










Serviços Personalizados
Journal
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versão impressa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.7 no.3 Texcoco Abr./Mai. 2016
Articles
Genotype-environment interaction in extra-large white chickpea seed in northwestern Mexico
1 Campo Experimental Costa de Hermosillo-INIFAP. Pascual Encinas No. 21, Col. La Manga, C.P. 83220, Hermosillo, Sonora. Tel. 0155 38718700. (ortegampedro@gmail.com).
2 Campo Experimental Bajío, Km 6.5. Carretera Celaya-San Miguel de Allende, C. P. 38110, Celaya, Guanajuato. Tel: 0155 38718700. (acosta.jorge@inifap.gob.mx).
3 Campo Experimental Valle de Culiacán-INIFAP. Carretera a El Dorado, km 16.5, C. P. 80430, Culiacán, Sinaloa. Tel: 0155 38718700. (valenzuela.victor@inifap.gob.mx; velarde.sixto@inifap.gob.mx).
4 Campo Experimental Norman E. Borlaug-INIFAP. Calle Norman E. Borlaug, km 12, Valle del Yaqui, C. P. 85000, Cd Obregón, Sonora. Tel: 0155 38 71 87 00. (padilla.isidoro@inifap.gob.mx).
5 Campo Experimental Todos Santos, Agricultura entre México y Durango, Emiliano Zapata, C. P 23050, La Paz, B. C. S. Tel: 0155 38718700. (gutierres.erasmo@inifap.gob.mx).
In Mexico white chickpea (Cicer arietinum L.) grain export large planted in the states of Sonora, Sinaloa and Baja California Sur. In 90% of the sowing area Blanco Sinaloa 92 variety, which has been sown for over 20 years it is used. The aim was to analyze the genotype-environment interaction (IGA) of16 advanced lines and two commercial varieties, and identify locations of low IGA, and lines of higher yield and grain weight (number of grains in 30 g). The 18 genotypes were established during the autumn-winter 2011-2012 under irrigation or temporary (residual moisture) in a randomized complete design in seven environments in Sonora, Sinaloa and Baja California Sur blocks. The IGA analysis was performed using the AMMI model. Yield and grain weight significant differences between environments, genotypes and genotype-environment interaction (p< 0.05) were detected. Environments greater performance were Santo Domingo, BCS with 3.63 t ha-1 under a system ofdrip irrigation and in Navojoa, Sonora, under traditional irrigation with 2.91 tha-1,with significant difference between them and the rest of environments. The highest average yield genotypes were Hoga 02 febrero, 2001 with 2.72 tha-1 and Hoga 06 marzo, 2002 with 2.60 t ha-1, only the first was significantly higher (p< 0.05) witnesses Blanco Sinaloa 92 and Blanoro. By weight of the grain, Culiacan, Sinaloa, Hermosillo, Sonora, and La Angostura, Sinaloa, localities were larger with 41, 47 and 47 grains in 30 g, respectively. The Hermosillo environment was the farthest from the origin, with greater capacity for discrimination, while Culiacan, Navojoa and Santo Domingo were the least interaction and represent environments where you can select stable varieties.
Keywords: Cicer arietinum L.; grain size; irrigation; residual moisture; yield.
