











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink1. Introduction
Central Mexico is characterized by the presence of a volcanic chain that crosses the country from E to W at around 18° to 22°N. This volcanic chain is known as the Trans-Mexican Volcanic Belt (TMVB) and is the largest Neogene volcanic arc of North America, with a length of 1000 km and an area of 160000 km2 (Ferrari et al., 2012). Along this wide surface, topographic heterogeneity favors the presence of several lacustrine basins across a broad range of climatic conditions which are determined mostly by the significant altitudinal gradient (the highest peaks reach > 5000 masl). Besides temperature, this topographic gradient is also associated with differences in precipitation and/or evaporation, with generally drier climates on the western, lower altitude basins; but the driest region, under a rain shadow effect, is on the easternmost high-altitude basin (Oriental).
Throughout the history of the region the lakes along the TMVB have been core areas for cultural development (Faugère-Kalfon, 1996; Niederberger, 1979; Serra-Puche, 1988) as they are favorable locations for agriculture, fishing, hunting, and recreational activities. Thus, human impact around these lakes has also a long history, which in modern times has been sharpened by stressors such as global warming, urban development and widespread changes in land use (Alcocer and Bernal-Brooks, 2010).
Lakes are sensitive ecosystems to changes in their environment (Adrian et al., 2009) and we know that in several of these lakes the impact of modern stressors has already caused, for example, important reductions in lake levels or increases in turbidity and trophic status (Alcocer et al., 2000a; Caballero et al., 2006; Kienel et al., 2009; Komárková and Tavera, 2003). These changes can generate social and economic problems as well as a significant reduction in the biodiversity of these ecosystems, so it is important to have basic limnological information that allows a better understanding of these aquatic systems as a basis for further ecological studies, and also for the management and restoration of the water resources they represent. In some of these lakes the changes have been very quick, but the rate of change is difficult to evaluate as there are relatively few limnological studies undertaken in the area to allow understanding the degree and speed of such transitions. Limnological information about these lakes is therefore relevant to document their current status, which is necessary as a reference when evaluating past conditions. In the same way, it is important to identify the organisms that live in these waterbodies, especially those that can be used as bioindicators in modern and paleo-environmental research given their high potential of preservation in the sediments. Phytoplankton is probably the most sensitive but also the remains of diatoms, testate amoebae, cladocerans and ostracodes preserved in sediments can be very useful to determine the conditions of the water body in which they lived. All these groups have short life cycles, so they quickly respond to environmental changes, enabling to understand the present conditions of the lacustrine ecosystem and open the possibility of evaluating recent changes through paleolimnological methods by, for example, comparing the bioindicators contents in modern and pre-human impact sediment samples.
Previous limnological studies in the TMBV region have focused on a few of its lakes, mainly the largest and/or deepest (e.g. Alchichica: Alcocer et al., 2000b; Kaźmierczak et al., 2011, Chapala: Hansen and van Afferden, 2001; Quiroz et al., 2008, Pátzcuaro: Alcocer and Bernal-Brooks, 2002; Bischoff et al., 2004, Zirahuén: Chacon-Torres and Rosas-Monge, 1998; Bernal-Brooks and MacCrimmon, 2000a). Scarce limnological information is available for most of the smaller waterbodies, but even for the best characterized lakes, results are often difficult to compare, as studies are carried out following different methodologies and/or only report results on specific physico-chemical characteristics or biological groups. Only a handful of studies have been performed simultaneously in more than one lake to allow for regional comparisons (e.g. Armienta et al., 2008; Davies et al., 2002).
The aims of this study are, therefore (1) to describe the main morphometric and limnological characteristics of 30 waterbodies on the TMVB in central Mexico using standardized analytical methods so that the data are easily comparable, (2) to provide information about their current nutrient levels and trophic state, (3) to report the dominant aquatic bioindicator taxa (phytoplankton and sedimentary diatoms, testate amoebae, cladocerans and ostracodes) present at each site as a contribution to future ecological and paleoecological research, (4) to assess the influence of climate and geographic location over the limnological variables of these waterbodies.
