











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La regeneración es la restitución de alguna estructura corporal dañada o perdida por otra igual que logra recuperar la forma y funcionalidad de la original. También es una estrategia de reproducción asexual en diversos grupos taxonómicos. La regeneración puede ocurrir por medio de dos mecanismos principales, epimorfosis y morfalaxis (Agata et al., 2007). La epimorfosis se caracteriza por la formación de un tejido indiferenciado que forma una estructura transitoria llamada blastema de regeneración y ha sido descrita en platelmintos, moluscos, anfibios y equinodermos (Ofiuroideos y Crinoideos). La morfalaxis se refiere al tipo de regeneración en que la parte perdida es remplazada por remodelación del tejido adyacente sin la formación de blastema. Esta regeneración ocurre en organismos como las hidras, tunicados y equinodermos (Asteroideos).
La autotomía es una estrategia de defensa común en muchos equinodermos y una forma de reproducción asexual (Candia-Carnevali, 2006). Por lo regular, en la clase Asteroidea la reproducción asexual ocurre mediante la fisión del organismo en dos partes que requieren porciones del disco central. Pocos géneros como Coscinasterias y Linckia se reproducen regularmente por autotomía de los brazos sin necesitar alguna parte del disco central (Edmonson, 1935; González-Moran, 2001). L. guildinguii (Gray, 1840) se distribuye en aguas tropicales poco profundas con excepción de la zona oriental del Pacífico y se caracteriza por tener un pequeño disco central con cinco brazos largos, cilíndricos y de punta roma. Esta especie se alimenta de partículas en suspensión y detritos y se reproduce asexualmente en etapa juvenil y sexual y asexualmente en etapa adulta (Stropes, 2003). Estas características hacen de L. guildinguii un modelo interesante para estudiar la regeneración como forma de reproducción asexual. En este estudio se aprovecharon estas ventajas para determinar experimentalmente la capacidad regenerativa de esta especie al igual que se ha hecho en otros organismos modelo más conocidos como la planaria y la hidra, en los cuales los primeros experimentos que se hicieron determinaron el tamaño mínimo de un fragmento capaz de regenerar y la relación de los ejes corporales con la regeneración.
Materiales y Métodos
La recolecta de L. guildinguii se realizó en el arrecife En Medio que forma parte del Sistema Arrecifal Lobos-Tuxpan (SALT), localizado a 13.5 kilómetros de la desembocadura del Río Tuxpan en el Oeste del Golfo de México (21.45707 N 97.151938 O). Se recolectaron 40 organismos de diferentes tamaños durante los meses de mayo, agosto y octubre del 2013 a una profundidad menor a un metro en la zona de pastos marinos o debajo de roca coralina. Estos organismos se mantuvieron en acuarios equipados con eliminador de proteínas, filtro biológico con piedra coralina y agua de la misma zona arrecifal, la cual se mantuvo a 35 ups pH 8.2 y 26 °C. Dado que las estrellas sobreviven en cautiverio por más de 6 meses y no es necesario administrarles algún alimento en específico toda vez que se mantienen con los detritos y partículas en suspensión que circulan en el acuario, sólo fue importante realizar cambios parciales de agua al acuario semanalmente y suplementarla con fórmulas comerciales de calcio y magnesio para invertebrados marinos.
Para realizar la amputación de los brazos, las estrellas fueron colocadas en una caja petri con agua de mar y con una navaja desechable para micrótomo se cortaron los brazos transversalmente sujetando con firmeza el brazo a cortar. Los brazos y los discos centrales obtenidos fueron devueltos cuanto antes al acuario donde permanecieron libres. Los brazos en regeneración fueron fijados con formaldehido a diferentes tiempos post-amputación para hacer el análisis histológico. Otro tipo de cortes rectangulares se llevaron a cabo, removiendo la parte proximal y distal del brazo y tomando en cuenta la longitud respecto a su anchura para mantener una relación 1:1 (cuadrados), 2:1, 3:1 4:1 etc. (Tabla 1). Otros brazos fueron cortados en diagonal, en un ángulo de 45o respecto al surco ambulacral. A algunos brazos amputados se les dejó una porción del disco central en forma triangular como rebanada de pastel. Finalmente otros cortes realizados fueron longitudinales, separando en dos mitades la parte distal de los brazos. Para cada tipo de corte se realizaron al menos cinco repeticiones. Para el análisis histológico las muestras se fijaron con formaldehído al 4% en agua de mar durante 24 hrs, posteriormente se descalcificaron con ácido acético al 10% durante 24 hrs y después se lavaron con agua destilada por 1 hr, para después impregnarlas con una solución de sacarosa al 15% y al 30%. A continuación se colocaron en gel para cortes en congelación durante 24 hrs a temperatura ambiente previo a la congelación para realizar los cortes. Estos se realizaron en un criostato (Microm HM 520) a -25 ºC y fueron de 10 mm de grosor. Los cortes fueron colectados en porta objetos previamente cubiertos con poli-L-lisina y procesados para la tinción tricrómica de Masson (Kit H-T15, Sigma-Aldrich).