En México el garbanzo blanco (Cicer arietinum L.) de grano grande para exportación se siembra en los estados de Sonora, Sinaloa y Baja California Sur. En el 90% de la superficie de siembra se utiliza la variedad Blanco Sinaloa 92, la cual se ha sembrado por más de 20 años. El objetivo fue analizar la Interacción Genotipo-Ambiente (IGA) de 16 líneas avanzadas y dos variedades comerciales, e identificar localidades de baja IGA, y las líneas de mayor rendimiento y peso de grano (número de granos en 30 g). Los 18 genotipos se establecieron durante el ciclo otoño-invierno 2011-2012 bajo condiciones de riego o temporal (humedad residual) en un diseño de bloques completos al azar en siete ambientes en Sonora, Sinaloa y Baja California Sur. El análisis de la IGA se realizó con el modelo AMMI. En rendimiento y peso de grano se detectaron diferencias significativas entre ambientes, genotipos e interacción genotipo-ambiente (p< 0.05). Los ambientes de mayor rendimiento fueron Santo Domingo, BCS con 3.63 t ha-1 bajo un sistema de riego por goteo y en Navojoa, Sonora, bajo riego tradicional con 2.91t ha-1, con diferencia significativa entre ellos y con el resto de ambientes. Los genotipos de mayor rendimiento promedio fueron Hoga 2001-2-2 con 2.72 t ha-1 y Hoga 2002-6-3 con 2.60 tha-1, sólo el primero fue significativamente superior (p< 0.05) a los testigos Blanco Sinaloa 92 y Blanoro. En peso del grano, las localidades Culiacán, Sinaloa., Hermosillo, Sonora, y La Angostura, Sinaloa, fueron las de mayor tamaño con 41, 47 and 47 granos en 30 g, respectivamente. El ambiente Hermosillo fue el más alejado del origen, con mayor capacidad de discriminación, en tanto que Culiacán, Navojoa y Santo Domingo fueron los de menor interacción y representan ambientes donde se pueden seleccionar variedades estables.
Palabras clave: Cicer arietinum L.; calibre del grano; humedad residual; rendimiento; riego.
Introduction
Chickpea big white grain (type "Kabuli") that occurs in the northwestern region of Mexico is exported to Europe, North Africa and the Middle East (Yadav et al., 2007). In the period 2012-2013 Mexico ranked sixth worldwide in chickpea production, with 108 799 t, with India the largest producer with a volume of 1 000 100 t followed by Turkey with 675 000 t. As for export of chickpea in the same period, Mexico ranked third worldwide with 141 382 t, surpassed only by Turkey and Australia (FAOSTAT, 2013). White chickpea production in Mexico for export is concentrated in the areas of irrigation Northwest (Baja California Sur, Sonora and Sinaloa). In the autumn-winter 2013 cycle 123 895 hectares were planted with a production of209 941 tons with an average yield of1.82 t ha-1, with a production value of $ 2 622 million pesos and an average rural price $ 12 492.00 t-1 (SIAP, 2013).
The choice of variety to plant in a particular agro-ecological region, is a central aspect in the production technology of chickpea, this determines the efficiency with which the use of available resources. The proper definition of the environment and the characterization of the response of genotypes in an integrated way, contribute to a better understanding of the average productivity (adaptation) of different genotypes and variations (stability) of them, and you understand and exploit the inevitable genotype x environment interaction (Berger et al., 2007).
A variety or genotype is considered stable or adapted when growing up in various environments presents above-average production, since the environment has an important influence on the expression of performance. Considering these aspects, the production of a cultivating determines the effect of genotype (G), the environment (E) and their interaction (G x E or IGA) (Yan and Kang, 2002). In recent years to better understand the interaction G x E years more complex statistical methods are used that analysis of variance, as is the AMMI model, which combines ANVA with principal components (CP). This method removes the effect of genotype and environment, and major components used to explain the interaction G x E (Romagosa and Fox, 1993). Zobel et al. (1988) also noted that AMMI is a combination of a NOVA for the main effects of genotypes and the environment together with principal component analysis (ACP) of the genotype x environment interaction. The best known is the AMMI biplot, which provides a graphical representation of the genetic part (G) and stability (CP) genotypes simultaneously (Yan and Tinker, 2006). Based on this, the ideals cultivars should have a high genetic value (high output) and a small value in CP (high stability).