2. Materials and methods
Thirty waterbodies located along the TMVB (Figure 1) were sampled once from late summer to early autumn (lakes El Sol and La Luna in August 2010 and the rest of the lakes between June and October 2011). The geographical location (latitude, longitude, altitude) of each site was taken using a handheld navigator (GARMIN GPSMAP 62 stc) and confirmed in Google Earth®. All of them were studied following uniform methodologies for field work, laboratory analyses and taxonomic determinations of bioindicators (phytoplankton and sedimentary diatoms, testate amoebae, cladocerans and ostracodes). The limnological and biological data thus obtained were complemented with additional information and compiled to generate an individual chart for each lake, summary data are also presented in Table 1. Charts display information on climate, limnology, main aquatic bioindicator taxa present, and previous published work. Finally, numerical analyses were used to synthesize and analyze the limnological and climatic information. Description of items included in charts, and of numerical analysis performed follows:
2.1. Climate
For all sites, meteorological data such as mean annual temperature, annual temperature range, total annual precipitation and annual evaporation were taken from the closest meteorological stations (SMN, 2015). Climate types for each site were identified according to the INEGI climate map (INEGI, 2008) which follows García, 2004.
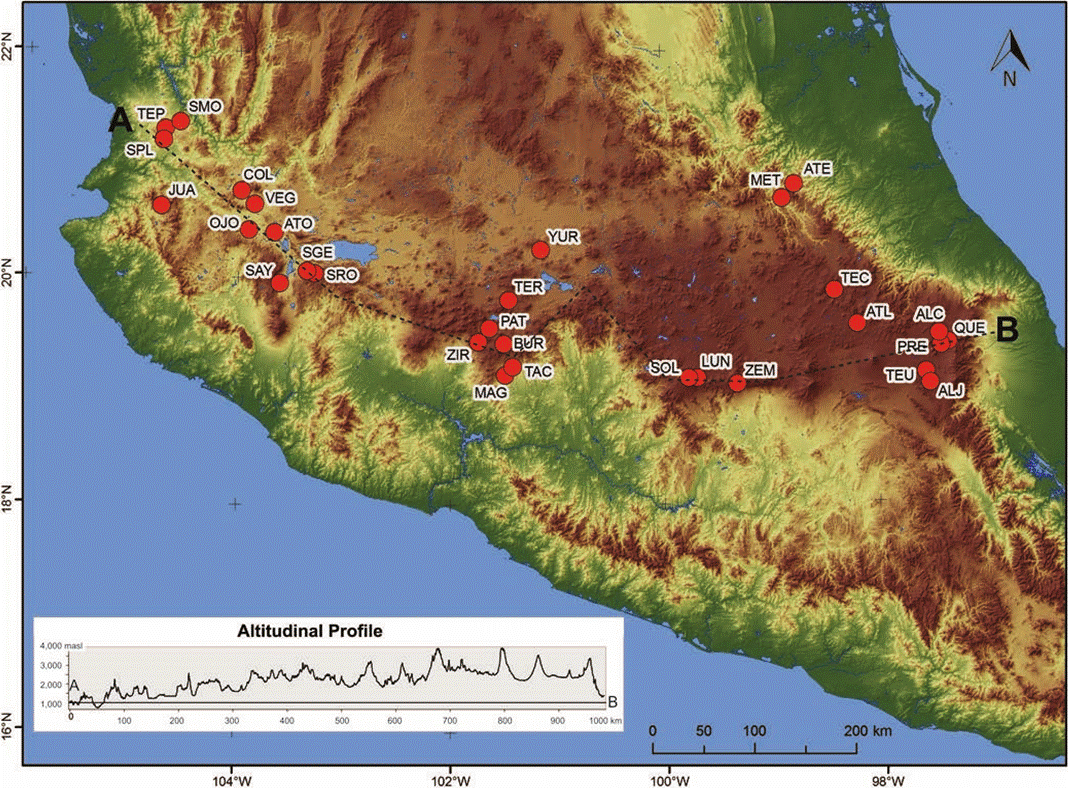
Figure 1 Map with the location of the 30 lakes studied along the Trans-Mexican Volcanic Belt, central Mexico. Abbreviations correspond those in Table 1
Table 1 Main limnological characteristics of 30 lakes studied along the Trans-Mexican Volcanic Belt, central Mexico. Salinity and trophic categories as defined in Table 1.