Resultados
Se observó que estrellas de mar L. guildinguii de diferentes tamaños, desde 0.5 cm hasta 6.5 cm, tienen la capacidad de regenerar correctamente: el disco central regeneró los brazos amputados y cualquiera de los brazos regeneró una nueva estrella. Por la forma que adquieren estos brazos en regeneración se les conoce como "cometas" (Fig. 1 a, b) (Tabla 1). La capacidad para formar un nuevo organismo de forma asexual está presente en organismos muy pequeños aunque en este estudio, no determinamos lo que ocurre con organismos menores a 0.5 cm (ver Fig. 1i). A partir de fragmentos del brazo de diferentes longitudes también se regeneró una nueva estrella (Fig. 1c). Esta capacidad está presente a lo largo de todo el brazo, con excepción de la punta, pues fragmentos de la parte más distal, donde se adelgaza el brazo, no lograron regenerar y murieron. Después de 32 días empezaron a notarse a simple vista los nuevos organismos en el extremo proximal de los brazos, aunque fueron más notorios después de 40 días post amputación (Fig.1i).

Figuras 1a-m Distintos tipos de cortes hechos para explorar el potencial regenerativo de L. guildinguii. a) "Cometas" provenientes de brazos de diferentes tamaños. b) Discos centrales en regeneración. c) Cometas de fragmentos de diferente longitud. d) Fragmentos rectangulares. e) Fragmentos cuadrados, incapaces de regenerar. f) Cortes en forma triangular con un fragmento de disco central. g) Cortes diagonales regeneraron organismos orientados perpendicularmente. h) Cortes longitudinales que no formaron nuevos organismos. i) Estrella después de 40 días. j) Estrella separada del brazo original, la flecha señala un nuevo brazo. k) Puntas en regeneración indicadas por flechas. l) Pequeños brazos regenerados de novo. m) Nueva estrella después de 32 días, la flecha indica la boca. Barra de escala: 2 cm.
También se determinó lo que ocurría con fragmentos del brazo amputados distal y proximalmente. Se observó que a partir de fragmentos rectangulares de diferentes longitudes, se regenera la punta faltante en el lado distal y una nueva estrella en el lado proximal (Fig. 1d). En ocasiones se observó que fragmentos rectangulares de un poco menos del doble de largo que de ancho, regeneraron en dos estrellas, una en cada extremo (15. 3%, n=20) (Fig. 1d) (Tabla 1), pero en ningún caso se observó que regeneraran puntas en ambos extremos. Sin embargo, los fragmentos cuadrados no lograron regenerar y aunque permanecieron vivos por algunos días, terminaron fragmentándose y muriendo (Fig. 1e). Estos resultados nos indican que hay un tamaño mínimo del brazo, que debe ser de al menos el doble de largo que de ancho, capaz de regenerar correctamente un nuevo individuo sin que existan errores para interpretar el eje proximal-distal correctamente.
Se realizaron otros tipos de cortes para explorar cómo se interpreta en la regeneración el eje proximal-distal, encontrándose que los brazos amputados que incluían una porción del disco central en forma triangular y cuyo vértice era la boca original, logran regenerar una estrella en la parte distal faltante y la boca de la nueva estrella se posiciona justo tomando como referencia el inicio del surco ambulacral (Fig. 1f). En cambio los cortes en diagonal que no incluían parte del disco central, regeneraron en una nueva estrella perpendicular al eje proximal-distal, sin embargo la nueva boca también surge con referencia al surco ambulacral del brazo (Fig. 2g). Además se realizaron cortes longitudinales a lo largo del surco ambulacral separados del disco central, sin embargo estas mitades no sobrevivieron más de tres días. Por ello realizamos cortes longitudinales pero sin separarlos del disco, encontrando que estos casos no se formaron nuevas estrellas, sino que solamente ocurrió la regeneración de la mitad faltante (Fig. 1h). Con el estudio se determinó también que las nuevas estrellas que se forman en la parte distal de un brazo pueden sobrevivir si se les separa del mismo (Fig. 1i). En estos casos la pequeña estrella vuelve a regenerar un nuevo brazo en el sitio donde estaba el original (Fig. 1j). Estas estrellas a su vez mantienen la capacidad de regenerar los brazos que se le amputan (Fig. 1k), y los pequeños brazos (Fig. 1l) a su vez son capaces de regenerar una nueva estrella aún más pequeña (Fig. 1m) (Tabla 1). Sorprendentemente el proceso para formar una nueva estrella a partir tanto de brazos pequeños como grandes toma el mismo tiempo, alrededor de 32 días. Los experimentos realizados muestran que la regeneración de una nueva estrella ocurre siempre tomando como referencia el surco ambulacral y que la capacidad para generar un segundo nuevo individuo puede ocurrir a partir de los 32 días, cuando los nuevos brazos ya tienen un surco ambulacral bien definido.

