained for MLD, SST, wind stress curl, and surface chlorophyll (Fig. 1). Average MLD and SST show higher values offshore and in the oceanic area to the south of the Baja California peninsula (Fig. 1 a, b). In contrast, average wind stress curl and chlorophyll values derived from satellite imagery were higher in the coastal zone and decreased off-shore and to the south of the peninsula (Fig. 1 c, d). Therefore, 2 typical average conditions were observed in the study area. The first occurs in the coastal area and off the northern part of the peninsula, and seems to be more dynamic according to the wind stress curl values; consequently, the water column is expected to be less stratified. The second, which is less dynamic, occurs mainly in the oceanic area and off the southern part of the peninsula, with conditions typical of a more stratified environment.
1997-2016 time series in L100IMEC
The effect of the interannual variability in water column structure on phytoplankton was characterized from time series of the variables of interest in the L100IMEC coastal and oceanic areas. Particular periods during which the Chla int signal showed obvious variations were selected (Fig. 2a). These periods correspond to the period from 2002 to 2006 (anomalous intrusion of subarctic water), La Niña 2008, and the ocean warming during 2013 to 2016 resulting from “The warm Blob” and El Niño 2015-2016.
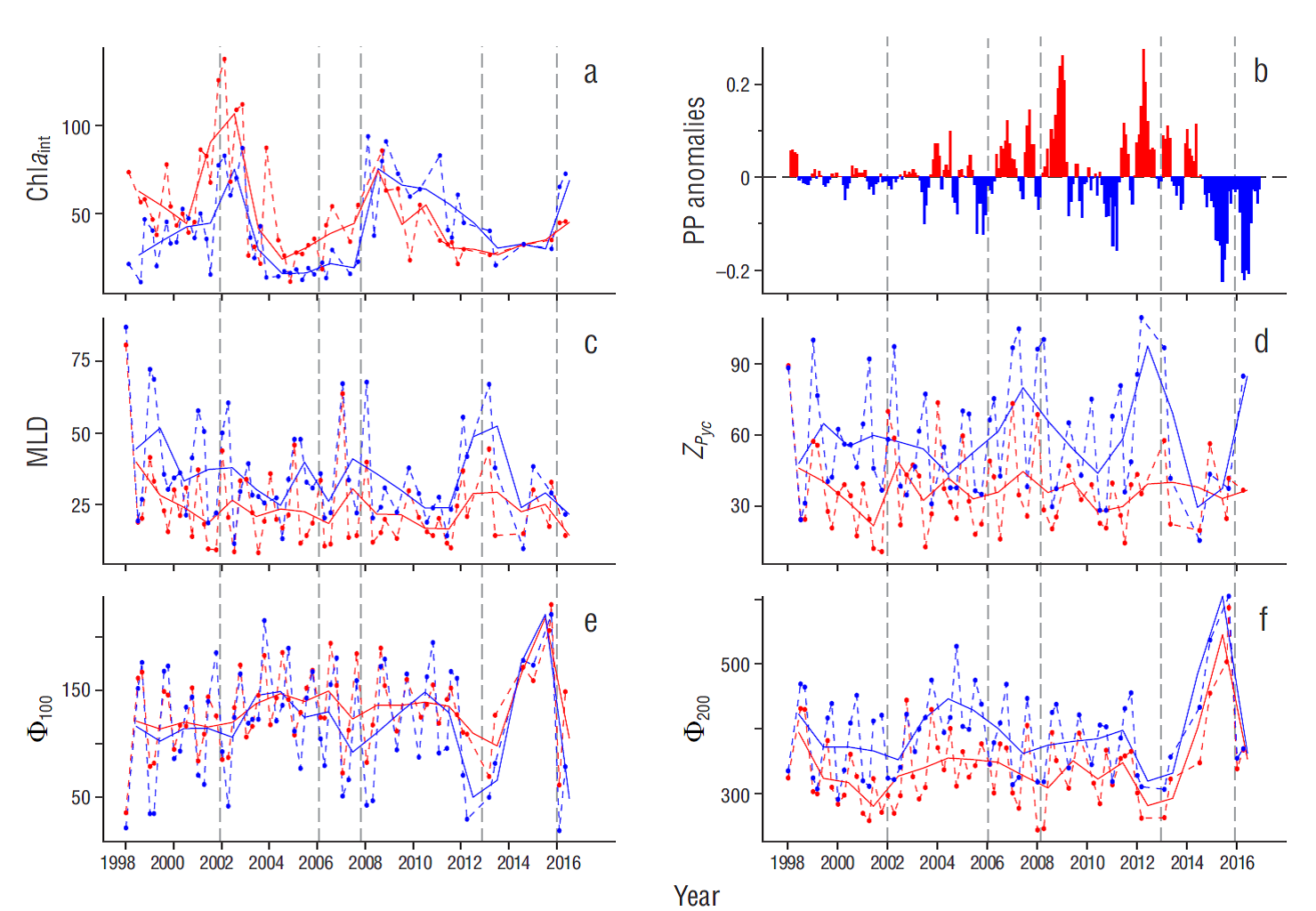
Figure 2 Time series (1998-2016) for (a) water-column integrated chlorophyll a (Chlaint, mgꞏm-2), (b) primary production (PP, g Cꞏm-2ꞏd-1) anomalies obtained from SeaWiFS (1998-2002) and MODIS-Aqua (2003-2016) sensors, (c) mixed layer depth (MLD, m), (d) pycnocline depth (Z Pyc , m), and water column stratification parameter at (e) 100 and (f) 200 m depth (Ф100 and Ф200, respectively; Joules) along IMECOCAL line 100. Colors indicate the time series for coastal (red) and oceanic (blue) stations. Dashed color lines denote the monthly mean and solid color lines represent the yearly mean. Gray dashed lines indicate the periods selected for the analysis..
The time series of the annual Chla int average showed a clear response to interannual events in coastal and oceanic areas (Fig. 2a). The lowest Chla int values (< 40 mg m-2) in the coastal and oceanic areas occurred between 2002 and 2007. In contrast, La Niña 2008 was associated with the maximum Chla int values of the time series in both areas, with similar values in the coastal (~76 mg m-2) and oceanic (~75 mg m-2) areas. From 2008 to late 2016 there was a decline in Chla int values in the oceanic area, reaching minimum