The AMMI model has ability to interpret a high number of genotypes in several environments, considers genotypes and environments as additives and linear effects allowing its study by an analysis of variance (ANOVA), while the IGA, considers effects multiplicative, which are analyzed by a principal component analysis (Crossa et al., 1990). Analysis using graphical representations 'biplots' (Gabriel, 1971) allowing obtained: (1) observation, in the same chart genotypes (points) and environments (vectors); and (2) exploring the effects attributable to the interaction of G x E. In biplots patterns, the angles between the vectors representing genotypes and environments show the interaction, and the distances from the origin indicate the degree of interaction that They show genotypes across environments. The aim was to analyze the genotype- environment interaction (IGA) of16 advanced lines and two commercial varieties of chickpea, and identify locations IGA low, and high-performance lines and grain weight (number of grains in 30 g).
Materials and methods
In the autumn-winter 2011-2012 cycle, seven trials white chickpea performance in the states of Sonora, Sinaloa and Baja California Sur were established. In Sonora two established, one in the Coast of Hermosillo (Y1) and another in the Valle del Mayo (Y2), one in the Valley of Santo Domingo in BCS (Y4) in Sinaloa four, one in each locality: Valle del Fuerte (Y3), Culiacan (Y5), Mocorito (Y6), La Angostura (Y7) (Table 1). In Sinaloa trials they were established in November and in other locations in December. The evaluated genotypes were developed in the breeding programs of the National Institute of Livestock Agricultural and Forestry Research (INIFAP on the Costa de Hermosillo (Hoga's) and the Valley of Culiacan (Cuga's), in addition to the varieties used White Sinaloa 92 and Blanoro as commercial controls (Table 2).

Table 2 Main Agronomic features of 18 white chickpea genotypes for export, Culiacan, Sinaloa 2006-2012.

The performance tests were established and conducted according to cultural recommendations in each region, including irrigation (except Mocorito and La Angostura which were conducted with residual moisture temporary) and different fertilization level between environments, according to the type of soil fertility level experimental area, as well as conventional insect control, mainly Heliothis virescens and Liriomyza cicerina.
The size of the experimental plot was four rows of five meters long with 0.8 m spacing between rows, with four replications, the treatments were distributed under a completely randomized design blocks. In each experimental unit in the two central rows grain yield and the size there of it was determined. The latter was determined by weighing 30 grams and count the number of seeds of a sample passed through a mesh 9 mm.
For statistical analysis of grain yield and grain weight a factorial design was used considering environments and genotypes and their interaction, for mean comparison Tukey test was used and only estimation performance interaction is genotype-environment with AMMI model (Additive Effects and Multiplicative Interaction) using the adjusted means to calculate the test Gollob (Burgueño et al., 2001). The statistical model AMMI is:

Where, Yij= average yield of i-th genotype in the j-th environment; μ= effect of general average; Gi= effect of the ith genotype; Aj= effect of the j-th environment; n= number of principal components retained in the model; Bk= singular value for main component;  values of the eigenvectors of genotypes for each major component;
values of the eigenvectors of genotypes for each major component;  values of environments for each main component; Eij= experimental error.
values of environments for each main component; Eij= experimental error.
Results and discussion
The analysis of variance of both characters, yield and seed size showed statistical differences in the effects of both environments, as in the genotypes and their interaction (p< 0.0001) (Table 2). The coefficients of variation were 18 and 6%, respectively, indicating that random variations allow consider as reliable assessments. Variability represented by IGA effects, determined from the ratio of the sum of squares with respect to the sums of squares of the factors under study represented 32% for yield and 19% by weight of the grain, percentages are considered important. This fully justifies the detailed analysis of the IGA (Yan and Tinker, 2006).
Among environments there were significant differences (p< 0.0001) in grain yield (Table 2), differences mainly related to crop management [date of planting, irrigation and precipitation (Table 1)] and the incidence of insects and disease, among them mainly the complex of soil fungi that cause root rots (Arvayo et al., 2011). The differences between genotypes (p< 0.0001) and significant effect of genotype-environment (p< 0.0001) interaction showed the differential response of genotypes to occur in different environmental conditions.