| Lake | Tag | Type | Area (ha) | Máximum depth (m) | Relative depth (%) | Dominant anions | Dominant cations | pH | Salinity category | Trophic category |
|---|---|---|---|---|---|---|---|---|---|---|
| Alberca de Tacámbaro | TAC | Volcanic (crater) | 8.2 | 28 | 8.7 | HCO3- | Mg2+ | 7.8 | Freshwater | Eutrophic |
| Alberca de Teremendo | TER | Volcanic (crater) | 15 | 9 | 2.1 | HCO3- | Na+ - Mg2+ | 8.3 | Freshwater | Hypertrophic |
| Alchichica | ALC | Volcanic (maar) | 200 | 62 | 3.8 | Cl- | Na+ | 8.3 | Hyposaline | Mesotrophic |
| Aljojuca | ALJ | Volcanic (maar) | 42 | 51 | 7 | HCO3- | Na+ | 9.5 | Subsaline | Mesotrophic |
| Atezca | ATE | Volcanic (dam) | 27 | 16 | 2.7 | HCO3- | Ca2+ | 9.3 | Freshwater | Mesotrophic |
| Atlangatepec | ATL | Reservoir (dike) | 800 | 7 | 0.2 | HCO3- | Na+ | 7.7 | Freshwater | Eutrophic |
| Atotonilco | ATO | Tectonic | 1370 | 0.5 | <0.1 | HCO3- | Na+ | 9.6 | Hyposaline | Mesotrophic |
| Burro | BUR | Probably volcanic | 9 | 1 | 0.3 | HCO3- | Na+ | 6.9 | Freshwater | Eutrophic |
| Colorada | COL | Reservoir (dike) | 410 | 3 | 0.1 | HCO3- | Ca2+ | 7.8 | Freshwater | Hypertrophic |
| El Sol | SOL | Volcanic (crater) | 18 | 15 | 3.2 | HCO3- | Ca2+ | 7.2 | Freshwater | Oligotrophic |
| Juanacatlán | JUA | Volcanic (dam) | 20 | 25 | 5 | HCO3- | Ca2+ | 9.2 | Freshwater | Mesotrophic |
| La Luna | LUN | Volcanic (crater) | 2.5 | 10 | 5.6 | HCO3- | Ca2+ | 7.7 | Freshwater | Ultra-oligotrophic |
| La Magdalena | MAG | Reservoir (dike) | 22 | 4 | 0.8 | HCO3- | Ca2+ | 8.8 | Freshwater | Hypertrophic |
| La Preciosa | PRE | Volcanic (maar) | 91 | 46 | 4.3 | HCO3- | Mg2+ | 9.3 | Subsaline | Oligotrophic |
| La Vega | VEG | Reservoir (dike) | 1610 | 6 | 0.1 | HCO3- | Na+ | 8.5 | Freshwater | Hypertrophic |
| Metztitlán | MET | Tectonic | 2940 | 10 | 0.2 | HCO3- | Ca2+ | 8.9 | Freshwater | Eutrophic |
| Ojo de Agua | OJO | Reservoir (dam) | 58 | 1.5 | 0.2 | HCO3- | Ca2+ | 8.5 | Freshwater | Eutrophic |
| Pátzcuaro | PAT | Volcanic (dam) | 10000 | 9.4 | 0.1 | HCO3- | Na+ | 8.3 | Subsaline | Hypertrophic |
| Quechulac | QUE | Volcanic (maar) | 64 | 40 | 4.4 | HCO3- | Mg2+ | 9 | Freshwater | Oligotrophic |
| San Pedro Lagunillas | SPL | Volcanic (dam) | 296 | 7 | 0.4 | HCO3- | Na+ | 8.2 | Freshwater | Hypertrophic |
| Santa Gertrudis | SGE | Reservoir (dike) | 50 | 2.3 | 0.3 | HCO3- | Ca2+ - Mg2+ - Na+ | 8 | Freshwater | Eutrophic |
| Santa María del Oro | SMO | Volcanic (crater) | 370 | 65 | 3 | HCO3- - Cl- | Na+ | 8.6 | Subsaline | Mesotrophic |
| Santa Rosa | SRO | Reservoir (dam) | 1.3 | 1.8 | 1.4 | HCO3- | Mg2+ | 9 | Freshwater | Mesotrophic |
| Sayula | SAY | Tectonic | 10700 | 1 | <0.1 | HCO3- - Cl- | Na+ | 9.3 | Subsaline | Mesotrophic |
| Tecocomulco | TEC | Tectonic | 1900 | 2 | 0.1 | HCO3- | Na+ | 8.8 | Freshwater | Eutrophic |
| Tecuitlapa | TEU | Volcanic (maar) | 18 | 2.5 | 0.1 | CO32- | Na+ | 10.3 | Subsaline | Hypertrophic |
| Tepetiltic | TEP | Volcanic (crater) | 132 | 2.5 | 0.2 | HCO3- | Ca2+ | 8.3 | Freshwater | Hypertrophic |
| Yuriria | YUR | Reservoir (dam and river diversion) | 5000 | 2.3 | <0.1 | HCO3- | Na+ | 8 | Freshwater | Eutrophic |
| Zempoala | ZEM | Volcanic (dam) | 10 | 8 | 2.2 | HCO3- | Ca2+ | 8.8 | Freshwater | Mesotrophic |
| Zirahuén | ZIR | Volcanic (dam) | 930 | 40 | 1.2 | HCO3- | Ca2+ - Mg2+ - Na+ | 6.5 | Freshwater | Oligotrophic |
2.2. Limnology
The lake’s origin was determined following bibliographical sources or inferred from field observations and Google Earth® satellite images in the case that there were not previous studies. The categories identified were: tectonic (by faulting), volcanic (crater, maars and natural dam) and reservoir (dike, artificial dam and river diversion). The surface of each water body was measured in the Google Earth® satellite images and when possible confirmed with bibliographical data.
