










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkCiencias marinas
versão impressa ISSN 0185-3880
Cienc. mar vol.34 no.2 Ensenada Jun. 2008
Artículos
Estructura de la columna de agua y perfiles de biomasa fitoplanctónica en el Golfo de México
Water column structure and phytoplankton biomass profiles in the Gulf of Mexico
RM Hidalgo-González y S Alvarez-Borrego*
División de Oceanología, Centro de Investigación Científica y de Educación Superior de Ensenada (CICESE), Km 107 Carretera Tijuana-Ensenada, Ensenada, Baja California, México. * E-mail: alvarezb@cicese.mx
Recibido en enero de 2008.
Aceptado en mayo de 2008.
Resumen
Los sensores remotos proveen información sobre las concentraciones medias de pigmentos fotosintéticos del ~22% superior de la zona eufótica. Para modelar la producción primaria de la columna de agua a partir de datos de pigmentos fotosintéticos derivados de satélite es menester estimar la distribución vertical de la concentración de los pigmentos. Se utilizó una función de distribución Gausiana para representar los perfiles verticales de pigmentos con cuatro parámetros. Se usaron 352 perfiles de concentración de clorofila (Chlz) generados en cruceros oceanográficos para estimar estos parámetros para épocas del año y regiones en el Golfo de México. Se llevó a cabo un análisis de conglomerados con datos superficiales de temperatura y clorofila, Chlz en el máximo profundo de clorofila (DCM), y profundidad del DCM (Zm) para dividir el Golfo de México en tres regiones y el año en dos épocas. A las épocas se les denominaron fría y caliente. Se desarrollaron modelos de regresión para cada región y época para estimar Zm como una función de la profundidad de la capa de mezcla o un índice de estratificación. No fue posible generar modelos de regresión útiles para predecir los otros tres parámetros Gausianos. Por lo tanto, se calcularon sus medias representativas para cada región y época del año. El suponer un perfil homogéneo de biomasa fitoplanctónica resultó en la subestimación de la producción primaria integrada hasta en un 43% con respecto a los valores calculados a partir de perfiles no homogéneos.
Palabras clave: clorofila, perfiles verticales, Golfo de México, sensores remotos.
Abstract
Remote sensors provide information on the average photosynthetic pigment concentrations for the upper 22% of the euphotic zone. To model primary production in the water column from satellite-derived photosynthetic pigments, estimates of the vertical distribution of pigment concentration are required. A Gaussian distribution function was used to represent the pigment vertical profiles with four parameters. We used 352 chlorophyll concentration (Chlz) profiles generated during oceanographic cruises to the Gulf of Mexico to estimate these parameters for seasons and regions within the gulf. Cluster analyses of data on surface temperature and chlorophyll, Chlz at the deep chlorophyll maximum (DCM), and the depth of the DCM (Zm) were used to divide the Gulf of Mexico into three regions and the year into two seasons (cool and warm). Regression models were developed for each region and season to estimate Zm as a function of the mixed layer depth or an index of stratification. We were not able to generate useful regression models to predict the other three Gaussian parameters. Thus, representative means of these parameters were calculated for each region and season. Assuming a homogeneous biomass profile resulted in underestimation of integrated primary production by as much as 43% relative to the values derived from the nonhomogeneous profiles.
Key words: chlorophyll, vertical profiles, Gulf of Mexico, remote sensing.
Introducción
Los sensores remotos proveen información sobre las concentraciones medias de pigmentos fotosintéticos para la primera profundidad óptica (el ~22% superior de la zona eufótica) (Kirk 1994). Se han comparado algoritmos empíricos y semianalíticos para estimar la producción primaria a partir de datos de pigmentos fotosintéticos derivados de satélite (Platt y Sathyendranath 1993, Behrenfeld y Falkowski 1997). En estos modelos la producción primaria se expresa en función de la concentración de clorofila a para cada profundidad Z (Chlz), la cual representa la biomasa fitoplanctónica debido al papel central que ésta juega en los procesos fotosintéticos y porque es la variable biológica que se monitorea con mayor facilidad desde el espacio exterior (Platt y Sathyendranath 1993). Estos modelos de producción primaria se aplican a toda la zona eufótica e idealmente deben usar todo el perfil vertical de Chlz (Morel y Berthon 1989). Por ende, existe una brecha entre la información limitada del satélite y lo que se requiere al modelar.
El suponer una capa de mezcla con una distribución vertical homogénea de Chlz puede producir estimaciones inexactas de la producción primaria integrada (PPint) (Platt et al. 1991). El máximo profundo de clorofila (DCM) es una característica presente en todo el océano (Dandonneau 1979, Cullen y Eppley 1981). Desde sus primeros estudios se ha puesto énfasis en entender a qué se debe el DCM (Varela et al. 1992 y las referencias ahí citadas). Generalmente, el tomar en cuenta la presencia del DCM aumenta las estimaciones de PPint y debido a que aquel a menudo aparece por debajo de la capa de mezcla, es muy posible que la mayoría de la producción sea nueva (Sathyendranath et al. 1995). Lewis et al. (1983) y Platt et al. (1988) propusieron una función Gausiana para representar el perfil vertical de Chlz. Un problema que se presenta al estimar la producción oceánica a partir de mediciones superficiales es que hay diferencias regionales en la forma de la distribución vertical de Chlz. La suposición implicada en la distribución Gausiana es que para una región oceánica dada, en una época o estación del año, la forma típica del perfil vertical de Chlz es estable. Por lo tanto es necesario usar datos históricos de Chlz para caracterizar los parámetros de la función Gausiana para cada provincia o región oceánica.