Higher performance environments were Baja California Sur with 3.63 t ha-1 and Navojoa with 2.91 t ha-1, yields statistically different from each other (p< 0.05), and with respect to other environments. Lower yields were obtained in Culiacan and Los Mochis environments with 1.62 and 1.11 t ha-1, respectively (Table 3). In the environment of higher performance in B.C.S. cultivation it was conducted with drip irrigation, without pressure from insects and diseases. In the environment of lower yield, Los Mochis, the crop was affected during the grain filling stage by foliar diseases and lack of irrigation, causing terminal drought. In the case of environments Mocorito and La Angostura, Sinaloa, the culture was established and led the residual moisture provided by rainfall (Table 1), which allowed acceptable yields (Table 3).
Table 3 Analysis of variance combined performance and grain size of18 chickpea genotypes in seven environments in northwestern Mexico 2011-2012.

The highest caliber export between environments was obtained in Culiacan with 41 grains in 30 grams, followed by Costa de Hermosillo and La Angostura with 47 grains in 30 grams; while the lowest grain weight was obtained in Los Mochis and Navojo a with 50 and 57 grains in 30 grams, respectively (Table 4). Within environments there was no association between performance and the caliber of the grain; in fact their joint linear association determined by the method of linear Pearson correlation was r=0.02, not significant.
Table 4 Yield (t ha-1) of 18 white chickpea genotypes in seven rooms of the Northwest region of Mexico, 2011-2012 cycle.

1= Cuga; H= Hoga; 2= ambientes como en el Cuadro 1; 3= separación de medias por Tukey al 0.05.
Genotypes showed higher average yield were Hoga 2001-2- 2 with 2.72 t ha-1, Hoga 2002-6-3 with 067 with Hoga 2.60 and 2.52 (Table 4), while the lowest yields were obtained with genotypes Cuga 08-1090, Cuga 08-3096 and Hoga 2002-40-6 with 2.09, 2.03 and 1.99 t ha-1, respectively. The Hoga 2002-2-2 only significantly superior to commercial varieties Blanco Sinaloa 92 and Blanoro, they showed a similar average yield. The similarity in performance of most experimental genotypes with the witness Blanco Sinaloa 92 (Gomez et al., 2003), in part due to this popular variety is one of the most used in genetic improvement programs parents region of. Through test environments, different varieties occupied the top positions in performance. So contrasting the performance of some genotypes across locations such as lines Cuga 08-3096 and Cuga 08-3160 between the towns of Hermosillo (Y1) and BCS (Y4) response emphasizes the presence of cross-talk, which justify analyzing the IGA.
In the number of grains in 30 g, genotypes with higher average weight were Cuga 08-743, Hoga 067 and Hoga 021 with 45, 45 and 47, the lower weight were Hoga 2002-6-3, Hoga 2002-20-10 and Cuga 09-2067 with 50, 50, and 53 (Table 5).
Table 5 Calibrate grain (number of grains in 30 g) of18 white chickpea genotypes in seven rooms of the Northwest region of Mexico, 2011-2012 cycle.