Water depth (m) was measured in the field using a portable depth sounder (Speedtech Instruments), but not always the maximum depth spot at each site was located. Maximum water depth for all sites was taken from bibliographical data when available or from our own depth measurements. Relative depth (Zr in %) was calculated with the formula:
where Zmax = maximum depth, and Ao = surface area (Wetzel and Likens, 1991).
The lake’s mixing pattern was taken from bibliographic data when available, otherwise it was assumed to be warm monomictic if water depth was > 8 m, relative depth was >1 % and our temperature depth profiles confirmed stratification; if these criteria were not fulfilled, the lake was assumed to be warm polymictic (Lewis, 2000). If a thermocline and/or oxycline were detected, the depth (m) at which they occurred was specified. Water transparency was measured in the field as visibility (m) using a Secchi disk.
Temperature (°C), oxygen concentration (mg/L), pH and electric conductivity (µS/cm) depth profiles were also measured in the field using a multiparametric probe (Hydrolab Quanta G). Temperature and oxygen data are presented as depth profiles in lakes where the sampling site was > 6 m deep, or in table format for shallower locations. Electric conductivity and pH are presented in tables, showing a littoral site (only measurement in small, shallow lakes), and a centric site which could have only surface (shallow lakes), or surface and bottom data (if depth ≥ 3 m).
Water samples for chemical determinations and for phytoplankton analyses were collected using a Van Dorn-type sampling bottle from a littoral and from a centric site, with samples from surface (0.5 m) and bottom of the water column (1 m above the sediments if depth ≥ 3 m). Samples for cation determinations were acidified using concentrated (55 %) HNO3. All samples were kept in refrigeration until analyzed in the laboratory. Major ions analyses were performed following standard procedures (APHA, 1995, 1998; APHA et al., 2005; Armienta et al., 2008).
Major ions concentrations are expressed in mg/L but these data were also transformed to meq/L and then to percentages to determine ionic dominance (ion’s relative concentrations), which was determined merging [HCO3-] + [CO32-] and [Na+] + [K+].
Total dissolved inorganic carbon (DIC) was measured with an AutoMate carbonate preparation device. Approximately 5 mL of sample were weighed into septum top tubes and placed into the carousel. To purge the sample vial of atmospheric gas, a double needle assembly and CO2-free nitrogen carrier gas were used. The sample vial was injected with acid and evolved CO2 was carried through a silver nitrate scrubber to the coulometer where total C was measured.
Samples for ammonium and nitrates were acidified using concentrated (98 %) H2SO4. Ammonium (N-NH4, Nessler’s method), nitrites (NO2, diazotization), nitrates (N-NO3, brucine colorimetric method), total phosphorus (PT, persulfate digestion) and orthophosphate (P-PO4, ascorbic acid method) were determined in a Thermo Scientific GENESYS 20 Visible spectrophotometer. Silica (SiO2) was colorimetrically determined by the molybdosilicate method. Nutrients concentrations are expressed as mg/L, merging [N-NH4] + [N-NO3] + [N-NO2] as dissolved inorganic nitrogen (DIN). Nutrient concentrations were also transformed to µM and molar nutrient ratios were calculated (DIN:PT, DIN:P-PO4, SiO2:DIN, SiO2:P-PO4).
