











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink1.Introduction
In the initial stage of photosynthesis, light is absorbed by pigment-protein complexes (PPC) and the electronic excitation energy is transfered to a reaction center (RC), where a charge separation reaction occurs 1.
Green sulfur bacteria is a photosynthetic organism whose light harvesting complex is composed of a chlorosome antenna anchored to aggregates of a protein called Fenna-Matthews-Olson complex (FMO) 2, 3. The FMO complex serves as a quantum wire that delivers electronic energy from the chlorosome to the RC with a quantum efficiency close to 100% 4.
The FMO complex is a trimeric protein of identical subunits which function independently of each other. Subunits of the FMO complex consist of two 𝛽 sheets that form a hydrophobic pocket which holds 7 strongly coupled pigment molecules, called chromophores or bacteriochlorophylls a (BChla) 2, 3.
Quantum coherence can be used as an operational resource to acomplish tasks that would not be possible by classical means 5-7. Ultrafast two-dimensional electronic spectroscopic experiments have revealed that during energy transport the FMO complex sustains electronic coherence between the seven chromophores. Coherence lasts for at least 300 fs at room temperature, long enough to impact energy transfer dynamics 8-10. In the FMO protein, the functional role of coherence manifests in an interplay between coherent dynamics of the system and incoherent effects of environment that assists quantum transport efficiency 4 11-14. Subsequent two-dimensional electronic spectroscopy experiments 15, 16 and a theoretical examination 17 demonstrate that the excitation coherently moves through two alternative energy transfer pathways depending on the site that is initially excited.
Quantum correlations between different parts of a system are a true signature of their non-classical nature. Quantum discord is a quantifier of quantum correlations that includes entanglement as a subset and is more robust than entanglement against environment influences 19 20. Quantum discord can be exploited as a useful resource to perform tasks in manners that are classically impossible like the implementation of quantum computation and quantum communication protocols 21-26.
Investigations in the energy transfer dynamics of the FMO complex have revealed the existence of bipartite and global entanglement between the electronic states of spatially separated chromophores 14, 27-31. These studies of excitation dynamics in the FMO protein show that entanglement lasts for 1-2 ps under Markovian models and for over 4 ps under more accurate non-Markovian models. According to the authors, the fact that the entanglement lifetime is comparable with the timescale of energy transfer in the FMO complex, a few picoseconds, suggests that quantum correlations may benefit excitation transport. Interestingly, quantum discord between pairs of chromophores lives for longer times than entanglement 29. A functional role for quantum correlations in pigment-protein complexes is consistent with coherence enhancing properties of energy transport like efficiency or robustness 4 11-14. We believe that the results of recent research where it is demonstrated that entanglement is maximum along the two electronic energy transfer pathways also support the idea of a functional role for quantum correlations 32-34. Three investigations have studied the role that non-classical correlations between chromophores play in energy transmission efficiency: two of them 35, 36 did not find any conclusive results and the third 37, with positive results, was subject to harsh criticism from the group who first quantified entanglement in the FMO protein 30.
The minimum quantum contribution to the uncertainty associated to the measurement of composite systems with local observables is known as local quantum uncertainty (LQU) . Recently, it has been proved that LQU meets the criteria for a discord-like quantifier of quantum correlations. The LQU approach has been used as a reliable measure to characterize nonclassical correlations in a few frameworks 34-44
The general motivation for studying coherence and quantum correlations in light-harvesting structures is that the precise characterization of these quantum resources is essential, on the one hand, for understanding their functional relation with the energy transfer mechanism and, on ther other, to be able to construct naturally robust quantum devices based on that mechanics. Our principal aim is to demonstrate that the Local Quantum Uncertainty is a highly flexible, robust, and consistent metric to characterize quantum correlations, which is and a novel approach that had not been studied before. We determine the Local Quantum Uncertainty in subsets of the chromophores network in a Gibbs thermal ensemble because a correct picture of which chromophores remain quantum correlated at long times depends on the thermal equilibrium state reached by the system.
