











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCTION
In Mexican horticulture, tomato (Solanum lycopersicum L.) is a primordial crop, it being one of the most consumed vegetables per capita worldwide, as well as one of the most widely grown in greenhouses, both in Mexico and in Europe and the United States of America. The greenhouse tomato production system consists of the use of varieties with an undetermined development type (Resh, 1992; Sánchez-del Castillo et al., 2009). Mexico produces over two million tons, with an estimated value of 15 billion pesos, in a surface area of 50 thousand h (SIAP, 2016).
However, this crop is attacked by pests that cause severe damages, one of them being Bactericera cockerelli (Liu & Trumble, 2004). This pest causes toxin type direct and indirect damages (Garzón-Tiznado et al., 2005) and has caused losses in the national production of this vegetable crop (Bujanos et al., 2005). In Mexico, B. cockerelli is one of the most critical pests in tomato, pepper, and potato crops (Vega-Gutiérrez et al., 2008). Since it is a vector of tomato diseases, B. cockerelli has an excellent capacity to increase its population density, as the female can deposit up to 1,400 eggs during its lifecycle (Liu & Trumble, 2006).
Because of this, high densities of this insect can cause significant damages to tomato crops. Liu et al. (2006) and Servín et al. (2008) mention than among the most relevant problems caused by this insect is its capacity to inject a toxin that causes modifications in the typical coloring of the leaves, turning them yellow or purple. This symptom is known as “psyllid yellows” or “potato purple tip,” which causes plant malformation and death. The symptoms in tomato plants include growth stop, the proliferation of new buds, chlorosis, purpling of the leaves, stimulated flowering, and overproduction of small, low-quality fruits. Given this problem, the present work consisted of determining the life history of B. cockerelli instars in the function of two tomato varieties, mean temperatures, and accumulated heat units.
MATERIALS AND METHODS
Study Area. Research experiments were carried out from October to December 2016 and, from January to March 2017, in the greenhouse of the Institute of Plant Health of the Colegio de Postgraduados, Campus Montecillo, Texcoco, State of Mexico.
The colony of Bactericera cockerelli. It sowed the seeds of tomato variety Charanda "F1" and "Rafaello" in germination trays for the establishment of the breeding of B. cockerelli. In Mexico, these varieties are the most cultivated by presenting a long period of fructification and post-harvest (Tigchelaar, 1986; Mendoza-De Jesus et al., 2010). Subsequently, 31 days after sowing (DDS) selected and transplanted, in a proper manner, 20 plants of each variety in polyethylene bags with two kilograms of the substrate (50% of Canada Growing Mix 1VM and 50% of tezontle). Later, the plants placed in the interior of entomological boxes (62 cm x 95 cm and 95 cm) lined with mesh organza. Right away, were put fourth and fifth instar nymphs and raised to B. cockerelli by seven generations to 14:10 h of light-dark and ambient temperature. The nymphs used in the breeding were collected previously in tomato plants of the "Rio Grande" variety in the greenhouse of Biological Control of the Colegio de Postgraduados. During reproduction, 300 ml of the nutrient solution was applied to each plant per day. The nutrient solution consisted of 500 ml sulfuric acid, ferrous sulfate 75 g, 1,200 ml phosphoric acid, 8.4 kg calcium nitrate, 3.5 kg potassium nitrate, 4.5 kg magnesium sulfate, 3 kg potassium sulfate, 40 g manganese sulfate, 5 g zinc sulfate, 5 g copper sulfate, 50 g borax for 10,000 liters of water.
Establishment of the experiments. The experiments developed in entomological boxes, previously described, and in tomato plants of the variety "Charanda F1" and "Rafaello" with 31 days. For each variety of tomato, there were three entomological boxes with five floors and 65 adults of B. cockerelli. The plants were placed in the entomological boxes. Immediately, the adults set in the plants for 24 h. Adults were removed and counted; the number of eggs present in the leaves were adjusted to 100. Subsequently, every 24 h were the nymphal stages up to the adult phase were counted. Also, during the process of the experiment, the temperature and humidity was recorded every 30 minutes using an electronic sensor (HOBO® model H-08-032-08 # 333699), temperatures ranged from 2°C to 40°C. The substrate, nutrient solution, and care of the tomato plants were similar to the one used in the development of the breeding of B. cockerelli.