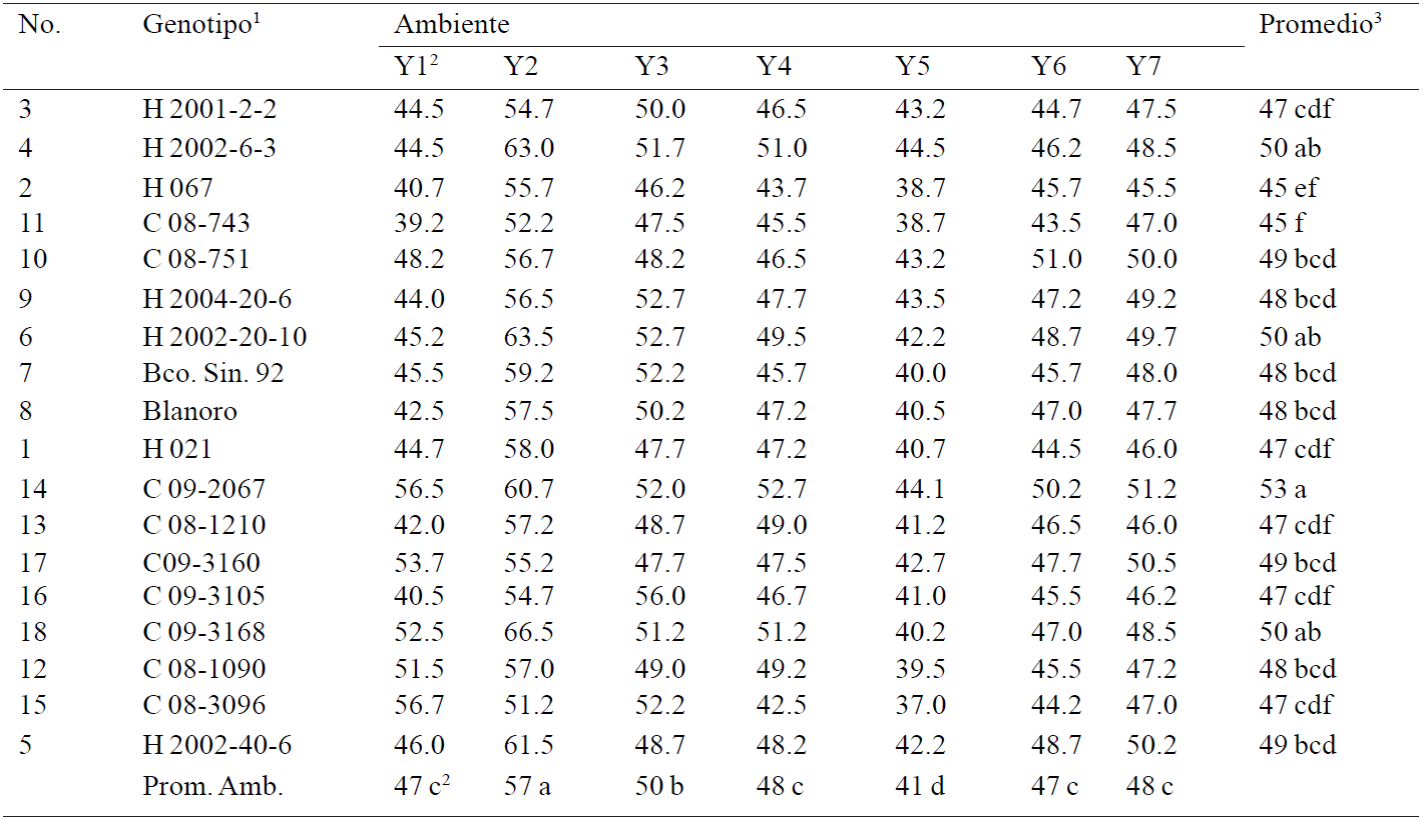
1= Cuga; H= Hoga; 2= ambientes como en el Cuadro 1; 3= separación de medias por Tukey al 0.05.
The average performance and control varieties was similar caliber, same that were only surpassed in yields per Hoga 2001-2-2, this line emerges as candidate for registration as a new variety. The fact that the towns and genotypes with higher yield and grain size were different or contrasting indicates that the potential yield is independent of the grain size; this is the last of the direct components of performance defined (Güler et al., 2001) and depends on the ability of remobilization of each genotype and environmental conditions in the final stage of grain filling.
Analysis of IGA
Analysis of the IGA to yield indicated that the first two principal components (70.1 and 15.6%, respectively) two possible to explain 85% of the phenotypic variation present in the matrix effects of genotypes and IGA, so 14.3% variation was not considered in the analysis (Table 6) Kandus et al. (2010), they found that the sum of squares for the environmental effect was greater than the sum of squares for the effect of interaction GxE more lines, being 48.3% and 43.7% of the sum of total square respectively suggesting data analysis with the SREG model. The first two main components for this model explained 93% of the variance of the data.