Specifically, we implement the LQU method in subsystems of pure and mixed states represented by the eigenstates and by the thermal equilibrium state determined by the FMO Hamiltonian from Chlorobium tepidum. Remarkable advantages of the LQU approach discussed in this work over other methods are: LQU is a discord-like quantifier of quantum correlations so it is more general than the entanglement based techniques, LQU is easier to calculate than direct definition of quantum discord, the LQU approach can be readily implemented in dynamical systems, and the LQU method flexibility allows to describe the sets with the strongest non-classical associations in qubits networks. The identification of the strongest associations as subsystems of three sites serves to investigate whether it is possible to relate quantum correlations with energy transport pathways. Finally, we take the LQU of the most quantum correlated subsets of chromophores as the signature of the non-classicity of the system to study physical properties such as populations, energy fluctuations, and specific heat. This analysis shows that the availibility of energy levels determined by increasing temperature dictates the relation between a measurable macroscopic magnitude and non-classical resources.
This paper is organized as follows: Sec. 2 describes the quantum states that emerge from the excitonic Hamiltonian of the FMO complex; Sec. 3 describes the aspect of quantum coherence that defines it as a quantifiable resource to be consumed in quantum information protocols; Sec. 4 develops the LQU method of quantifying quantum correlations in a chromophores network; Sec. 5 shows results of LQU, concurrence and coherence in subsystems of pure and mixed states of the FMO complex; finally, in section 6 we present an overview of the implementation of the LQU approach: usefulness, advantages, limitations and scope.
2.Model for the FMO complex chromophores network
The FMO protein is a trimer constituted of three identical monomers which function independently. For this reason, we focused on electronic energy transfer (EET) in only one of the monomers. Each monomer has a 7 chromophores network embedded in its structure. In the network, wave functions of the chromophores are distant enough from each other to overlap.
The electronic state of chromophore m goes from its ground state
where the base vector
where site energies Em are the optical transition energies for each site and interaction terms V nm of energy transfer correspond to dipole-dipole electric interactions between pairs of site excitons.
Adolphs and Renger 45 calculated individual excitation energies Em running a genetic algorithm that takes site energies as parameters that are optimized to fit experimental linear absorption spectra and linear and circular dichroism; interaction terms V nm were calculated using structural data from the FMO protein crystal.
We find the delocalized exciton eigenstates
with 𝑀=1,...,7. Eigenstates
where
where the density matrix elements of the eigenstates are:
and the superindex M indicates the eigenstate they belong to.
If the system is in thermal equilibrium the mixed state can be written as:
where
are the density matrix elements when the state is spanned in terms of the site basis.
3.Coherence as a quantifiable resource
In quantum information science, coherence is used as a resource that is consumed to
perform tasks that are not possible by classical means. The resource theory for
coherence interprets physical laws as constraints on the set of free quantum
operations to be implemented on quantum states. If a coherent state is the resource
to be consumed then the set of free quantum operations are incoherent operations and
the available free states are incoherent states. A proper measure of coherence C
must fulfill the following conditions: C(ρ) = 0 when ρ is a incoherent state, C does
not increase under incoherent operations (monotonicity), C does not increase on
average under selective incoherent operations (strong monotonicity), C does not
increase under mixing of quantum states (convexity). The l1 norm of
coherence
fulfills the former conditions so we will use it as a proper measure of coherence.
For a d-dimensional Hilbert space with a reference basis
For the state
4.Quantum correlations in
The first proposals for measuring the non-classical share of correlations were based on the nonseparability of the subsystems states, i.e., quantum entanglement. However, entanglement does not account for all non-classical correlations because separable mixed states usually contain quantum correlations. Quantum discord is a more general quantifier of non-classical correlations than the entanglement based ones 48. Quantum discord is based on the idea that by measuring one of the subsystems one can extract the shared classical information. Except for a few simple cases quantum discord is hard to compute, nevertheless a seemingly disadvantageous feature of quantum mechanics (uncertainty) provides an easier method for calculating quantum discord.
There is a close link between quantum-induced uncertainty and quantum correlations that can be exploited to establish a discord-like quantifier of quantum correlations 38. In quantum systems the measurement uncertainty may have classical and quantum contributions. If the state to be measured is incoherent in the observable eigenbasis the measurement output statistics will be classical: measurement of a pure state is completely deterministic and it will exhibit zero variance but for mixed states the contribution to the variance comes from classical uncertainty due to the mixedness of the state. In states displaying coherence, the measurement uncertainty has two contributions, apart from the classical randomness that comes from the mixing, there is a quantum randomness that is intrinsic to coherence. This quantum randomness manifests in the interference pattern of the outcome statistics and is an additional quantum component to the uncertainty.