Millán-Núñez et al. (1997) usaron datos históricos del programa CalCOFI para generar un conjunto de modelos de regresión para calcular los parámetros Gausianos a partir de valores superficiales de clorofila (Chls), para diferentes regiones del Sistema de la Corriente de California, y para diferentes épocas del año. Hidalgo-González y Álvarez-Borrego (2001) usaron datos históricos de Chlz para ajustar una función Gausiana para representar el perfil medio de Chlz para diferentes épocas del año y regiones del Golfo de California. Millán-Núñez et al. (1997) e Hidalgo-González y Álvarez-Borrego (2001) concluyeron que suponer un perfil homogéneo de Chlz resulta en una subestimación de la PPint hasta en un 30% en el Sistema de la Corriente de California, y hasta en un 60% en el Golfo de California, con respecto a los valores obtenidos a partir de perfiles no homogéneos.
El propósito de este trabajo fue relacionar empíricamente los parámetros Gausianos con propiedades que se pueden medir con sensores en satélites, tales como la concentración de clorofila (Chlsat) y la temperatura superficial (Ts ), así como también relacionar estos parámetros con propiedades físicas como la profundidad de la capa de mezcla y un índice de estratificación, para regiones del Golfo de México. Se exploró el efecto de considerar perfiles no homogéneos de Chlz versus perfiles homogéneos, con la misma Chls, en los cálculos de producción primaria. La intención no es predecir perfiles de Chlz para estimar la producción primaria local instantánea, sino proveer medias representativas de perfiles verticales de Chlz para estimar la producción para una época completa del año y para toda una región del golfo. Morel y Berthon (1989) indicaron que no es razonable, y posiblemente sea hasta superfluo, visualizar el uso de modelos de producción fotel por fotel cuando se interpretan las imágenes de satélite.
Métodos y materiales
El Golfo de México es un mar marginal profundo localizado entre 18° y 30° N, y 81° y 98° W; está conectado con el Mar Caribe a través del Canal de Yucatán y con el Atlántico Norte a través del Estrecho de Florida. El mecanismo forzante principal de la circulación del Golfo de México es el transporte asociado al flujo que entra por el Canal de Yucatán (80 cm s-1). Este flujo alimenta la característica oceanográfica principal del golfo: la Corriente de Lazo (~44 cm s-1) (Monreal-Gómez and Salas-De-León 1997). Esta corriente es parte del Sistema de la Corriente del Golfo (Stommel 1966). Entra al golfo a través del Canal de Yucatán (la profundidad del umbral es ~2040 m) gira anticiclónicamente, y sale a través del Estrecho de Florida (profundidad del umbral ~800 m). La surgencia a lo largo de la orilla de la Corriente de Lazo es una fuente importante de nutrientes a la zona eufótica. El aumento de Chlsat a lo largo de la orilla de la Corriente de Lazo, detectado en imágenes de satélite, atestigua la importancia de este proceso (Müller-Karger et al. 1991). Los datos de temperatura superficial de 1946 a 1998 sugieren que la variación interanual es pequeña en todo el golfo.
A medida que la Corriente de Lazo penetra hacia el norte en el golfo, su trayectoria se hace inestable y se desprenden anillos (Hurlburt y Thompson 1980). Estos anillos son hasta de 400 km o más de diámetro, y se propagan lentamente hacia el oeste. Parece existir una relación clara entre los flujos de vorticidad a través del Canal de Yucatán y el comportamiento de la Corriente de Lazo. El desprendimiento de anillos anticiclónicos de la Corriente de Lazo es precedida por periodos de aporte de vorticidad positiva (ciclónica) del Mar Caribe al Golfo de México, lo cual causa que la Corriente de Lazo se retraiga, y no se debe, como podría pensarse, al aporte y acumulación de vorticidad negativa (anticiclónica), los cuales incrementan el crecimiento de la Corriente de Lazo y su extensión en el golfo (Candela et al. 2002).
El ecosistema del Golfo de México tiene un amplio espectro de productividad fitoplanctónica, desde aguas costeras eutróficas hasta condiciones oligotróficas en las aguas oceánicas profundas (Lohrenz et al. 1999). Los procesos químicos y físicos principales que afectan la productividad incluyen la influencia de un sistema de ríos grandes y la Corriente de Lazo con penetración altamente variable en el norte y este del Golfo de México (Wiseman Jr. y Sturges 1999). En general, datos recientes apoyan lo reportado por El-Sayed y Turner (1977), en el sentido de que pico y nanofitoplancton contribuyen con más de tres cuartas partes de la abundancia de fitoplancton y más de dos tercios de la producción primaria del agua profunda (v.g., Signoret et al. 2006, Sánchez-Iturbe et al. 2006, Hernández Becerril et al. 2008). El Río Mississippi es el más grande de Norteamérica, y algunos de los valores más altos de producción primaria medidos con 14C en la zona costera norte (hasta 5 gC m-2 d-1) son atribuibles al aporte de nutrientes de los ríos Mississippi y Atchafalaya (Lohrenz et al. 1990).