Figuras 2a-f Apariencia externa del proceso de regeneración a lo largo de 32 días. a) Cavidad celómica expuesta. b) Contracción de la herida. c) Una semana después la contracción de la epidermis se ha recubierto la zona amputada. d) Hacia el día 14 se observa tejido nuevo de color claro arriba del surco ambulacral. e) Tejido nuevo en el día 20; la flecha señala el nuevo disco central. e) Nuevos brazos, señalados por flechas, son evidentes hacia el día 32. Barra de escala: 2.5 mm.
Durante estos eventos de regeneración no se observó la formación de tejido indiferenciado o blastema de regeneración y para confirmarlo, hicimos un seguimiento más detallado del proceso de regeneración con cortes histológicos, observando lo siguiente: en el momento posterior a la amputación del brazo, se observa la cavidad celómica del mismo totalmente expuesta al agua de mar (Fig. 2a). Después de unas horas se aprecia la contracción de la epidermis y de la región alrededor de la herida (Fig. 2b). Internamente esta contracción genera que las placas ambulacrales más cercanas a la herida entren en contacto con el techo de la cavidad celómica, que se caracteriza por tener una abundante acumulación de tejido conectivo, justo debajo de las placas carinales (Fig. 3a). Este contacto limita la entrada de agua al interior del brazo. El proceso de cerrado de la herida es lento (Fig. 2c, 3d) y hacia el día 14 el crecimiento de la epidermis sobre la herida es más notorio (Fig. 3d). Para el día 20 la nueva epidermis cubre totalmente la herida y se aprecia una protuberancia de tejido nuevo de color más claro por encima del surco ambulacral (Fig. 3e). Dentro del organismo ocurre la elongación y ramificación del ciego pilórico desde el sitio de angostamiento (donde las placas ambulacrales contactaron el techo del celoma) en dirección hacia donde se están formando los nuevos brazos (Fig. 3b-c). Hacia los días 22 a 26 este abultamiento de tejido nuevo adquiere un color marrón claro y aumenta de tamaño prefigurando el disco central de la nueva estrella (Fig. 3e). Alrededor del día 32 es que se observan las puntas de la nueva estrella proyectándose desde el nuevo disco central (Fig. 2f, 3f). Después del día 35 hay un aumento de tamaño del estómago cardiaco y los ciegos pilóricos aparecen bien delimitados dentro de la cavidad celómica de cada brazo. El análisis de los cortes histológicos mostró que este tipo de regeneración es de tipo morfaláctica, puesto que en ningún momento del proceso se apreciaron células de tipo mesenquimático indiferenciadas formando un blastema. Únicamente hay remodelación de los tejidos adyacentes, mismos que crecen lentamente formando el primordio del disco, a partir del cual en pocos días surgen los brazos de una nueva estrella.
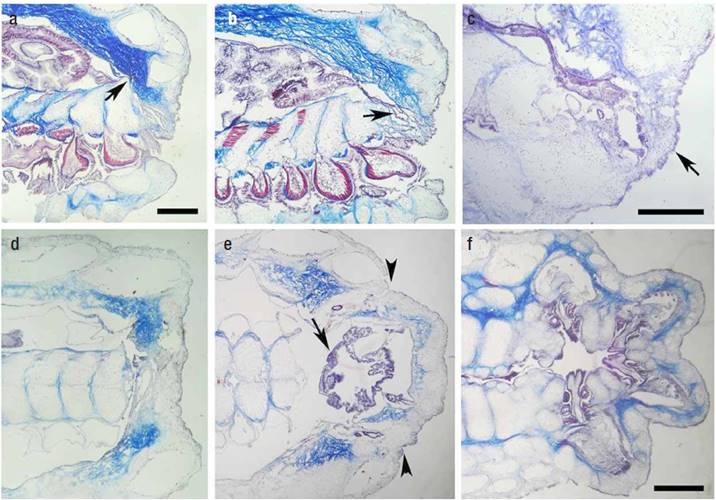
Figuras 3a-f Tinción tricrómica del proceso de regeneración en cortes longitudinales a lo largo del surco ambulacral, a, b, c o perpendiculares al surco ambulacral, d, e, f. a) Contracción de la herida y contacto de las placas ambulacrales con el techo de la cavidad celómica abundante en colágeno (flecha). Barra de escala: 500 mm. b) Día 20, extensión de los nuevos ciegos pilóricos. c) Día 22, tejido nuevo que prefigura el disco central (flecha). Barra de escala: 250 mm. d) Día 14, herida epitelizada. e) Día 26, se aprecia el nuevo disco central indicado por las cabezas de flecha. La flecha señala al estómago cardiaco. f) Día 32, los nuevos brazos están completamente formados. Barra de escala: 500 mm. En todas las imágenes la parte proximal esta hacia el lado derecho.