Table 6 Principal component analysis, individual component variability and accumulated for the performance of18 chickpea genotypes in seven environments in the Northwest region of Mexico 2011-2012.

1CP= componente principal; GL= grados de libertad; CM= cuadrado medio; (%) IND= porcentaje individual; ACUM= porcentaje acumulado; Fc;= prueba de F calculada; P> F= significancia estadística.
Regarding environments, Hermosillo (Y1) represents the farthest from the origin, where most IGA is expressed, whereas Culiacan (Y5), Navojoa (Y2) and BCS (Y4) environments are closest to the origin, i.e. those where the interaction showed lower expression and represent locations where you can select more reliable (stable varieties). In fact the variety Blanco Sinaloa 92, was developed in Culiacan (Gomez et al., 2003).
One of the merits G x E biplot model is that it can graphically display which was better and that atmosphere in a data set (Yan, 2000). With the five genotypes furthest from the origin (0, 0), Hoga 2002-6-3 (4), Blanoro (8), Hoga 2002-20-10 (6), Cuga 08-3096 (15) and Cuga 09-3105 (16), a polygon so that the marks of the other genotypes were inside the polygon was drawn. On each side of the polygon perpendicular lines they were drawn and match them to the five sectors of the biplot origin were generated (Figure 1). The genotype at the apex of each sector is the most adapted to the environments included in this sector, this only if the first two CP accounted for the largest proportion of the effects of the IGA (Yan and Tinker, 2006), as happened in this studio. In this case, as the data set only includes information of a year, it is not possible to infer anything about the IGA through years, essential for correct identification of stable varieties requirement through time and space, therefore there to be cautious in interpreting the biplot (Yang et al., 2009). However, the information obtained is useful for genetic improvement in the region as the towns of lower yield Mocorito (heavy rain) and Los Mochis (low rainfall) and Hermosillo, in contrast to the rest they showed stress environmental drought or disease, which identified the limitations of genotypes.

Figure 1 Representation of genotypes and environments over the first two axes of major components of aMMI analysis of the average yield of18 chickpea genotypes evaluated in seven environments Northwest Mexico.
One sector of Figure 1 includes genotypes: Cuga 08-3096 (15), Cuga 08- 1090 (12), Cuga 08-1210 (13), Cuga 09- 2067 (14), Cuga 09-3160 (17) and Cuga 09-3168 (18), presented in the environment adaptation Y2 (Navojoa), in this representative May Valley environment, genotype Cuga 08-3096 (15) located in the vertex of the polygon, is the best option for your specific adaptation. Culiacan (Y5) and Mocorito (Y6) genotype Cuga 09-3105 (16) located in the corresponding vertex, is the most specific adaptation. The Hoga 2002-2-2 (3), Hoga 2002-6-3 (4), Cuga 08 743 (11) and Blanco Sinaloa 92 (7), genotypes exhibit excellent adaptation in Angostura.
Hoga genotypes 021 (1), Hoga 067 (2), Hoga 2004-20-6 (9) and Cuga 08 751 (10) are genotypes which, being closer to the origin represent larger or wide adaptation to test environments. In environments the BCS (Y4) and Mochis (Y3) genotypes Hoga 2002-40-6 (5), Blanoro (8) y Hoga 2002-20-10 (6), had excellent adaptation (Figure 1). Hoga 021 (4) is the highest performance in Angostura (Y7) and Mocorito (Y6) environments, and is near Hoga 2001-2-2 genotypes (3) and Cuga 08-743 (11); average of all trials, these genotypes are higher yield potential. For genotypes 6 and 7 to have the same behavior, shown in Figure only the second.