The quantum contribution to the uncertainty is due to the non-commutativity between
the state ρ and the observable K 39. A reliable measure of quantum uncertainty is zero if and
only if ρ and K commute (ρ is a eigenstate of K or a mixture of eigenstates of K),
does not increase under classical mixing and is equal to the variance for pure
states. The Wigner-Yanase skew information
fulfills the former conditions so we will use it as a convenient measure of quantum uncertainty.
We have identified the quantum share to the measurement uncertainty and quantified it by a measure of the non-commutativity between the state ρ and the observable K. Now we will proceed to establish the relationship between quantum uncertainty and quantum correlations. In a bipartite system only zero discord states:
can commute with local observables
where the minimization is over the set of local observables with non-degenerate spectrum. It can be proven that the LQU fulfills the requirements to be a discord-like quantifier of non-classical correlations in bipartite states 38, 39. There is a geometric interpretation of the LQU that suggests a relation with symmetric discord: the LQU is the minimum Hellinger distance between the state ρ before and after a local root-of-unity unitary operator has been applied 38.
To minimize the skew information, the local observable KA can be parametrized using the Pauli matrices and the identity operator:
where α, β are complex scalars,
The advantage of this approach lies in the existence of a closed form to calculate
the LQU for bipartite systems where A is a qubit and B a qudit of dimension d (ρ is
a state defined on an Hilbert space
where the matrix elements of
Equation (15) involves the maximization of a quadratic form which results in:
where
5.Results
We calculated quantum resources in pure and mixed states of the FMO protein. Pure
states are constituted by the eigenstates
To demonstrate the advantageous features of the LQU approach we will follow this method: (1) to define families of partitions in the system, (2) to determinate the quantum states and (3) calculate the LQU between subsystems. We choose three three ways to divide the seven chromophores network; each partition defines a family of quantum correlations.
When the system is described by a eigenstate
where the
To calculate LQU between subsystems we need to obtain the
Eq. (19) is the general formula that we will use to calculate the three families of quantum correlations in the eigenstates and in thermal equilibrium state.
5.1.Family
The first family f1 corresponds to the LQU between one chromophore
In Fig. 1, two histograms show the LQU
between Subsystem A
1 and Subsystem A
2 for the seven eigenstates
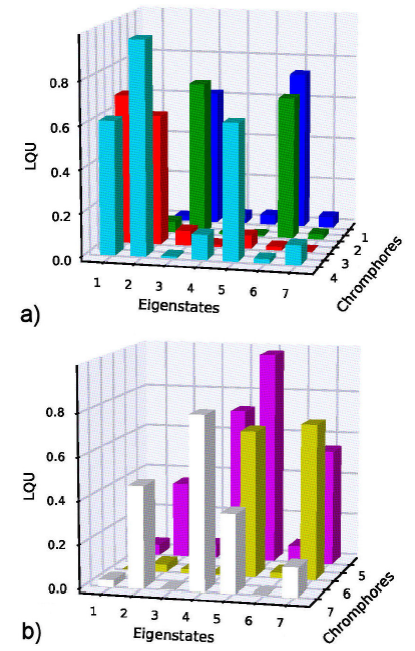
Figure 1 LQU between between one chromophore
The next step is to extend the quantum correlations analysis of pure states, represented by the eigenstates, to the state in thermal equilibrium. In Fig. 2 we show the LQU between individual chromophores Subsystem A 1 and the remaining 6 chromophores Subsystem B 1 as a function of temperature for the state in thermal equilibrium. In Fig. 2a) we show the LQU of chromophores C3 and C4 with the rest of the chromophores network. At low temperatures, chromophores C3 and C4 are the most quantum correlated with the rest of the network. This can be explained if we recall that the eigenstates with the greatest contribution to the LQU between C3 (or C4) and the rest of the network are the ground state and the first excited state. In Fig. 2b) we show the LQU of chromophores C1, C2, C5, C6, and C7 with the rest of the chromophores network. Regarding the associations of Fig. 2b), the values of LQU are marginal in amplitude and the curves show a local maximum around 100 K. Besides the difference in values of LQU, we separated the associations of family f1 in two sets because in Fig. 2a) the LQU is a strictly decreasing function while in Fig. 2b) the LQU first decreases, then increases and finally decreases again as the temperature rises. The LQU of the associations in Fig. 2b) takes low values at low energies because the eigenstates 1 and 2 do not have an important participation; as the temperature increases so does the LQU because eigenstates 3, 4, and 5 start to participate. At high temperatures, quantum correlations vanish in Gibbs states because all the eigenstates are equally probable, i.e., the mixedness is maximum and the non-classical correlations are lost.