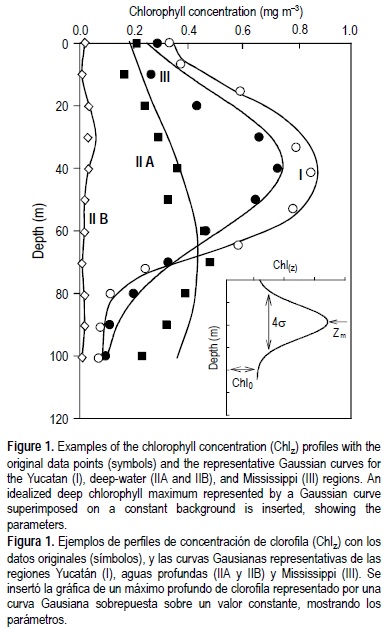
Para el presente trabajo se utilizaron datos históricos generados en cruceros de 1980 a 2000. La concentración de clorofila a se determinó por el método fluorométrico (Holm-Hansen et al. 1965) en la mayoría de los cruceros. En los cruceros de 1997, 1999 y 2000 se utilizaron mediciones de fluorescencia natural para generar perfiles continuos de Chlz (Barreiro-Güemez et al. 2003, Salas-De-León et al. 2004, Signoret et al. 2006). Los perfiles con más de un máximo de clorofila, y aquellos con menos de cuatro profundidades muestreadas, se descartaron (8%). La ecuación de Platt et al. (1988) se ajustó al resto de los perfiles de clorofila (352):

donde Chlz es la concentración de clorofila (mg m-3) en la profundidad Z (m), Chlo es la concentración base por debajo de la curva, h es la integral bajo la curva (mg m-2) por arriba de Chlo, σ controla el grosor de la capa del DCM, y Zm es la profundidad del máximo de clorofila (Chlm). La figura 1 muestra algunos ejemplos de perfiles de Chlz con las curvas Gausianas ajustadas. En la figura 1 se puede apreciar que en el caso de los perfiles de Chlz del Golfo de México no es necesario mejorar la ecuación 1 agregándole una variación lineal de Chlz bajo la curva Gausiana como propusieron Matsumura y Shiomoto (1993).
El parámetro de estratificación  fue una de las variables físicas utilizadas para caracterizar el DCM, donde
fue una de las variables físicas utilizadas para caracterizar el DCM, donde  s es la cantidad de energía mecánica requerida para llevar a cabo la mezcla vertical (J m-3; cero para una capa bien mezclada y aumenta con la estratificación), z es la coordenada vertical (positiva hacia arriba), k es la profundidad de integración en la columna de agua (k = -100 m), ρ es la densidad del agua,
s es la cantidad de energía mecánica requerida para llevar a cabo la mezcla vertical (J m-3; cero para una capa bien mezclada y aumenta con la estratificación), z es la coordenada vertical (positiva hacia arriba), k es la profundidad de integración en la columna de agua (k = -100 m), ρ es la densidad del agua,  es la ρ media para el intervalo 0 a -100 m, g es la aceleración de la gravedad, y Δz = 1 m (Simpson y Hunter 1974). La capa de mezcla (MLD, m) se calculó de acuerdo con Martínez-Sepúlveda (1994). Se consideró como la porción de la columna de agua de la superficie hasta la profundidad donde el gradiente vertical de temperatura se mantuvo ≤ 0.07°C m-1 y su fondo se definió como el punto donde la diferencia de temperatura con respecto a la de 10 m fue 1°C.
es la ρ media para el intervalo 0 a -100 m, g es la aceleración de la gravedad, y Δz = 1 m (Simpson y Hunter 1974). La capa de mezcla (MLD, m) se calculó de acuerdo con Martínez-Sepúlveda (1994). Se consideró como la porción de la columna de agua de la superficie hasta la profundidad donde el gradiente vertical de temperatura se mantuvo ≤ 0.07°C m-1 y su fondo se definió como el punto donde la diferencia de temperatura con respecto a la de 10 m fue 1°C.
Se tabularon los cuatro parámetros Gausianos, Ts, Chls, clorofila en el DCM (Chlm), s, y la profundidad de la capa de mezcla, para un total de 352 estaciones hidrográficas. La tabla 1 y la figura 2a muestran la distribución geográfica y temporal de estas estaciones hidrográficas. El análisis de conglomerados de Ts, Chls, Chlm y Zm agrupó las estaciones hidrográficas en tres regiones diferentes (fig. 2b). La región Yucatán (I) es la zona costera al sur de la isóbata de 200 m y adyacente a la Península de Yucatán; la región de aguas profundas (IIA y IIB) comprende la parte central del Golfo de México, con aguas con profundidades > 200 m; y la región norteña se denominó región Mississippi (III) y comprende la zona costera al norte de la isóbata de 200 m, de Texas a Florida. Los perfiles de Chlz en la zona costera estrecha con profundidades <200 m en el occidente del golfo, adyacente a Tamaulipas y Veracruz, se comportaron como los de la región de aguas profundas, por lo que, para el propósito de caracterizar la forma media del perfil de biomasa fitoplanctónica, esta área costera se incluyó en la región de aguas profundas. En la parte oriental del golfo los perfiles de Chlz son muy escasos. Sin embargo, con base en las condiciones oceanográficas se extrapoló la región Mississippi para incluir la zona costera nororiental hasta Florida. El análisis de conglomerados aplicado sólo a Zm produjo la misma división del golfo en tres regiones. La forma de los perfiles de clorofila se comportó esencialmente igual a lo largo del año para las regiones Mississippi y Yucatán, pero el resultado del análisis de conglomerados de Zm para la región de aguas profundas sugiere la división del año en dos épocas, con invierno y primavera como época fría (IIA) y verano y otoño como época caliente (IIB). Las medias de las variables y parámetros para las tres regiones y, en su caso, las dos épocas (presentadas en la tabla 2) se compararon para probar si las diferencias fueron significativas a un nivel de confianza de 95%. Se aplicó análisis de regresión lineal simple para generar modelos de Zm, h, Chlo y σ como funciones de Chls, Ts, s y/o la profundidad de la capa de mezcla, para cada región y época del año. Los coeficientes de correlación de estas regresiones también se probaron a un nivel de confianza de 95%.