Samples for chlorophyll a determinations were filtered in the field with a Whatman GF/C membrane and filters were kept in refrigeration and darkness until arriving to the laboratory. Extraction was made using methanol (90 %), concentrations were measured with a spectrophotometer (Thermo Scientific Genesys 20 Visible) and determined using Holden’s equations (Meeks, 1974); chlorophyll a concentrations are expressed as mg/m3. Salinity category for each lake was determined following their total dissolved solids (TDS) values (Fritz, 2007; Kolbe, 1927) and trophic category was determined based on their superficial chlorophyll a concentrations (OECD, 1982) (Table 2).
Table 2 Boundary values criteria for salinity (Kolbe, 1927; Fritz, 2007) and trophic (OECD, 1982) categories.
| Salinity | |||||
| Category | Freshwater | Subsaline | Hyposaline | Mesosaline | Hypersaline |
| TDS (mg/L) | ≤ 500 | 500 - 3000 | 3000 - 20000 | 20000 - 50000 | > 50000 |
| Trophic category | |||||
| Category | Ultra-oligotrophic | Oligotrophic | Mesotrophic | Eutrophic | Hypertrophic |
| Chl a* [mg/m3] | ≤ 2.5 | 2.5 - 8.0 | 8.0 - 25 | 25 - 75 | ≥ 75 |
*annual maximum of chlorophyll.
2.3. Biological analysis
Water samples for phytoplankton analyses were collected at the same time as those for chemical determinations and preserved using Lugol’s solution. Two aliquots of 0.1 mL from each phytoplankton sample were analyzed using a compound microscope (Nikon Eclipse 80i) at 600x to 1000x. Semiquantitative abundance determinations were based on the following rankings: rare (1 specimen), scarce (2 - 10), abundant (11 - 50) and very abundant (> 50). In this study, only the species falling in the last two categories (abundant and very abundant) are reported. Organisms were identified using specialized bibliography for each (Komárek and Anagnostidis, 1999, 2005; Komárek et al., 1983; Krammer and Lange-Bertalot, 1991a, b; Krammer and Lange-Bertalot, 1997; Krammer and Lange-Bertalot, 1999; Prescott, 1962).
Surface sediments were collected from all lakes at a littoral (only sample in small, shallow lakes) and a central station for biological (diatoms, testate amoebae, cladocerans and ostracodes) determinations; if water depth was very shallow (< 1 m) sediment was collected directly with a spatula and if deeper with an Ekman dredge, taking care of collecting only the topmost sediment. Sediments samples for testate amoebae and ostracodes were preserved with anhydrous ethanol and all samples were kept refrigerated until analyzed in the laboratory.
Diatoms were extracted from 1 cm3 sediment samples. Sediment was freeze-dried, weighed and treated with HCl, H2O2 and HNO3 to eliminate carbonates and organic matter. Permanent slides were mounted with 200 µL aliquots of final solution, using Naphrax® as mounting medium. Counts of a minimum of 400 individuals were performed using an Olympus BX50 microscope (1000x). Species relative abundances were calculated and only the most abundant species, those accounting for 50 % of the counts in each sample, were reported. Species identification was based on specialized literature (Gasse, 1986; Krammer and Lange-Bertalot, 1991a, b; Krammer and Lange-Bertalot, 1997; Krammer and Lange-Bertalot, 1999).
Testate amoebae were extracted from 2 cm3 samples with a fine brush under a stereomicroscope (Zeiss STEMI 2000-C Schott SeriesEasyLED) at 64x to 100x. Identification was made following specialized literature (Alcocer et al., 2000b; Charman et al., 2000; Kumar and Dalby, 1998; Lee et al., 2000; Ogden and Hedley, 1980). All the tests found were counted, but only the genera or species for which more than 100 specimens were found are included in this study.
For cladocerans analysis, 1 cm3 of sediment was heated in a 10 % KOH solution and stirred for 20 - 30 min. Treated samples were washed through a 38 µm sieve, and then analyzed using an Olympus BX50 microscope at 100x to 400x. All Cladocera remains were counted (headshields, shells, ephippia, postabdomens) and the most abundant body part of each species was considered representative of the number of individuals, with a minimum of 200 individuals counted per sample. Identification and nomenclature of Cladocera remains was based on specialized literature (Cervantes-Martinez et al., 2000; Elías-Gutiérrez et al., 2008; Sinev and Zawisza, 2013). In this study, only the taxon with the highest frequency in each sample was reported, except in the samples were two species had equally high abundances, in these cases both taxa were reported.