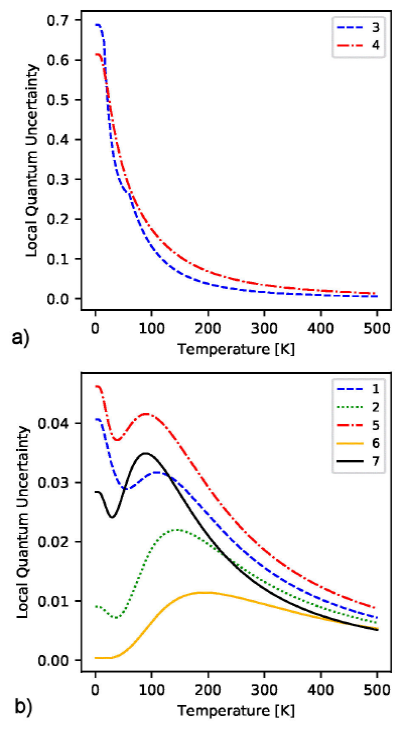
Figure 2 LQU between between one chromophore
To investigate the relation between two different quantum resources we calculated
the quantum coherence of the system and compared it with LQU. In the inset of
Fig. 3 we plot the quantum coherence

5.2.Family
The second family f2 accounts for the LQU between chromophore
We determine the quantum correlations defined by family f2 in
subsystems of two chromophores. We calculated the partial trace to the state of
the whole system
First, we examine the associations determined by family f2 in the
lowest eigenstates of the FMO Hamiltonian but in Fig. 4 we only present results from the ground state. For the lowest
eigenstates the highest correlation is between chromophores C3 and
C4. In decreasing order, correlations that also stand out when
the system is in the first excited state are:

After studying correlations in the eigenstates, we used the thermal state to establish reliability of the LQU approach by comparing it with concurrence (a valid quantifier of entanglement in the two qubit subspace). We calculate concurrence and LQU in all the combinations of pairs of chromophores but only show the pairs with the highest values.
Figure 5a) exhibits LQU curves for the
pairs
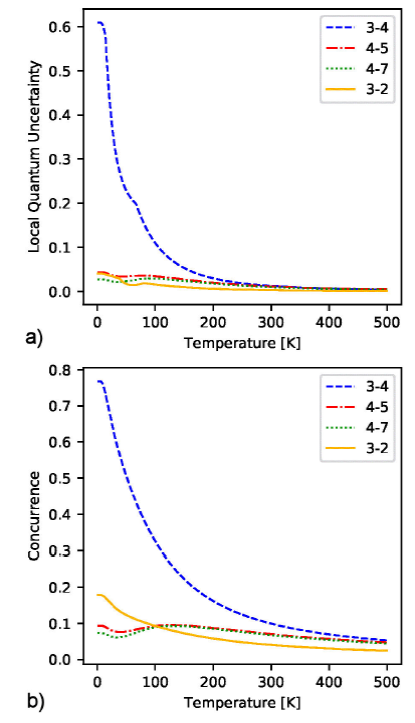
Figure 5.a) LQU and b) concurrence as a function of temperature for chromophore pairs{C3,C4}, {C4,C5},{C4,7} and {C3,C2} .
The rest of the pairs, with weaker contributions to the quantum correlations, also show a qualitative similarity between LQU and concurrence curves. These results confirm that the LQU approach is a reliable method for measuring quantum correlations in subsystems of the FMO complex.