Statistical analysis. We used the number of individuals registered to calculate the survival curves (n x ) of the stages of B. cockerelli in 2016 and 2017. The other hand analyzed the behavior of insects and the effect of temperature on the longevity of the population. To measure both parameters, we have taken into account the following factors: stages of B. cockerelli: eggs, nymph one, two, three, four, five, and adult. Like so tomato varieties: Charanda F1 (CF1) and Rafaello (RAF); crop cycles: C1 and C2, and the average temperature in cycle one and cycle two: temPC1 and temPC2 respectively. The statistical analysis was a development out with a complete block design in the statistical software (SAS Institute, 2005). The significance of the factors considered were tested through the application of the procedure GLIMMIX with an alpha of 0.05. To investigate the effect of the four factors in B. cockerelli, we performed a one-way, two-way, and three-way analysis of variance. Tukey’s method used for multiple comparisons and contrasts for main effects and interactions. The statistical analysis carried out in the SAS statistical software (SAS Institute, 2005). The GLM procedure used to generate the analysis of variance table and to estimate and to test contrast.
Finally, we analyzed the behavior of the psyllid through heat units per cycle and tomato varieties using the method of average temperature (Herms, 2004; Sridhar & Reddy, 2013). The parameter evaluated was, the unit's heat of the nymphal stages of B. cockerelli, and were calculated using the following formula:
Where: HU = Heat Units
Moreover, 7ºC corresponds to the minimum temperature threshold of development of B. cockerelli (González-Maldonado & García-Gutiérrez, 2012).
The development of the psyllid through heat units was analyzed, first between the two crop cycles and then comparing between the two tomato varieties. The t-test in the INFOSTAT statistical software was used (UNC, 2013).
RESULTS