Ostracode abundance was determined in 50 cm3 sediment samples by washing them through a 63 µm mesh sieve and separating adult valves using fine brushes under an Olympus SZX12 stereoscope (40x to 60x). Ostracodes valves were stored in micropaleontological slides, specimens with well-preserved soft parts were kept at 5 °C in small Eppendorf vials filled with 96 % ethanol. For identification to genus level, hard parts were studied in detail and well-preserved specimens were dissected (Meisch, 2000). Identifications followed specialized literature (Forester et al., 2015; Pérez et al., 2011; Pérez et al., 2015). In this work, only the three most abundant genera (> 100 adult and juvenile valves in 50 cm3 of sediment) at each site were reported.
2.4. Previous work
A bibliography search for published papers for each lake was done; if the waterbodies had been extensively studied, only some of the most recent publications were selected.
2.5. Numerical analysis
The dataset was synthesized through a Principal Component Analysis (PCA), which was performed using geographic (latitude, longitude, altitude), climatic (mean annual temperature, mean annual precipitation, mean annual evaporation) and limnological data (lake area, maximum depth, water transparency, chlorophyll a, average surface and bottom TDS, DIN and TP and average water column temperature, dissolved oxygen, pH and electric conductivity) from the 30 studied localities. Subsequently, another PCA was run excluding geographic location (latitude, longitude and altitude). The ordination of sites in both PCAs was compared through the Procrustes technique, a method that consists of fixing the first ordination, whereas the second is rotated using its origin as the vertex and aiming to minimize the distance between the same sites in both ordinations (Borcard et al., 2011). All analyses were performed using Package ‘vegan’ (Oksanen et al., 2015) in R (R Development Core Team, 2009).
3. Results
An example of the charts made for each lake is presented in Figure 2, and the full set of individual charts for all the lakes is presented in alphabetic order in Electronic Supplement 1. Several of these waterbodies have been listed as Ramsar sites (Atotonilco, Sayula, Atlangatepec, Tecocomulco, Yuriria, La Vega, Metztitlán and Pátzcuaro), which shows the increasing concern for studying and preserving these ecosystems. However, to the best of our knowledge, this is the first published record of the basic limnological characteristics of some of the lakes (Alberca de Teremendo, Burro, Colorada, La Magdalena, Ojo de Agua, Santa Rosa, Santa Gertrudis, Tepetiltic) and for many this is the first record of their aquatic bioindicator’s diversity, mostly for testate amoebae and cladocerans (full taxonomic and distribution list in Electronic Supplement 2). A summary of the lake’s main characteristics is presented in Table 1.
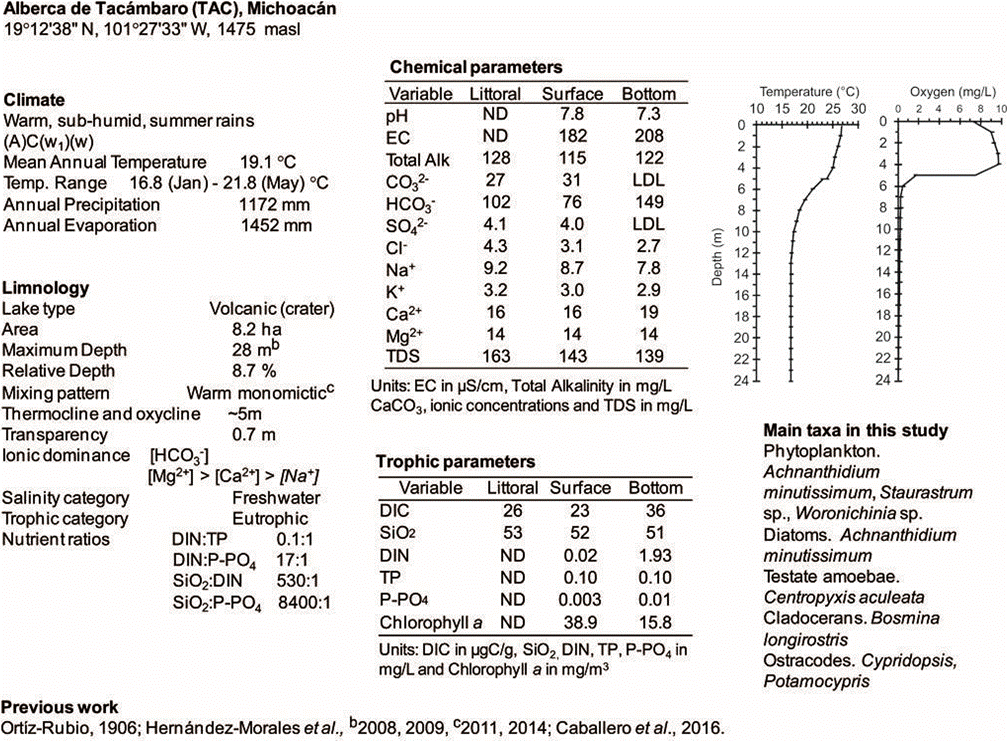
Figure 2 Climatic and limnological data of Alberca de Tacámbaro. a and b indicate the bibliographical source of the data. Ionic dominance includes ions present at >5 % relative concentrations in meq/L, in italics are ions between 5 and 25 %; “-“ was used when ionic relative concentrations were similar; “>” when they were less than double and “>>” when they were higher than double. LDL = Lower Detection Limit: 0.01 mg/L CO32-, 4 mg/L SO42-. ND = Not Determined. Date of sampling: June 17, 2011.