La producción primaria se calculó usando el modelo de Platt et al. (1991) para comparar la PPint (mgC m-2 h-1) que resulta de los perfiles Gausianos de Chlz versus la que resulta de distribuciones homogéneas de Chlz. La expresión de Platt et al. (1991) se utilizó con la modificación sugerida por Giles-Guzmán y Álvarez-Borrego (2000) para aguas caso 1 (Chlz < 1.5 mg m-3). Esta modificación corrige la pendiente inicial (α*PAR) de la relación fotosíntesis-irradiancia (curvas P-E) de acuerdo a la distribución espectral in situ de la irradiancia escalar (PARz). En lugar de α*PAR en la expresión de Platt et al. (1991) se utilizó 43.2maxā *ph(z,Chl) (Giles-Guzmán y Álvarez-Borrego 2000), donde max es la eficiencia fotosintética cuántica máxima a baja irradiancia (mol C (mol cuanta)-1). La media del coeficiente específico de absorción del fitoplancton es ā *ph(z,Chl) (m2 mgChl-1), ponderada por la distribución espectral in situ de PARz y calculada de acuerdo con Giles-Guzmán y Álvarez-Borrego (2000). El factor 43.2 convierte moles de C a mgC, segundos a horas, y moles de cuanta a emoles de cuanta. Los cálculos se realizaron con el promedio de los parámetros fotosintéticos reportados por Malone (1971) para aguas del Caribe y del Atlántico aledañas a Cuba. Se utilizó una irradiancia escalar superficial constante (PARo = 1000 μmol cuanta m-2 s-1) de tal manera que las diferencias de PPint se pudiesen atribuir sólo a cambios en los perfiles de Chlz.
Resultados
La región de aguas profundas tuvo la Ts media más baja y la más alta: 23.5°C para la época fría y 28.3°C para la caliente (tabla 2). No hubo diferencia significativa entre las medias de Ts de las regiones Mississippi y Yucatán. Tampoco hubo diferencias significativas entre las medias de Chls de las regiones Mississippi y Yucatán (0.28 y 0.23 mg m-3, respectivamente), mientras que las de la región II fueron significativamente menores (0.12 mg m-3 para IIA y 0.08 mg m-3 para IIB) (tabla 2). La media de Chls para la época caliente de la región de aguas profundas fue significativamente menor que su valor correspondiente para la época fría, aunque la diferencia fue marginal. Las medias de la concentración de clorofila en el máximo profundo (Chlm) fueron significativamente más bajas para ambos periodos de la región de aguas profundas que las de las otras dos regiones. La media más alta de Chlm fue la de la región Yucatán (0.93 mg m-3) (tabla 2).
La integral de Chlz bajo la curva, h, fue significativamente más alta para la región Yucatán (46 mg m-2) que para la región Mississippi (38.5 mg m-2) y la época fría de la región de aguas profundas (42 mg m-2). El valor más bajo fue el de la época caliente de la región de aguas profundas (28.5 mg m-2) (tabla 2) . Todas las regiones y épocas tuvieron esencialmente el mismo promedio de Chlo (~0.05 mg m-3). La media de Chlo de la región Yucatán fue relativamente alta (0.12 mg m-3) pero su error estándar fue grande (0.05 mg m-3). Los promedios de σ para la regiones Mississippi y Yucatán no fueron significativamente diferentes entre sí, y fueron más bajos que el de la región de aguas profundas (~41 m, tabla 2). La profundidad media del DCM, Zm, tuvo valores >35 m, hasta de ~79 m para la región de aguas profundas (tabla 2).
La media mayor de MLD fue la de la región Mississippi (107 m) y la menor fue la de la época caliente de la región de aguas profundas (34 m) (tabla 2). Como era de esperarse, los valores de las medias de s para la región de aguas profundas, 84 J m-3 para la época fría y 119 J m-3 para la época caliente, fueron significativamente mayores que los de las regiones Yucatán y Mississippi (~45 J m-3) (tabla 2).
Los modelos de regresión lineal simple de los parámetros Gausianos en función de Chls, Ts, MLD, ó s no mostraron coeficientes de correlación significativos, con excepción de los de Zm. La profundidad del máximo de clorofila, Zm, tuvo correlación significativa únicamente con MLD y con s (tabla 3, fig. 3) . En algunos casos, las gráficas de los parámetros Gausianos versus una de las variables independientes sugirió una relación lineal, y en otros casos mostraron una relación exponencial (no se ilustra). La regresión del ln de los parámetros versus el ln de las variables independientes no resultó en coeficientes de correlación significativos en los casos de Chlo, h, y σ; y en el caso de Zm no produjo resultados mejores que los modelos lineales. El agregarle una segunda variable independiente a los modelos no contribuyó significativamente a mejorar el coeficiente de correlación (r). Por lo tanto, para calcular perfiles representativos de Chlz se propone el utilizar las medias de Chlo, h, y σ, para cada región y época (tabla 2).

La profundidad del DCM, Zm, cambia directamente con s e inversamente con MLD, de tal manera que los mayores valores de s corresponden a los mayores valores de Zm, y viceversa (entre más estratificada la columna de agua más profundo el DCM) (fig. 3). Para el propósito de estimar perfiles de Chlz, se tienen tres alternativas para calcular Zm. Una es usar las ecuaciones de regresión de la tabla 3 para calcular Zm en función de MLD ó s. Desafortunadamente esto no es práctico, porque las variables que se miden con sensores remotos, como la Ts, no proveen estimaciones de MLD ó s. Estos parámetros tienen baja correlación con Ts. Está fuera del alcance de esta contribución proporcionar una descripción detallada de la variación espacial y estacional de MLD y s, así como desarrollar algoritmos para calcularlos para el Golfo de México a partir de variables como Ts. La segunda alternativa es usar la media de Zm para cada región y época, pero esto no deja grados de libertad para usar imágenes de satélite. Por lo tanto, la tercera alternativa para estimar Zm es usar la ecuación 1 y el promedio de la concentración de clorofila de la imagen compuesta de satélite (Chlsat), para esa región y época de un año en particular. Como una primera aproximación se puede tomar la media de Chls igual a la media de Chlsat. Por lo tanto:

Los valores medios de Zm derivados de la ecuación 2 son prácticamente iguales a los derivados directamente de los perfiles de clorofila. Un solo valor de Chls produce diferentes perfiles de Chlz para las diferentes regiones y épocas. Como ejemplos se calcularon los perfiles con Chls iguales a 0.08 y 0.12 mg m-3 para la región de aguas profundas, e iguales a 0.20 y 0.30 mg m-3 para las regiones Yucatán y Mississippi (fig. 4). En todos los casos hubo un cambio claro de la forma del perfil dependiendo de la región y época.