A direct result for any pair of chromophores {Cr,Cs} with
quantum state
There is a direct form of calculating concurrence in
This is useful because the quantum state of two chromophores
{Cr,Cs} is a
where the elements
In the specific case of two site excitons, the elements
{Q12,Q41,Q44} are zero, therefore for any
pair of chromophores {Cr, Cs} with quantum state
With the matrix in Eq. (22) we can also express the coherence for the state of two chromophores {Cr,Cs}:
which is equal to the concurrence, as can be seen in Eq. (23).
5.3.Family
Now that we have evidence that LQU is a valid quantifier of quantum correlations
in subsystems of the FMO complex we search triads of chromophores whose
association results in stronger correlations than those found in bipartite
systems. To do so, we calculated LQU between one chromophore
We calculated the partial trace to the state of the whole system
We continue with the analysis in sets of three chromophores considering the eigenstates with lower energy but in Fig. 6 we only present an histogram with results from the ground state. Results show that for both the ground state the sets with the highest values of LQU are chromophore C4 with the pair {C3,C7}, chromophore C3 with the pair {C2,C4}, and chromophore C4 with the pair {C3,C5}.

After considering the eigenstates we will broaden the analysis to the thermal
state. In Fig. 7, we present the selected
triads where the LQU between one chromophore Subsystem A
3 and a pair of chromophores Subsystem B
3 is function of temperature a) and coherence b). Figure 7 shows the sets of chromophores with
the highest values of LQU (
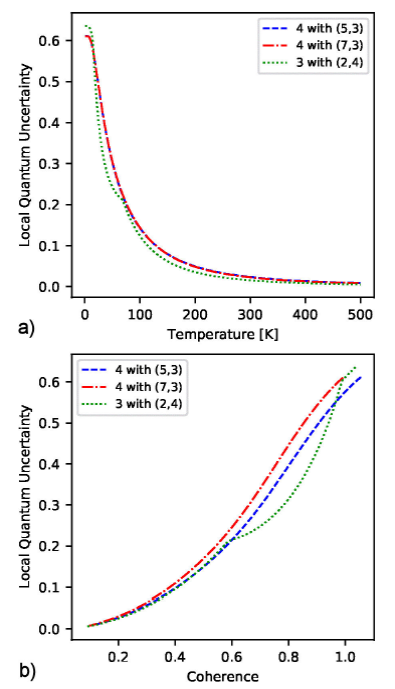
Figure 7 LQU between chromophores {Cr} and {Cs, Ct} for the thermal state as function of a) temperature and b) coherence.
With respect to the parametric curves of LQU versus coherence, results show that the triads of chromophores with the highest values of LQU, {C4,C3,C7}, {C3,C2,C4}, and {C4,C3,C5}, display a functional relation with their coherence. A monotonically increasing function was also present in the strongest associations of chromophores in families f1 and f2.
5.4.Quantum correlated subsets of chromophores in the physical framework of the FMO system in thermal equilibrium
The FMO protein mediates excitation energy transfer between the chlorosomes and
the reaction center. Chromophores 3 and 4 are the linker pigments between the
FMO complex and the RC. We use the thermodynamic equilibrium state in Eq. (7) to
calculate the functional dependence of the populations on the temperature, cf.
Fig. 8. At low temperatures and when
thermal equilibrium state has been reached, the highest probability is finding
the system in the localized excited state
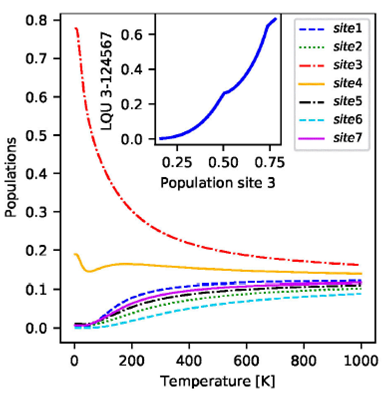
Figure 8 The main panel shows the populations of the seven sites as function of the temperature. The inset shows the LQU between C3 and the rest of the network as a function of the population of site 3.