In the first crop cycle, the B. cockerelli populations, it was noted to have a shorter lifespan in the CF1 in comparison with the populations present in the RAF. In the RAF, higher mortality in the development of nymphs one and three, the average mortality of eggs, and lower mortality of adults were observed (Fig. 1). Nevertheless, in the second crop cycle, in the number one entomological box of CF1, high mortality was obtained in the nymph five and adult. Contrary to what obtained inside the RAF growth chamber number 2, where the results revealed that all stages of the insect suffered from higher mortality compared to the remaining chambers (Fig. 1).
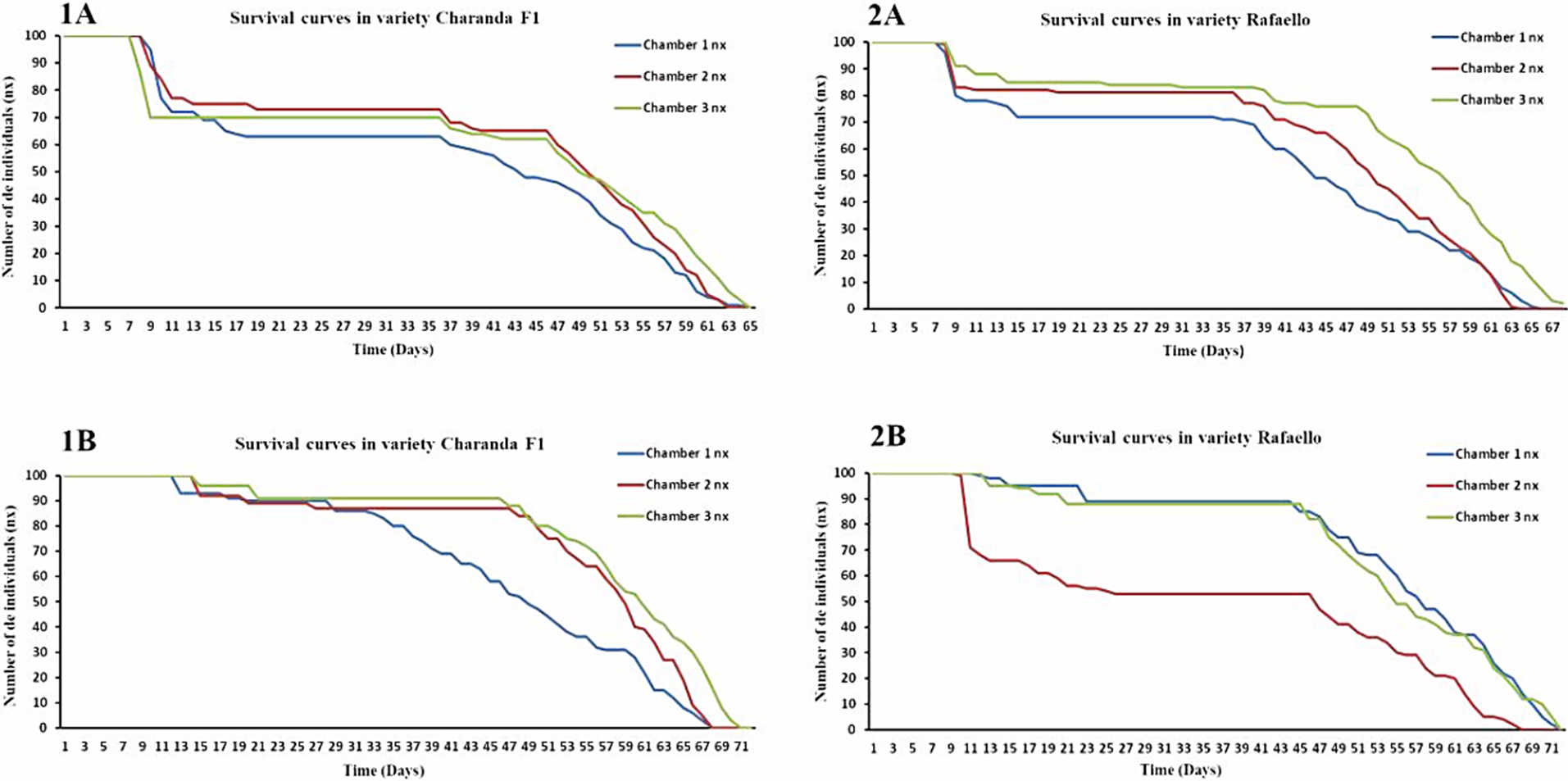
Figure 1 Survival curves (nx) of B. cockerelli in the tomato varieties Charanda F1 (1A and 1B) and Rafaello (2A and 2B) grown in the greenhouse during October-December 2016 (1A and 2A) - January-March 2017 (1B and 2B).
In the development analysis of B. cockerelli instars, the factors: cycle, variety, and instar, as well as the interactions: cycle-variety and cycle-instar, explained the biological cycle of the insect. In the mentioned factors, the biological cycle of the insects in C1 observed to be significantly different from that of C2 (Table 1). About temperature per cycle, Table 2 shows that insect survival was higher in C2 than in C1.
Table 1 Three-way of variance table for B. cockerelli in the greenhouse (October-December 2016 and January-March 2017).
| Source of variation | Degrees of freedom | Sum of the squares | Mean square | F-value | P-value |
|---|---|---|---|---|---|
| Cycle | 1 | 1226.67 | 1226.6 | 26.61 | <0.0001 |
| Variety | 1 | 192.01 | 192 | 4.17 | 0.0451 |
| Instar | 6 | 5930.61 | 988.4 | 21.44 | <0.0001 |
| Cycle*Variety | 1 | 520.01 | 520 | 11.28 | <0.0013 |
| Cycle*Instar | 6 | 619.90 | 103.3 | 2.24 | 0.0494 |
Table 2 Means comparison of temperatures of both cycles for the behavior of B. cockerelli grown in two tomato varieties in the greenhouse (October-December 2016 and January-March 2017).
| temPC | No. of observations | Means | P-value |
|---|---|---|---|
| TemPC1 | 42 | 21.30a | <0.0001 |
| temPC2 | 42 | 19.38b | <0.0001 |
tempPC1 = mean temperature in Cycle 1, temPC2= mean temperature in Cycle 2, means with the same letter are statistically equal, Tukey (P < 0.05).