Para el propósito de comparar la PPint resultante de los perfiles Gausianos con las correspondientes a las distribuciones homogéneas, se usó el procedimiento propuesto por Giles-Guzmán y Álvarez-Borrego (2000) para estimar el coeficiente de atenuación vertical de luz difusa. Como ejemplos, la clorofila integrada (Chlint, mg m-2) y PPint se calcularon para los perfiles de la figura 4 y para las distribuciones homogéneas correspondientes (éstas tuvieron una Chlz constante igual a Chls). Los valores de Chlint y PPint de los perfiles Gausianos fueron más altos que los obtenidos con las distribuciones homogéneas (tabla 4). El suponer un perfil de biomasa fitoplanctónica homogéneo resultó en una subestimación de PPint hasta en 43% y 12% con respecto a lo calculado con los perfiles no homogéneos, para los ejemplos con la más baja y la más alta Chls, respectivamente. La diferencia de PPint calculada con los perfiles de biomasa homogéneos y no homogéneos fue mayor para los ejemplos con las Chls bajas que para los de Chls altas (tabla 4).

Se pueden utilizar los resultados de la tabla 4 para analizar si los perfiles Gausianos de biomasa, derivados para diferentes regiones y épocas, con el mismo valor de Chls, producen valores diferentes de PPint. La PPint presenta casi los mismos valores para las regiones Yucatán y Mississippi con Chls iguales a 0.20 y 0.30 mg m-3 (55.4 versus 55.9, y 60.8 versus 60.1 mgC m-2 h-1, respectivamente). Con la misma Chls la región Yucatán tiene valores mayores de Chlz que los de la región Mississippi, con la diferencia aumentando con la profundidad (fig. 4). Esto se debe al valor más alto de h para la región I que para la región III (tabla 2). Sin embargo, la región III tiene una zona eufótica más profunda que la región I, de tal manera que la integración resulta en valores prácticamente iguales de PPint. Por otra parte, con Chls iguales a 0.08 y 0.12 mg m-3, las diferencias de PPint entre las dos épocas de la región de aguas profundas fueron 7% y 5%, respectivamente (con los valores más altos de PPint para la época fría), las cuales tampoco son significativas. Cuando se comparan valores de PPint, estimados con la misma Chls, para las regiones I ó III con los de la región II, se obtienen diferencias significativas >12% (no se ilustra).
Discusión
Cuando los anillos anticiclónicos que se forman de la Corriente de Lazo chocan con la costa occidental forman triadas (ciclón-anticiclón-ciclón) paralelas a la costa (Vidal et al. 1994). Entre el ciclón al norte y el anticiclón se forma una corriente que transporta agua de la plataforma continental mar adentro. Estas corrientes han sido detectadas con imágenes de satélite de color como lengüetas de Chlsat relativamente alta que se extienden de decenas a centenas de kilómetros del límite de la plataforma continental mar adentro (Biggs y Müller-Karger 1994). Los datos in situ disponibles de Chlz son escasos y no permitieron detectar estos fenómenos. Por lo tanto, los resultados de las medias de Chls y Chlm que se presentan aquí no los incluyen. Los valores de las medias de Chls y Chlm de las regiones Mississippi y Yucatán, más altos que los de la región de aguas profundas, se deben a las surgencias y también, en el caso de la región III, al aporte de nutrientes de los ríos, principalmente del Mississippi. A lo largo de Florida (al noreste del golfo) y en el Banco de Campeche la interacción por fricción entre la corriente y el fondo parece ser importante (Hsueh y O'Brien 1971), y se produce surgencia que acarrea agua más densa hasta cerca de la superficie reduciendo la estratificación (Wiseman Jr. y Sturges 1999) y aumentando la biomasa fitoplanctónica. La plataforma de Luisiana y Texas tiene una circulación ciclónica, excepto en verano cuando el flujo es hacia el este (Zavala-Hidalgo et al. 2003). Esta circulación ciclónica aporta aguas ricas en nutrientes a la superficie. De acuerdo con estos últimos autores, la circulación sobre la parte occidental del Banco de Campeche es hacia el sur, frente a la costa occidental de la Península de Yucatán, a lo largo de todo el año. Cuando el flujo costero converge frente al sur de Texas y frente al sur de Veracruz hay un flujo mar adentro que se puede detectar en imágenes de satélite como lengüetas de alta Chlsat. La divergencia asociada con la Corriente de Lazo puede ser un factor que produzca surgencia en el Banco de Campeche (frente a la Península de Yucatán), y frente a la punta suroeste de Florida, y esta divergencia ocurre a lo largo de la mayor parte del año (Lohrenz et al. 1999).