We evaluate the relation between most correlated sets of chromophores and
population of site 3, the most probable of the linker pigments to be excited
when thermal equilibrium state has been reached. In the inset of Fig. 8 we show a parametric plot of the LQU
between site 3 and the rest of the network versus population of site 3, i.e.,
As an intermediate step, before searching for a relation between non-classicity
and a measurable macroscopic magnitude, we evaluate the fluctuation of the
energy
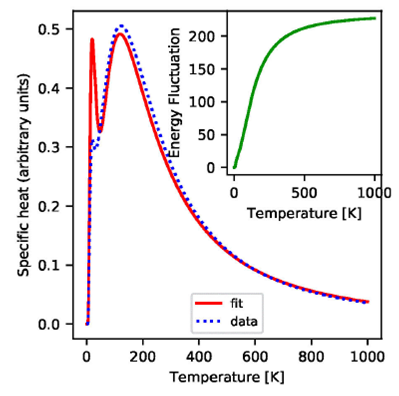
Figure 9 The main panel shows the specific heat at constant volume as
function of the temperature. The inset shows the energy fluctuation
of the system
With this result we can now evaluate the relation between the variance, that
accounts for both classical and quantum uncertainty, and the LQU, a quantifier
of the quantum contribution to the uncertainty. In the inset of Fig. 10, we show a parametric curve of the
fluctuation of the energy versus the LQU of chromophore C3 with the
rest of the network, i.e.,
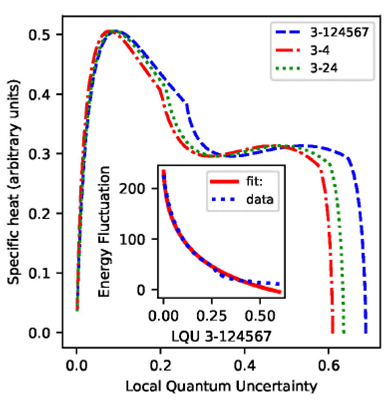
Figure 10 The main panel shows the specific heat at constant volume as
function of the LQU associated to the most correlated subsets of
chromophores, i.e. C3 with the rest of the network,
C3 with C4, and C3 with
{C2,C4}. The inset shows the energy
fluctuation of the system
To evaluate the relation between non-classicity and a measurable macroscopic
magnitude we begin by analyzing the functional dependence of the specific heat
at constant volume on the temperature
The system presents a Schottky-like anomaly in the specific heat data as a
function of temperature. The Schottky anomaly manifests itself as a maximum in
the specific heat when the energy gap
A multilevel system might presents multiple peaks in the specific heat that
correspond both to the discrete energy gaps
In a model for a multilevel system in thermal equilibrium with no degeneracies, the specific heat Cv is:
where
To investigate the relation between non-classicity and a measurable macroscopic
magnitude, in Fig. 10 we present a
parametric plot of specific heat versus the LQU of the strongest associations:
The peak in specific heat at the right, associated to the prevalence of the
energy levels 𝜀 0 and ε1, shows that as temperature rises from 0 to
∼ 50 K the LQU is constant, present the highest values, and the specific heat
sharply increases from 0 to 0.3. The eigenstate analysis shows that by accesing
to the lowest energy levels ε0 and ε1, the system presents
specific non-classical features with the highest values. Therefore, at low
temperatures, the fast increment in the capacity of the system to absorb heat
energy is related to the prevalence of specific non-classical correlations. As
the temperature increases and LQU decreases from 0.6 to ∼0.3 the specific heat
remains constant. The peak in specific heat at the left, associated to the
availibility of the energy gap
6.Conclusions
This article presents a systematic LQU based method intended to examine the structure of quantum correlations in pigment-protein complexes. To do so LQU, concurrence, and coherence were computed in subsystems of pure and mixed states of the FMO complex from green sulfur bacteria.
The LQU method consists in selecting families of partitions in the system, determination of the quantum state and calculation of LQU between subsystems. Partitions are selected to define two subsystems where one is a qubit and the other is a qudit (in the FMO complex the qubit corresponds to a single site and the qudit can be any number of sites). Determination of the quantum states is achieved by calculating the corresponding partial trace to the density matrix of the whole system. Calculation of LQU is accomplished by selecting the maximum eigenvalue of a 3 x 3 matrix. The LQU method input is a quantum state of site excitons therefore it can be readily implemented in dynamical models.