In the town of Los Mochis, the Hoga 2002-20-10 line, located at the top of the section, was the highest performance. The average yield obtained in this locality was low this was because during the crop development was pressure by foliar diseases: gray mold (Botrytis cinerea Pers Fr. Ex.), Powdery mildew [Leuveillula taurica Salmon (Lev.)] y wilting Fusarium [Fusarium oxysporum Schlechtend. Emend. Snyd. Hans et f. sp. ciceris (Padwick) K. Sato et Matuo] (data not shown). Also in the city of Hermosillo, which was the furthest from the origin (Figure 1), there was strong pressure from black root rot [Fusariumsolani (Mart.) Sacc.], Mainly affecting CUGA genotypes from the program Sinaloa improvement. The occurrence of diseases, the towns of Los Mochis and Hermosillo can be considered high capacity of discrimination of genotypes and used to practice selection against these diseases.
Multiple disease resistance is considered as an important component of the comprehensive adaptation (Chataika et al., 2010).The low yields in specific locations with out moisture deficits and soil fertility was due to pressure from specific diseases such as occurred in Los Mochis and Hermosillo. Earliness to flowering mentioned as an important feature in the wide adaptation (Berger et al., 2006), especially in environments where the culture was grown under conditions of residual moisture with high possibility of terminal drought stress. In this study, including all genotypes they are phenology and similar grain size. In fact, the focus on the recovery of grain characteristics, size and color in segregating populations, reduces diversity used in breeding programs in advanced testing lines.
Conclusions
The environmental effect and genotype by environment interaction were greater for performance than for the grain size, indicating the instability of the performance and stability of grain weight across environments.
The environments and their ability to interact with the response of genotypes, genotypes allowed the identification of specific adaptation to similar environments.
Hoga 021 was the genotype most Hoga 2002-2-2 adaptation and the highest average yield.
The atmosphere of the Coast of Hermosillo represented the farthest from the origin, where most IGA said. Less interaction environments, where you can select more successful stable varieties were Culiacan and La Angostura in Sinaloa and Navojoa, Sonora.
Literatura citada
Arvayo, O. M.; Esqueda, M.; Acedo, F. E.; Sánchez, A. and Gutiérrez, A. 2011. Morphological variability and races ofFusarium oxysporum f. sp. ciceris associated with chickpea (Cicer arietinum) crops. Am. J. Agric. Biol. Sci. 6(1):114-121. [ Links ]
Berger, J. D.; Ali, M.; Basu, P. S.; Chaudhary, B. D.; Chaturvedi, S. K.; Deshmukh, P. S.; Dharmarah, P. S.; Dwivedi, S. K.; Gangadhar, G. C.; Gaur, P. M.; Kumar, J.; Pannu, R. K.; Siddique, K. H. M.; Singh, D. N.; Singh, D. P.; Singh, S. J.; Turner, N. C.; Yadava, H. S. and Yadav, S. S. 2006. Genotype by environment studies demonstrate the critical role of phenology in adaptation of chickpea (Cicer arietinum L.) to high and low yielding environments of India. Field Crops Res. 98(2):230-244. [ Links ]
Berger, J. D.; Speijers, J.; Sapra, R. L. and Sood, U. C. 2007. Genotype by environment interaction and chickpea improvement. In: chickpea, breeding and management. Yadav, S. S.; Redden, R. J.; Chen, W. and Sharma, B. (Eds.). CAB Intl. Wallinford, Oxfordshire OX108DE. UK. 72-100 pp. [ Links ]