Para el propósito de definir regiones para aplicar modelos de producción primaria a datos de satélite de color del océano, es mejor que estas regiones sean lo más grande posible. Sathyendranath et al. (1995) indicaron que la mayoría de las fronteras entre regiones biogeoquímicas tienen diferencias significativas de Zm. Tal es el caso de las regiones del Golfo de México que se han definido aquí, como se muestra en la tabla 2. La profundidad del DCM define claramente tres regiones en el Golfo de México, dos regiones costeras en los extremos norte y sur, con valores medios de Zm similares y relativamente bajos (Mississippi y Yucatán), y la región de aguas profundas con valores mayores de Zm (74 y 79 m). La profundidad del DCM para la región de aguas profundas (IIA y IIB) fue mucho mayor que la profundidad de la capa de mezcla (MLD), como ha sido reportado por Barreiro-Güemes et al. (2003). Sathyendranath et al. (1995) reportaron una media de Zm para la región del Atlántico adyacente al Golfo de México igual a 84 m, la cual no es significativamente diferente de la de la región IIB. Los valores pequeños de Zm indican aguas superficiales ricas en nutrientes, y viceversa. Millán-Núñez et al. (1997) reportaron valores medios de Zm < 20 m para la subregión del norte del Sistema de la Corriente de California, cercana a la costa, e Hidalgo-González y Álvarez-Borrego (2001) reportaron valores medios de Zm< 15 m para la época fría de las partes central y norte del Golfo de California. Los valores medios de Zm del Golfo de México son similares a los de las subregiones oceánicas del Sistema de la Corriente de California frente a Baja California (45 to 85 m) (Millán-Núñez et al. 1997) y a los de la entrada al Golfo de California (28 to 37 m) (Hidalgo-González and Álvarez-Borrego 2001).
Los promedios de h para el Golfo de México son similares a los reportados por Millán-Núñez et al. (1997) para las subregiones alejadas de la costa del Sistema de la Corriente de California, y también son similares a los reportados por Hidalgo-González y Álvarez-Borrego (2001) para la época fría de la región de la entrada al Golfo de California. Los valores del parámetro h son más altos que la Chlint para la zona eufótica porque los valores de h incluyen toda el área bajo la curva Gausiana.
Con más de 3000 perfiles de Chlz Millán-Núñez et al. (1997) no pudieron construir modelos para estimar a para la mayoría de las regiones del Sistema de la Corriente de California; y en algunos casos no pudieron construir modelos para Chlo. Sathyendranath et al. (1995) concluyeron que es impráctico calcular σ a partir de Chls, o de otras variables. Millán-Núñez et al. (1997) y Barocio-León et al. (2007) indican que en los casos en que las regresiones no fuesen significativas, se deben utilizar las medias de los parámetros de las regiones y épocas correspondientes. Debido a lo escaso de los datos del Golfo de México no fue posible encontrar correlaciones significativas entre los parámetros Gausianos y las propiedades superficiales que pueden ser derivadas de datos de sensores en satélites. Por lo tanto, se propone utilizar las medias de Chlo, h, y σ, para cada región y época (tabla 2), para calcular perfiles representativos de Chlz.
Las ecuaciones de la tabla 3 muestran una relación muy clara entre la estructura física de la columna de agua y los perfiles de Chlz del Golfo de México, de una manera similar a como se mostró para el Golfo de California por Hidalgo-González y Álvarez-Borrego (2001). Sin embargo, los parámetros de las ecuaciones de la tabla 3 son significativamente diferentes a los de las ecuaciones del Golfo de California.
Los valores de Chlint y PPint de los perfiles Gausianos fueron más altos que los obtenidos con las distribuciones homogéneas (tabla 4). Sathyendranath et al. (1995) encontraron que en la escala de provincias individuales, la producción con Chlz no uniforme siempre resultó igual o mayor que la de la uniforme. En nuestro caso la diferencia de PPint calculada con los perfiles de biomasa homogéneos y no homogéneos fue mayor para los ejemplos con las Chls bajas que para los de Chls altas (tabla 4). Millán-Núñez et al. (1997) e Hidalgo-González y Álvarez-Borrego (2001) reportaron resultados similares para el Sistema de la Corriente de California y para el Golfo de California, respectivamente. Por otro lado, para el propósito de estimar promedios de PPint para las regiones I y III se pueden utilizar los mismos parámetros Gausianos; y también para estimar PPint para la región II se puede utilizar un solo conjunto de parámetros Gausianos para las dos épocas. Sin embargo, con la misma Chls, Zm y la concentración de clorofila en el DCM (Chlm) muestran cambios grandes entre regiones y épocas (fig. 4) y esto puede ser importante para estudios de pesquerías. Se ha sugerido que el DCM provee la abundancia necesaria de dinoflagelados para la supervivencia de larvas de peces pelágicos que comienzan a alimentarse en el Sistema de la Corriente de California (Lasker 1975).
Una limitación importante de este estudio es que la mayoría de los datos son discretos, basados en muestras de botellas. La fluorometría in vivo de flujo continuo hizo posible la observación de que el fitoplancton se puede concentrar en capas que a menudo no se muestrean con esquemas tradicionales con botellas (Strickland 1968). Gieskes et al. (1978) reportaron movimientos verticales de estas capas. Desafortunadamente, hay sólo algunos pocos perfiles continuos de Chlz disponibles y sólo para la región de aguas profundas (Barreiro-Güemes et al. 2003, Salas-De-León et al. 2004, Signoret et al. 2006). Actualmente, el uso de datos de Chlz de HPLC produce estimaciones mejores que las obtenidas por fluorometría. Sin embargo, Trees et al. (2000) reportaron que hay una relación lineal fuerte entre el logaritmo de los pigmentos accesorios y el logaritmo de Chlz, y esto en gran parte explica el éxito de los datos de Chlsat obtenidos por sensores remotos, aun cuando las poblaciones de fitoplancton pueden variar en su composición y en sus contenidos de pigmentos.