To show the versatility of the LQU method, we calculated quantum correlations between subsystems that had never been considered before, i.e., LQU between individual chromophores and the remaining 6 chromophores (family f1). Correlations from family 𝑓 1 present a global perspective of the chromophores individual participation. Results of family f1 show that chromophores C3 and C4 are the most quantum correlated with the rest of the network. In subsequent partitions, the robustness of the LQU allows us to find the specific associations of chromophores that sustain quantum correlations of chromophores C3 and C3.
To validate the LQU as a discord-like quantifier we compared it with a valid entanglement quantifier of two qubits (concurrence). The LQU-concurrence comparison was made using the thermal equilibrium state for all the combinations of two chromophores (family f2). According to results of family f2, quantum correlations of chromophores C3 and C4 with the rest of the network mainly arise from the following pairs of chromophores:{C3,C4},{ C4,C5},{C4.C7}, and {C2,C3}.
LQU between chromophores C3 and C4 is an order of magnitude higher than for any other pair but, as the relation between pairs suggests, by adding a specific third site the association strengthens even more. Associations suggested by family f2 are confirmed by results of family f3 which show that the most quantum correlated triads are: C3 with {C2,C4}, C4 with {C3,C5}, and C4 with {C3,C7}.
Three additional results were found: (1) By comparing the LQU approach with entanglement based techniques in pairs of chromophores we proved that the former is a valid discord-like quantifier of quantum correlations. (2) When we calculated quantum correlations using the eigenstates we found that some subsystems have high values of LQU in the ground and first excited state. When considering the state of these subsystems in thermal equilibrium we found that they have remarkably high LQU at low temperatures. This relation is present in the three families. (3) A monotonically increasing function between two quantum resources, coherence and quantum correlations, is present in the most correlated associations of families f1 and f3. In all associations of family f2 coherence equals concurrence.
One of the most interesting problems in quantum biology is finding out if quantum correlations in light harvesting complexes play a functional role enhancing energy transport efficiency. Investigations addressing this issue did not find any conclusive results 35, 36 or were subject to criticism 37. In our work, we quantified quantum resources in a system that remains in the same state as time elapses: the system is either in a stationary state or in thermal equilibrium. We did not work with a dynamic model so we cannot directly relate our results with excitation transport efficiency. Nevertheless, there is a previously reported relation between the most quantum correlated subsets of the chromophores network and the two energy transport pathways in the FMO protein (32)-(34).
In the FMO complex, the excitation coherently moves through two alternative energy
transfer pathways depending on the site that is initially excited . When chromophore
C1 is the initial excited pigment, the excitation propagates through
pathway A:
The relation between strong quantum correlations and energy transport pathways is consistent with previous research of multipartite entanglement in the FMO protein . In our work, we show that the LQU is a promising metric and provides a framework to establish the posible relation of quantum energy transmission with the non-classical correlations in the FMO complex.
The non-classicity of the system, represented by the LQU of the most quantum correlated subsets of chromophores, can be used to analyze physical properties such as populations, energy fluctuations, and specific heat. As the temperature rises, the number of accesible states is increased and the double peak structure that shows up in the specific heat reflects the availability of the energy gaps Δ1 and Δ2. The first peak, where the states ε0 and ε1 prevail, shows a correspondence between a sharp increase in heat capacity and constant LQU. The second peak, where the states ε0, ε1, and ε2 are available, the quantum signatures of the system are marginal and heat capacity begins to diminish. Thus, the accessibility of energy levels, mapped by the specific heat, defines the relation between a measurable macroscopic magnitude and non-classical resources.
The benefits of the LQU approach over other techniques are: (1) being LQU a discord-like quantifier it is more general than the entanglement based techniques, (2) LQU is easy to compute because the inherent minimization process associated with discord reduces to finding the maximum eigenvalue of a 3 x 3 matrix, (3) LQU permits characterization of correlations that had not been studied before (family f1 and f3), (4) the LQU method discloses the strongest associations in qubits networks and (5) the LQU technique can be readily implemented in dynamical systems. The principal limitation of the LQU approach is that it is a bipartite quantifier so it cannot offer a global measure of quantum correlations.
We introduced a quantum information method based on local quantum uncertainty to comprehensively characterize non-classical correlations in subsystems of the FMO complex. Identification of the most quantum correlated subsystems reveals a detailed map of the exceptionally efficient structure of the FMO complex. The study of biological networks has potential applications in engineering and quantum information technology.