Discusión
El potencial para regenerar en los metazoarios ha sido explicado a través de un modelo de "barras y estrellas" en el que se ordenan diversos organismos dependiendo de su mayor o menor capacidad regenerativa (Rinkevich y Rinkevich, 2013). Por el lado de las barras están los organismos que solamente pueden regenerar partes del soma y no la línea germinal y que además no tienen células adultas troncales totipotentes. Por el lado de las estrellas están los seres con capacidad para regenerar tanto el soma como la línea germinal, tienen la capacidad de restablecer los ejes corporales y presentan células troncales totipotentes adultas. Los datos mostrados indican que L. guildinguii pertenece a este último grupo de organismos. Aunque la temperatura puede influir la velocidad de regeneración, en general las estrellas de mar se regeneran más lentamente comparadas con las ofiuras. Por ejemplo, para el caso de la ofiura Ophiothrix lineata (Lyman, 1860), la regeneración del brazo ocurre en poco menos de 10 días (Alarcón et al., 2013). A pesar de que en ambas clases el desprendimiento de los brazos ocurre por autotomía, una diferencia importante es que las ofiuras presentan regeneración epimórfica y forman rápidamente un blastema de regeneración, sin embargo hasta el momento no se conoce alguna especie que a partir del brazo perdido, regenere un nuevo organismo como ocurre en los Asteroideos.
A pesar de la sorprendente capacidad de regeneración del género Linckia, no se había explorado en detalle la capacidad de regeneración en este organismo deuterostomado de simetría radial secundaria. Organismos protostomados como las planarias (bilaterales) e hidras (radiales), han sido modelos importantes para estudiar la regeneración y reproducción asexual. Así, los primeros estudios hechos con estos organismos se enfocaron en entender cómo se restablecen los patrones corporales y en determinar cuál era el tamaño mínimo de un fragmento capaz de regenerar un organismo completo (Lenhoff & Lenhoff, 1986; Reddien & Sánchez-Alvarado, 2004). Uno de los fenotipos más interesantes que observamos fue la regeneración de dos estrellas a partir de fragmentos rectangulares cortos. Pudiéramos hacer una analogía de este tipo de regeneración con la conocida como "Janus heads" en planarias, en la que fragmentos cortos anteriores regeneran dos cabezas en ambos extremos (Morgan,1898). Este tipo de alteración indica que se requiere de una longitud de al menos el doble de largo respecto a lo ancho para que se especifique correctamente la polaridad del las partes a regenerar. L. guildinguii tiene una menor capacidad para regenerar que la planaria, pues no es capaz de formar nuevos organismos a partir de segmentos longitudinales del brazo. Aún así, pocos organismos presentan una capacidad de regeneración tan notable y esta especie además puede producir nuevamente un organismo a partir de pequeños brazos apenas diferenciados.
Durante la regeneración es necesario que solamente se regenere la parte que se perdió y que se mantenga la correcta identidad y disposición espacial original. En el caso de la regeneración que da origen a un nuevo organismo, deben redefinirse todos los ejes corporales. La formación de los ejes corporales se ha estudiado principalmente durante el desarrollo embrionario pero poco se sabe de cómo ocurre post-embrionariamente (Cuervo & Chimal-Monroy, 2013). Un hallazgo importante del presente trabajo es que poco después de la amputación del brazo y de la contracción de la herida, las placas ambulacrales se juntan con el techo de la cavidad celómica generando un angostamiento en dirección oral-aboral justo en medio, sobre el surco ambulacral. En este momento la simetría del brazo es bilateral con un eje proximal-distal (Ji et al., 2012), sin embargo debe ocurrir en algún momento, previo a la regeneración, el restablecimiento de las señales moleculares y procesos que permitan redefinir un eje penta radial en ese sitio para poder dar inicio al nuevo organismo. Nuestros resultados sugieren que este sitio de angostamiento, es el sitio clave donde debe ocurrir la redefinición de ejes para establecer el punto a partir del cual se formará la boca como centro de la simetría radial del nuevo individuo (ver Fig. 3e).
Estudiar la formación de los ejes corporales a nivel molecular es factible con este modelo de regeneración comparando los extremos distal y proximal utilizando las nuevas tecnologías de secuenciación. Con estrategias como ésta se empezaría a entender cómo es que se establecen los ejes corporales en este organismo de simetría radial secundaria, que en un inicio proviene de una larva bilateral.