En conclusión, la capacidad de asociar perfiles no homogéneos de Chlz a datos de sensores de color en satélites nos permite mejorar las estimaciones de producción primaria. El suponer un perfil homogéneo de biomasa para el Golfo de México puede resultar en una subestimación significativa de PPint. En el Golfo de México, como en el Golfo de California (Hidalgo-González y Álvarez-Borrego 2001), la distribución vertical de la biomasa fitoplanctónica está fuertemente correlacionada con la estructura de la columna de agua. La división del Golfo de México en regiones y épocas resulta en diferencias significativas de Zm y de concentración de clorofila en el DCM, lo cual puede ser de importancia para estudios de pesquerías. Sin embargo, para estimar la PPint se requiere un solo conjunto de parámetros Gausianos para las regiones I y III, y otro para las dos épocas de la región II.
Agradecimientos
JM Domínguez y F Ponce realizaron la versión electrónica de las figuras. Este trabajo fue financiado parcialmente por el Consejo Nacional de Ciencia y Tecnología como parte del proyecto "Fuentes y sumideros de carbono en las márgenes continentales del Pacífico Mexicano" (clave SEP 2004/45813).
Referencias
Barocio-León OA, Millán-Núñez R, Santamaría-del-Ángel E, González-Silvera A. (2007) Phytoplankton primary productivity in the euphotic zone of the California Current System estimated from CZCS imagery. Cienc. Mar. 33: 59-72. [ Links ]
Barreiro-Güemes MT, Signoret-Poillon M, Bulit-Gámez C. 2003. Patrones de distribución de biomasa y producción primaria fitoplánctica en el Golfo de México. In: Barreiro-Güemes MT, Meave-Del-Castillo ME, Signoret-Poillon M, Figueroa-Torres MG (eds.), Planctología Mexicana. Sociedad Mexicana de Planctología, México, pp. 125-142. [ Links ]
Behrenfeld MJ, Falkowski PG. 1997. A consumer's guide to phytoplankton primary productivity models. Limnol. Oceanogr. 42: 1479-1491. [ Links ]
Biggs DC, Müller-Karger FE. 1994. Ship and satellite observations of chlorophyll stocks in interacting cyclone-anticyclone eddy pairs in the western Gulf of Mexico. J. Geophys. Res. 99: 7371-7384. [ Links ]
Candela J, Sheinbaum J, Ochoa J, Badan A, Leben R. 2002. The potential vorticity flux through the Yucatan Channel and the Loop Current in the Gulf of Mexico. Geophys. Res. Lett. 29: 2059, doi:10.1029/2002GL015587. [ Links ]
Cullen JJ, Eppley RW. 1981. Chlorophyll maximum layers of the Southern California Bight and possible mechanisms of their formation and maintenance. Oceanol. Acta 4: 23-32. [ Links ]
Dandonneau Y. 1979. Concentrations en chlorophylle dans le Pacifique tropical sud-ouest: Comparison avec d'autres aires océaniques tropicales. Oceanol. Acta 2: 133-142. [ Links ]
El-Sayed SZ, Turner JT. 1977. Productivity of the Antarctic and tropical/subtropical regions: A comparative study. In: Dunbar MJ (ed.), Polar Oceans. Arctic Institute of North America, Proc. of Conf., McGill Univ. Press, pp. 463-503. [ Links ]
Gieskes WWG, Kraay GW, Tijseen SB. 1978. Chlorophylls and their degradation products in the deep pigment maximum layer of the tropical North Atlantic. Neth. J. Sea Res. 12: 195-204. [ Links ]
Giles-Guzmán AD, Álvarez-Borrego S. 2000. Vertical attenuation coefficient of photosynthetically active radiation as a function of chlorophyll concentration and depth in case 1 waters. Appl. Opt. 39: 1351-1358. [ Links ]
Hernández-Becerril DU, Garcia-Resendiz JA, Salas-De-León DA, Monreal-Gómez MA, Signoret-Poillon M, Aldeco-Ramírez J. 2008. Nanoplankton fraction in the phytoplankton structure in the southern Gulf of Mexico (April 2000). Cienc. Mar. 34: 77-90. [ Links ]
Hidalgo-González RM, Álvarez-Borrego S. 2001. Chlorophyll profiles and the water column structure in the Gulf of California. Oceanol. Acta 24: 19-28. [ Links ]
Holm-Hansen O, Lorenzen CJ, Holmes RW, Strickland JDH. 1965. Fluorometric determination of chlorophyll. J. Cons. Perm. Int. Explor. Mer 30: 3-15. [ Links ]
Hsueh Y, O'Brien JJ. 1971. Steady coastal upwelling induced by an along-shore current. J. Phys. Oceanogr. 1: 180-186. [ Links ]
Hurlburt HE, Thompson JD. 1980. A numerical study of Loop Current intrusions and eddy shedding. J. Phys. Oceanogr. 10: 1611-1651. [ Links ]
Kirk JTO. 1994. Light and Photosynthesis in Aquatic Ecosystems. Cambridge Univ. Press, New York, 509 pp. [ Links ]
Lasker R. 1975. Field criteria for the survival of anchovy larvae: The relation between inshore chlorophyll maximum layers and successful first feeding. Fish. Bull. 73: 847-855. [ Links ]
Lewis MR, Cullen JJ, Platt T. 1983. Phytoplankton and thermal structure in the upper ocean: Consequences of nonuniformity in chlorophyll profile. J. Geophys. Res. 88: 2565-2570. [ Links ]
Lohrenz SE, Dagg MJ, Whitledge TE. 1990. Enhanced primary production at the plume/oceanic interface of the Mississippi River. Cont. Shelf Res. 10: 639-664. [ Links ]