4. Discussion
4.1. Distribution and morphometric characteristics
The study covered 30 lakes within a latitudinal band from 19° 00’ to 21° 20’ N and in longitude from 97° 20’ to 104° 40’ W. They were distributed along a ~ 3550 m altitudinal gradient, from 737 masl (Santa María del Oro) to 4283 masl (La Luna and El Sol). Most of the lakes were volcanic in origin (5 maars, 6 crater lakes and 7 volcanic dams), followed by reservoirs (8) and tectonic lakes (4). About a third of the lakes were shallow (< 6 m), some of them with just or under 1 m deep, these shallower lakes were mostly reservoirs (e.g. Ojo de Agua) or tectonic in origin (e.g. Atotonilco). Another third included some of the deepest lakes (> 25 m) in central Mexico, and these were mostly volcanic craters or maars, like Alchichica and Santa María del Oro (62 and 65 m deep, respectively).
There was a good agreement between the measured and the bibliographic depths in all the lakes. Regarding areas, those estimated using Google Earth® also had a good agreement with the available bibliographical data, with discrepancies only at four sites. For lake Tecocomulco the bibliography reports a slightly smaller surface (1900 vs. 1769 ha, Ramsar, 2003), this relatively small difference (< 10 %) can be explained as Tecocomulco is a large lake with ample inundation areas which can be flooded or dry depending on season and/or inter-annual climatic variability. For La Vega and Yuriria the bibliography reports larger areas (La Vega: 1610 vs. 1950 ha, Ramsar, 2011; and Yuriria: 7200 vs. 9500 ha, Ramos and Novelo, 1993) however, these water bodies are dams, which areas can easily change due to human management. For lake La Luna, the bibliography also reports a larger area (2.5 vs. 3.1 ha, Alcocer et al., 2004), but there is no discrepancy regarding its depth to confirm any change in this lake’s morphometry.
4.2. Hydrochemistry
Waterbodies covered a wide range of hydrochemical characteristics; at the time of sampling (rainy season) most of them were alkaline (pH > 7.5), and some even reached extremely alkaline pH values (> 9.5, Alchichica, Aljojuca, Atotonilco, Atezca, Tecuitlapa); only a few had pH values around or below 7 (El Sol 7.2, Burro 6.9, Zirahuén 6.5, La Luna 6.3).
Most of them were freshwater lakes, nine with particularly low mineralization (TDS < 100 mg/L, Atezca, Burro, El Sol, Juanacatlán, La Magdalena, La Luna, Tepetiltic, Zempoala, Zirahuén) but there were also six subsaline (Aljojuca, La Preciosa, Pátzcuaro, Santa María del Oro, Sayula, Tecuitlapa) and two hyposaline (Alchichica, Atotonilco).