Table 3 shows the temPC1-CF1 interaction significantly affects the longevity of the insect. The lifespan of the insect was less than 21ºC, while, as temperature decreased (19ºC), the insects increased its lifespan in the same variety. Contrastingly, this behavior not observed in the RAF variety.
Table 3 Means comparison of the temPC*variety interaction of the biological cycle of B. cockerelli grown in two tomato varieties in the greenhouse (October-December 2016 and January-March 2017).
| Interactions | No. of observations | Means | T-value | P-value | Mean temperature (°C) |
|---|---|---|---|---|---|
| temPC2*Charanda F1 | 21 | 89.9a | 60.68 | <0.0001 | 19.38 |
| temPC2*Rafaello | 21 | 87.9a | 59.36 | <0.0001 | 19.38 |
| temPC1*Rafaello | 21 | 85.2a | 57.56 | <0.0001 | 21.30 |
| temPC1*Charanda F1 | 21 | 77.2b | 52.16 | <0.0001 | 21.30 |
temPC1 = mean temperature in Cycle 1, temPC2 = mean temperature in Cycle 2, means with the same letter are statistically equal, Tukey (P < 0.05).
On the other hand, the C1-CF1 interaction significantly affects the longevity of the insect population compared against the other interactions in both cycles (Table 4). The insect longevity in the CF1 variety in C1 was lower at 21ºC, while as the temperature decreased to 19ºC, there was an increase in the longevity of the insect in the same variety. This behavior not observed in the RAF variety. From instar two to instar five, the lifecycle of the insect is affected by the mean temperature of the crop cycle. In Cycle 1, the development of the mentioned nymph instars was affected (Table 5). Delimitation of the heat units required for the development of the stages of B. cockerelli in two tomato varieties.
Table 4 Means comparison of the cycle*variety interaction of the biological cycle of B. cockerelli grown in two tomato varieties in the greenhouse (October-December 2016 and January-March 2017).
| Interactions | No. of observations | Means | P-value | Mean temperature (°C) |
|---|---|---|---|---|
| Cycle 1*Charanda F1 | 21 | 77.2b | <0.0001 | 21.30 |
| Cycle 1*Rafaello | 21 | 85.2a | <0.0001 | 21.30 |
| Cycle 2*Charanda F1 | 21 | 89.9a | <0.0001 | 19.38 |
| Cycle 2*Rafaello | 21 | 87.9a | <0.0001 | 19.38 |
Means with the same letter are statistically equal, Tukey (P < 0.05).
Table 5 Effect of the comparison of temPC*instar minimum mean square of B. cockerelli lifespan in two tomato varieties grown in the greenhouse (October-December 2016 and January-March 2017).
| Stages | Effect | No. of observations | Minimum Mean squares | T-value | P-value |
|---|---|---|---|---|---|
| Adult | temPC2 vs temPC1 | 12 | 6.3 | 1.62 | 0.1108 |
| Egg | temPC2 vs temPC1 | 12 | -79 | 0.00 | 1.0000 |
| Nymph 1 | temPC2 vs temPC1 | 12 | -0.1 | -0.04 | 0.9662 |
| Nymph 2 | temPC2 vs temPC1 | 12 | 11.5 | 2.93 | 0.0046 |
| Nymph 3 | temPC2 vs temPC1 | 12 | 14.6 | 3.74 | 0.0004 |
| Nymph 4 | temPC2 vs temPC1 | 12 | 12 | 3.06 | 0.0032 |
| Nymph 5 | temPC2 vs temPC1 | 12 | 9.1 | 2.34 | 0.0223 |
temPC1 = mean temperature in Cycle 1, temPC2= mean temperature in Cycle 2.