values as low as ~30 mg m-2 but slightly higher compared to those observed during the subarctic water intrusion (2002 to 2006). In 2013 Chla int remained relatively unchanged until 2015, when it increased. In the coastal area the lowest Chla int values were recorded between 2011 and 2014 and were similar to those observed during the subarctic water intrusion. In this area, Chla int increased in 2014 until reaching ~45 mg m-2 in 2016.
Phytoplankton production in the northern zone off Baja California showed periods with increasing and decreasing values through the time series (Fig. 2b). During the intrusion of subarctic water, no definite trend was observed. However, conditions during La Niña 2008 led to a rise in PP rates. Afterwards, due to the “The warm Blob” effect during 2014, phytoplankton production dropped and attained even lower levels during El Niño 2015-2016.
MLD was deeper in the oceanic than in the coastal area, but variation was similar in both areas from 1998 to 2016 (Fig. 2c). Aspects worth noting are the reduction in the thickness of the mixing layer during the subarctic water intrusion and the particularly noticeable deepening in the oceanic area in 2004. A period of shallow MLD also took place during La Niña 2008, reaching a minimum value in 2011 in the coastal and oceanic areas (12 and 25 m, respectively). However, the most superficial MLD values in both areas occurred from 2010 to 2012, after which the mixed layer became deeper in the oceanic area until reaching a maximum depth of ~50 m in 2013 under the influence of “The warm Blob”.
Pycnocline depth (Z Pyc ) was more variable in the oceanic than in the coastal area (Fig. 2d). In 2004, during the subarctic water intrusion, there was a slight reduction in Z pyc in the oceanic area (~40 m). The same behavior was observed during 2010, when Z Pyc reached its lowest in both areas (~30 and 45 m in the coastal and oceanic areas, respectively). However, the deepest Z Pyc value (~90 m) was recorded in the oceanic area in 2012, prior to “The warm Blob”. From that year, the pycnocline in this zone became shallower and reached the value closest to the surface in 2014 (~30 m). Subsequently, with the onset of El Niño, Z Pyc deepened in 2015 and reached its maximum value in 2016, with a value similar to the one observed in 2007 (~80 m, first peak in the time series). Although variations were not as marked in the coastal area as in the oceanic area, an aspect worth noting is the slight deepening of the pycnocline during the subarctic water intrusion in 2002 and from 2012 to 2016, under the influence of “The warm Blob” and El Niño.