Burgueño, J.; Crossa, J. and Vargas, M. 2001. SAS PROGRAMS for graphing GE and GGE biplots. International Maize and Wheat Improvement Center (CIMMYT) (consultado enero, 2010). htttp:www.cimmyt.org/biometrics . [ Links ]
Chataika, B. Y. E.; Bokosi, J. M.; Kwapata, M. B.; Chirwa, R. M.; Mwale, V. M.; Mnyenyembe, P. and Myers, J. R. 2010. Performance ofparental genotypes and inheritance of angular leafspot (Phaeosariopsis griseola) resistance in the common bean (Phaseolus vulgaris). Afr. J. Biotechnol. 9(28):4398-4406. [ Links ]
Crossa, J.; Vasal, S. K. and Beck, D. L. 1990. Combining ability study in diallel crosses of CIMMYT’s tropical early and intermediate maturity maize germplasm. Maydica 35(3):279-285. [ Links ]
FAO-FAOSTAT. 2013. http://faostat.fao.org/site/339/default.aspx. [ Links ]
Gabriel, K. R. 1971. Biplot display of multivariate matrices with application to principal components analysis. Biometrika. 58 (3): 453- 467. [ Links ]
Gauch, H. G. and Zobel, R. W. 1996. AMMI analysis ofyield trials. In: genotype-by-environment interaction. Kang, M. S. and Gauch, H. G. (Eds.). CRC Press. Boca Raton, FL, USA. 85-122 pp. [ Links ]
Gómez, G. R. M.; Gómez, G. L. y Salinas, P. R. A. 2003. Blanco Sinaloa 92, variedad de garbanzo blanco para exportación. Campo Experimental Valle de Culiacán; Centro de Investigación Regional Noroeste; Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias. Folleto técnico Núm. 24. 20 p. [ Links ]
Güler, M.; Adak, M. S. and Ulukan, H. 2001. Determining relationships among yield and some yield components using path coefficient analysis in chickpea (Cicer arietinum L.). Eur. J. Agron. 14(2):161-166. [ Links ]
Kandus, M.; Almorza, D; Boggio, R. R. y Salerno, J. C. 2010. Statistical models for evaluating the genotype-environment interaction in maize (Zea mays L.). Phyton, Int. J. Exp. Bot. 79: 39-46. [ Links ]
Romagosa, M. and Fox, P. N. 1993. Integration of statistical and physiological adaptation in barley cultivars. Theor. Appl. Gen. 86:822-826. [ Links ]
SAGARPA-SIAP. 2013. Estadísticas agrícolas de la producción agropecuaria por cultivo. http//www.siap.gob.mx/cierre-de- la-producción-agrícola-por-cultivo/. [ Links ]
Yadav, S. S.; Longnecker, N.; Dusunceli, S.; Bejiga, G.; Yadav, M.; Rizvi, A. H.; Manohar, M.; Reedy, A. A.; Xaxico, Z. and Chen, W. 2007. Uses. Consumption and utilization. In: chickpea, breeding and management. Yadav, S. S.; Redden, R. J.; Chen, W. and Sharma, B. (Eds.). CAB Intl. Wallinford, Oxfordshire OX108DE. UK. 72-100 pp. [ Links ]
Yan, W.; Hunt, L.A.; Sheng, Q. and Szlavnics, Z. 2000.Cultivar evaluation and mega-environment investigation based on the GGE biplot. Crop Sci. 40(3):597-605. [ Links ]
Yan, W. and Kang, M. S. 2002. GGE biplot analysis: a graphical tool for breeders, geneticists and agronomists. CRC Press. Boca Raton, FL, USA. 271 p. [ Links ]
Yan, W. and Tinker, N. A. 2006. Biplot analysis ofmulti-environment trial data: principles and applications. Can. J. Plant Sci. 86(3):623-645. [ Links ]
Yang, R. C.; Crossa, J.; Cornelius, P. L. and Burgueño, J. 2009. Biplot analysis of genotype × environment interaction: Proceed with caution. Crop Sci. 49(5):1564-1576. [ Links ]
Zobel, R. W.; Wright, M. J. and Gauch, H. G. 1988. Statistical analysis of a yield trial. Agron. J. 80(3):388-393. [ Links ]
Received: November 2015; Accepted: March 2016
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