The accumulated HU from egg to nymph 1 differed statistically in both crop cycles for the CF1 variety; HU was needed in the 2009 cycle to differentiate nymph 1 (F = 16.0, P = 0.01). In the rest of the instars, there were no significant differences between one cycle and the other (Table 6).
Table 6 Biological development of B. cockerelli chronologically (days) and physiologically (heat units) in the Charanda F1 variety in 2016 and 2017 cycles, grown under greenhouse conditions.
| Instar | P-value | F | gl | Sample size | HU duration (SE1) | ||
|---|---|---|---|---|---|---|---|
| 2016 | 2017 | 2016 | 2017 | ||||
| Egg-Nymph1 | 0.01 | 16.00 | 4 | 245 | 280 | 107.21±6.9 | 94.75±0.0 |
| Nymph1- Nymph2 | 0.21 | 2.22 | 4 | 172 | 269 | 64.85±5.0 | 167.74±9.0 |
| Nymph2- Nymph3 | 0.55 | 0.40 | 4 | 126 | 220 | 55.52±10.5 | 107.56±7.8 |
| Nymph3- Nymph4 | 0.88 | 0.02 | 4 | 92 | 139 | 62.67±10.6 | 77.30±11.7 |
| Nymph4- Nymph5 | 0.50 | 0.53 | 4 | 86 | 117 | 123.28±17.0 | 64.57±11.0 |
| Nymph5-Adult | 0.99 | 0.00 | 4 | 67 | 111 | 102.01±7.2 | 82.52±7.2 |
| Total | 453.39 | 594.44 | |||||
1SE= Standard error.
The accumulated HU in the RAF from egg to nymph was the same; there was no significant difference between both crop cycles. However, from nymph two to nymph three, and from four to five, there were significant differences in their development in both crop cycles, the test showed no significant differences. The two crop cycles differed statistically; more heat units were needed in the 2009 cycle to identify nymph 1. There were no significant differences between cycles regarding the rest of the instars (Table 7).
Table 7 Biological development of B. cockerelli chronologically (days) and physiologically (heat units) in Rafaello tomato variety in cycles 2016 and 2017, grown under greenhouse conditions.
| Instar | P-value | F | gl | Sample size | HU duration (SE1) | ||
|---|---|---|---|---|---|---|---|
| 2016 | 2017 | 2016 | 2017 | ||||
| Egg - Nymph1 | 0.01 | 16.00 | 4 | 236 | 274 | 121.14±0.0 | 112.23±0.0 |
| Nymph 1- Nymph 2 | 0.21 | 2.22 | 4 | 234 | 248 | 58.85±9.9 | 166.0±16.2 |
| Nymph 2 - Nymph 3 | 0.55 | 0.40 | 4 | 209 | 139 | 84.1±0.0 | 128.01±8.2 |
| Nymph 3 - Nymph 4 | 0.88 | 0.02 | 4 | 111 | 121 | 145.39±0.0 | 57.94±10.0 |
| Nymph 4 - Nymph 5 | 0.50 | 0.53 | 4 | 106 | 100 | 42.75±14.3 | 33.28±0.6 |
| Nymph 5 - Adult | 0.99 | 0.00 | 4 | 73 | 88 | 146.7±7.1 | 99.85±5.1 |
| Total | 598.94 | 597.38 | |||||
1SE= Standard error.
When analyzing the accumulated heat units data, considering the heat units means per cycles and variety, significant differences in both varieties (F = 20.23, P = 0.001) only found from nymph three to nymph four; there were no significant differences in the rest of the instars (Table 8). So, in general, in both varieties, the heat unit accumulation was the same from egg to adult.