The stratification indices Ф100 and Ф200 showed a similar variation in both areas (Fig. 2e, f). The time series for both indices reveal an increase in the stratification of the water column during the intrusion of subarctic water (up to Ф100 ~ 150 J m-3 in both areas, and Ф200 ~ 450 J m-3 and Ф200 ~ 350 J m-3 in the oceanic and coastal areas, respectively) and the peak observed in 2015 after the “The warm Blob” and the onset of El Niño 2015-2016 ( Ф100 ~ 225 J m-3 and Ф200 > 500 J m-3 in both areas).
Discussion
Of all the variables analyzed, phytoplankton biomass was the one that best projected the interannual variability due to the various events that influenced the ocean off the northern part of Baja California. Our hypothesis is that high-stratification periods can reduce phytoplankton biomass and affect phytoplankton photophysiology (decreased
Stratification was associated with decreasing and increasing phytoplankton production and biomass during the intrusion of subarctic water, La Niña 2008, and the ocean warming caused by “The warm Blob” and El Niño 2015-2016. A marked water column stratification was observed during the warm periods and the intrusion of sub-arctic water, coupled with the reduction in MLD and Z Pyc . In a poorly mixed water column, phytoplankton can remain in the upper, well-illuminated layer during sufficient time to maintain a population (Huisman 1999). In contrast, when the water column is well mixed, phytoplankton is transported throughout it and, in time, each microorganism experiences the depth -averaged light intensity, which decreases according to MLD. As a result, the depth-averaged specific PP drops as the depth of the water column increases (Huisman 1999, Diehl et al. 2002). In addition, MLD and mixing intensity are related to the accumulation of biomass through the losses of phytoplanktonic cells by sedimentation (Jäger et al. 2008). The probability that a phytoplanktonic organism or a colony sinks below the euphotic zone decreases as mixing intensity increases (Huisman et al. 2004). This occurs because turbulent mixing disperses phytoplankton throughout the water column, which partially offsets sedimentation. In general, losses by sinking affect primarily organisms that sink rapidly (e.g., diatoms) in shallow and poorly mixed water columns (Diehl et al. 2002). In summary, a poorly mixed water column (high stratification) favors PP rates but restrains the accumulation of biomass, as observed in the period of subarctic water intrusion between 2003 and 2005 and during “The warm Blob” in 2013-2014 (Fig. 2).
The largest reduction in PP rates was observed during El Niño 2015-2016, despite the fact that stratification in the northern region off Baja California was the highest in the past 19 years. This may have occurred because, for the first time in history, there was a warm event that preceded an El Niño event (Jacox et al. 2016). The marked deepening of the pycnocline and of the mixed layer during “The warm Blob”, coupled with the high water column stratification during El Niño 2015 (Figs. 2d-f), probably limited the availability of nutrients in the euphotic zone, leading to a decrease in phytoplankton growth rates and hence resulted in lower PP. In addition, the warming of the upper ocean layer (0-100 m) prompted by both events limited phytoplankton photosynthesis.
The depth of the pycnocline and the mixed layer showed greater variability in the oceanic area. This may be due to the prevalence of remote forcings over local forcings (Durazo et al. 2017). In general, in addition to remote signals, the coastal environment is also influenced by local forcings that can mitigate temporal variability. Therefore, Z Pyc and MLD in the oceanic area best represent the interannual variability in the northern zone off Baja California.