Lohrenz SE, Wiesenburg DA, Arnone RA, Chen X. 1999. What controls primary production in the Gulf of Mexico? In: Kumpf H, Steidinger K, Sherman K (eds.), The Gulf of Mexico Large Marine Ecosystem. Blackwell Science, Malden, pp. 151-170. [ Links ]
Malone TC. 1971. The relative importance of nannoplankton and netplankton as primary producers in tropical oceanic and neritic phytoplankton communities. Limnol. Oceanogr. 16: 633-639. [ Links ]
Martínez-Sepúlveda M. 1994. Descripción de la profundidad de la capa mezclada superficial en el Golfo de California. Tesis de licenciatura, Universidad Autónoma de Baja California, Ensenada, México, 44 pp. [ Links ]
Matsumura S, Shiomoto A. 1993. Vertical distribution of primary productivity function Φ(II) for the estimation of primary productivity based on satellite remote sensing. Bull. Nat. Res. Inst. Far Seas Fish. 30: 227-270. [ Links ]
Millán-Núñez R, Álvarez-Borrego S, Trees CC. 1997. Modeling the vertical distribution of chlorophyll in the California Current System. J. Geophys. Res. 102: 8587-8595. [ Links ]
Monreal-Gómez MA, Salas-De-León DA. 1997. Circulación y estructura termohalina del Golfo de México. In: Lavín MF (ed.), Contribuciones a la Oceanografía Física en México, Monografía 3. Unión Geofísica Mexicana, Ensenada, pp. 183-199. [ Links ]
Morel A, Berthon JF. 1989. Surface pigments, algal biomass profiles, and potential production of the euphotic layer: Relationships reinvestigated in view of remote-sensing applications. Limnol. Oceanogr. 34: 1545-1562. [ Links ]
Müller-Karger FE, Walsh JJ, Evans RH, Meyers MB. 1991. On the seasonal phytoplankton concentration and sea surface temperature cycles of the Gulf of Mexico as determined by satellites. J. Geophys. Res. 96: 12645-12665. [ Links ]
Platt T, Sathyendranath S. 1993. Estimators of primary production for interpretation of remotely sensed data on ocean color. J. Geophys. Res. 98: 14561-14576. [ Links ]
Platt T, Sathyendranath S, Caverhill CM, Lewis MR. 1988. Ocean primary production and available light: Further algorithms for remote sensing. Deep-Sea Res. 35: 855-879. [ Links ]
Platt T, Caverhill CM, Sathyendranath S. 1991. Basin-scale estimates of oceanic primary production by remote sensing: The North Atlantic. J. Geophys. Res. 96: 15147-15159. [ Links ]
Salas-De-León DA, Monreal-Gómez MA, Signoret M, Aldeco J. 2004. Anticyclonic-cyclonic eddies and their impact on near-surface chlorophyll stocks and oxygen supersaturation over the Campeche Canyon, Gulf of Mexico. J. Geophys. Res. 109: CO5012, doi: 1029/2002JC001614. [ Links ]
Sánchez-Iturbe A, Flores-Coto C, Zavala-García F. 2006. Seasonal variation in growth and mortality of larval Syacium gunteri (Piscis: Paralichthyidae) in the southern Gulf of Mexico (1992 and 1993. Cienc. Mar. 32: 83-95. [ Links ]
Sathyendranath S, Longhurst A, Caverhill CM, Platt T. 1995. Regionally and seasonally differentiated primary production in the North Atlantic. Deep-Sea Res. 42: 1773-1802. [ Links ]
Signoret M, Javier A, Barreiro MT. 2006. Variabilidad de perfiles de clorofila a obtenidos por fluorescencia natural en el oeste y centro del Golfo de México en junio de 1999. Hidrobiológica 16: 221232. [ Links ]
Simpson JH, Hunter JR. 1974. Fronts in the Irish Sea. Nature 250: 404-406. [ Links ]
Stommel H. 1966. The Gulf Stream: A Physical and Dynamical Description. Univ. California Press, Berkeley, 189 pp. [ Links ]
Strickland JDH. 1968. A comparison of profiles of nutrient and chlorophyll concentrations taken from discrete depths and by continuous recording. Limnol. Oceanogr. 13: 388-391. [ Links ]
Trees CC, Clark DK, Bidigare RR, Ondrusek ME, Mueller JL. 2000. Accessory pigments versus chlorophyll a concentrations within the euphotic zone: A ubiquitous relationship. Limnol. Oceanogr. 45: 1130-1143. [ Links ]
Varela, RA, Cruzado A, Tintoré J, García-Landona E. 1992. Modeling the deep chlorophyll maximum: A coupled physical-biological approach. J. Mar. Res. 50: 441-463. [ Links ]
Vidal VMV, Vidal FV, Hernández AF, Meza E, Pérez-Molero JM. 1994. Baroclinic flows, transports, and kinematic properties in a cyclonic-anticyclonic-cyclonic ring triad in the Gulf of Mexico. J. Geophys. Res. 99: 7571-7597. [ Links ]
Wiseman Jr WJ, Sturges W. 1999. Physical oceanography of the Gulf of Mexico: Processes that regulate its biology. In: Kumpf H, Steidinger K, Sherman K (eds.), The Gulf of Mexico Large Marine Ecosystem. Blackwell Science, Malden, pp. 77-92. [ Links ]
Zavala-Hidalgo J, Morey SL, O'Brien JJ. 2003. Seasonal circulation on the western shelf of the Gulf of Mexico using a high-resolution numerical model. J. Geophys. Res. 108: C12, 3389, doi: 10.1029/ JC001879. [ Links ]


{kind=link}
{kind=link}