Table 8 Accumulation of heat units in B. cockerelli in two tomato varieties: Charanda F1 and Rafaello, in two crop cycles.
| Stages | P-value | F | gl | Mean (SE1) | |
|---|---|---|---|---|---|
| Charanda F1 (Sample size) | Rafaello (Sample size) | ||||
| Egg - Nymph1 | 0.454 | 0.608 | 10 | 100.98±4.17 (525) |
116.68±1.99 (510) |
| Nymph1 - Nymph2 | 0.828 | 0.050 | 10 | 116.29±23.46 (441) |
112.45±25.44 (482) |
| Nymph2 - Nymph3 | 0.566 | 0.352 | 10 | 81.54±13.00 (346) |
106.06±10.48 (348) |
| Nymph3 - Nymph4 | 0.001 | 20.23 | 10 | 69.98±7.82 (231) |
101.66±20.06 (232) |
| Nymph4 - Nymph5 | 0.108 | 3.10 | 10 | 93.92±16.06 (203) |
38.015±6.75 (206) |
| Nymph5 - Adult | 0.052 | 4.85 | 10 | 92.27±6.33 (178) |
123.27±11.19 (161) |
| Total heat units | 464.98 | 598.13 | |||
1SE= Standard error.
DISCUSSION
The survival curves of B. cockerelli were similar in both crop cycles, and for each insect cycle, there being mortality in nymphs one to three. Similar results reported by Davis (1937), Liu and Trumble (2006), and Yang and Liu (2009). These authors mention that the highest mortality of nymphs happens from the first to third instars.
On the other hand, in the tomato variety and B. cockerelli instars, the biological cycle of the insects in C1 was found to be significantly different from those in C2. This behavior is as that reported by Yang and Liu (2009), Yang et al. (2010), and Vargas-Madríz et al. (2011), who mention that the host plants where B. cockerelli grows significantly influence the biological parameters.
The temPC-variety and cycle-variety interactions significantly affected the longevity of the insect populations; this is, the insect longevity was lower at a temperature of 21ºC. Although, when the temperature lowered to 19ºC, there was an increase in insect longevity in the same variety. These results coincide with those by List (1939) and Pavlista (2002), who mentions that temperature is a critical factor for the development of populations of this insect, with an optimum development being at temperatures of 27ºC. At higher temperatures, the eggs and nymphs can be affected. Moreover, Lehman (1930) mention that field populations of B. cockerelli are particularly abundant after warm winters and fresh environments. The time required for the development of one generation from egg to adult is 15 to 10 days. The optimum temperature for their development is 27ºC, while temperatures below 15ºC or over 32ºC affect their development and survival.
Heat units have been studied in a great variety of insects, among which is the whitefly, with optimum, maximum, and minimum values for thermal requirements for its development, varying between species and from one instar to another. In the case of Bemisia tabaci (Gennadius), the cited minimum and maximum temperature thresholds are 11ºC and 33ºC, respectively, while the highest development rate is at 28ºC. B. tabaci in cotton crops in the La Laguna region, Mexico, needs 275 heat units (HU = GDD) and a base temperature (bT) of 12ºC. However, other researchers mention that this species needs 315 thermal units (TU = GDD) to complete its cycle, and a bT of 10ºC (Ortiz et al., 2010).
Bactericera cockerelli shows a tendency like a whitefly. Several researchers have studied and determined different heat units for the development of B. cockerelli. For example, González-Maldonado and García-Gutiérrez (2012) indicate that the cycle of this insect is completed with 356 HU, while Ramírez et al. (2008) mention 336 HU. Likewise, Becerra (1989) determined, under laboratory conditions, the B. cockerelli completed its biological cycle with 335.8 HU and a threshold temperature of 7ºC. In the present research, the lifecycle of B. cockerelli completed with 464.98 and 598.13 HU in the CF1 and RF, respectively. Perhaps this difference is because incubation ranges from three to 15 days, depending on temperature (Pletsch, 1947).
For its part, Vaishampayan and Kogan (1980) state that the lifecycle of sweet potato whitefly, B. tabaci, can decrease its biological cycle when it develops on sick plants. This information opens the possibility that heat units are not only affected by temperature but also by other factors like the morphological condition of the plant, its nutritional state, and its general health. It suggested that the difference between varieties in this research might have been due to one of these mentioned conditions.
According to results, it concluded that the development of B. cockerelli on both tomato varieties is affected by the mean temperatures registered during the time of both crop cycles, and by the heat units observed for this insect. Another factor to consider could be the morphological characteristics of each variety, and the presence of trichomes and oils.