In order to assess the relationship between the variability of phytoplankton production and stratification, GAMs were used to determine the relationship between 2 of the characteristic phytoplankton photosynthetic parameters,
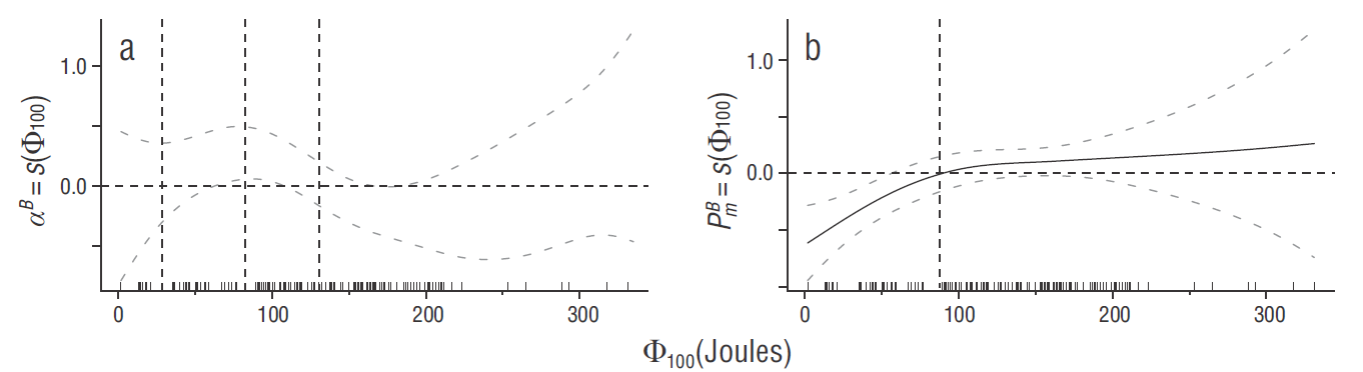
Figure 3 Partial response of (a) the maximum light utilization coefficient (
The above results suggest that PP rates are influenced by stratified environments (Huisman 1999, Diehl et al. 2002). This relationship is supported by the analysis with GAMs, revealing that high stratification is related to peak maximum photosynthetic rates. In addition, it is supported by the temporal variation in the
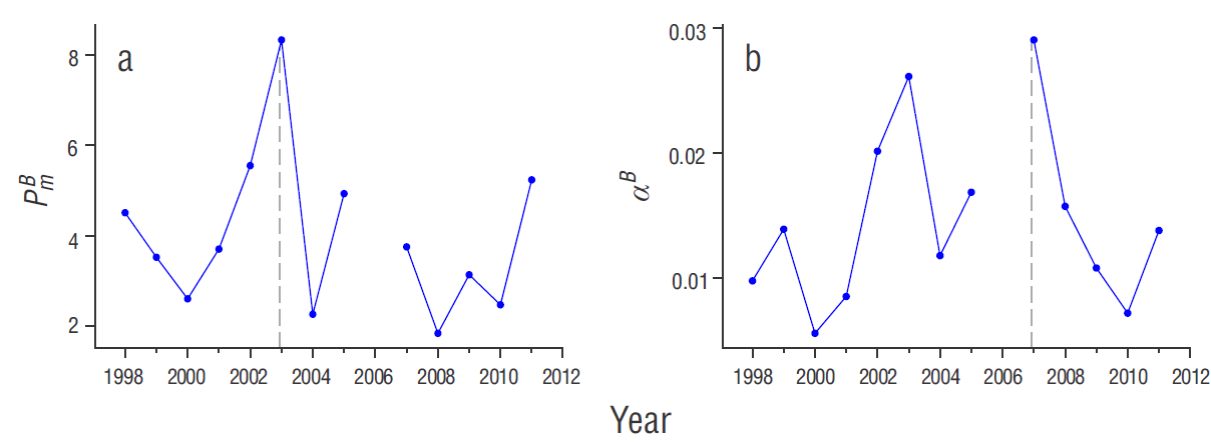
Figure 4 Time series for the average (a) maximum photosynthetic rates ( P m B , mg C [m Chla h]-1) and (b) the maximum light utilization coefficient (α B , mg C [mg Chla]-1 h-1 [ mol quanta m-2 s-1]-1) at the oceanic stations of the IMECOCAL program. Dashed lines indicate the peak P m B and α B values of the time series..
The results of this study support the conclusion that the periods of strongest water column stratification were the subarctic water intrusion from 2002 to 2006 and El Niño 2015-2016, which resulted in the greatest decline in phytoplankton biomass (in both periods) and PP (during El Niño 2015-2016) in the northern zone off Baja California. The photosynthetic parameters