Except for Colorada, the lakes had low proportions of sulfates (< 25 %) and, except for Alchichica, they were dominated by carbonates and bicarbonates (Figure 3). The high sulfate proportion in Colorada is very unusual and could be a sign of anthropogenic pollution (Davies et al., 2002). Amongst the subsaline and hyposaline lakes there is a trend towards higher [Cl-] proportions, with Alchichica, the only lake dominated by chlorides, towards the end of this trend. Subsaline and hyposaline waterbodies had also low [Ca2+] proportions (Figure 3) and except for La Preciosa (which was [Mg2+] dominated) they were dominated by [Na+] + [K+]. Freshwater lakes do not have a clear ionic dominance, some were dominated by [Na+] + [K+] (La Vega, San Pedro Lagunillas, Tecocomulco, Yuriria, Alberca de Teremendo, Atlangatepec), some by [Ca2+] (Colorada, El Sol, La Luna, La Magdalena, Juanacatlán, Metztitlán, Ojo de Agua, Santa Gertrudis Tepetiltic, Zempoala, Zirahuén), and others by [Mg2+] (Santa Rosa, Tacámbaro, Quechulac); however, those with the lowest TDS (< 100 mg/l) were mostly [Ca2+] dominated. This ionic dominance pattern suggest that the lowest TDS lakes represent the least evolved waters (Hardie and Eugster, 1970; Kilham, 1990), with ionic dominance determined mostly by weathering processes of the basaltic andesite rocks that dominate in the TMVB (Guilbaud et al., 2012), while the ionic composition of subsaline and hyposaline lakes is mostly controlled by evaporative concentration and carbonate precipitation (Eugster and Hardie, 1978; Kilham, 1990) leading to [Cl-] -[CO32-] - [Na+] brines such as in Sayula or Alchichica.
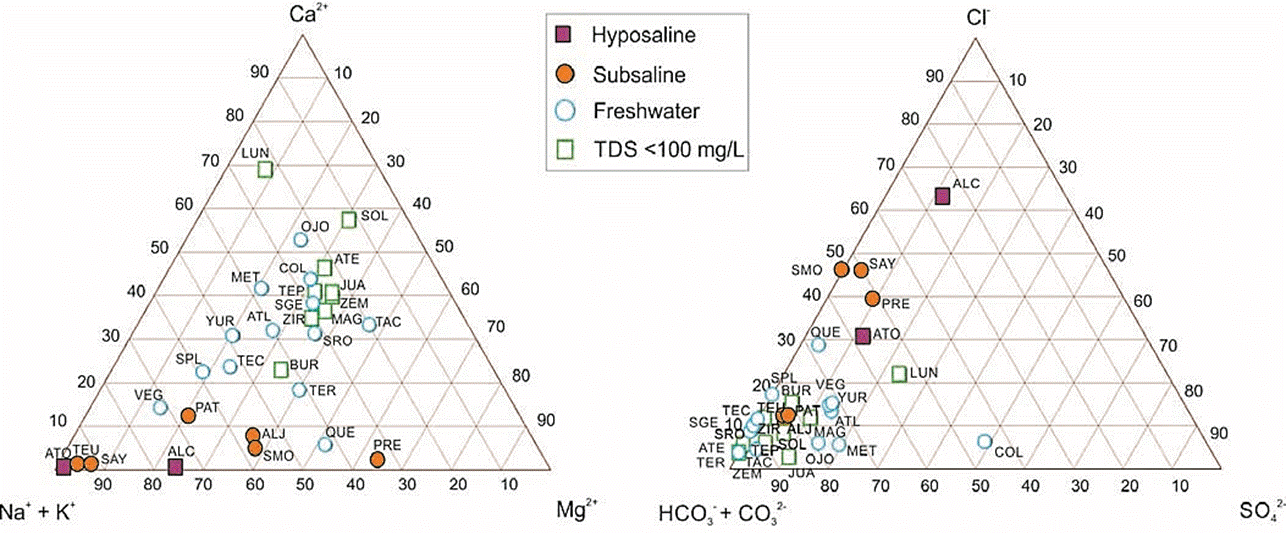
Figure 3 Triangular diagrams showing major cations and anions proportions in the 30 lakes studied along the Trans-Mexican Volcanic Belt, central Mexico. Abbreviations correspond to those in Table 1.
4.3. Nutrients and trophic state
According to the Redfield ratio (N:P = 16:1) half of the waterbodies could have their primary productivity limited by nitrogen (DIN:P-PO4 < 16:1) at the time of sampling, while the other half by phosphorus. However, several lakes had concentrations of these elements above the limiting values of 0.1 µM for P-PO4 and 7 µM for DIN proposed by Reynolds (Reynolds, 1999) and therefore their productivity could not be considered to be P or N limited (Figure 4A). Most of the lakes had also high concentrations of SiO2 (SiO2:P-PO4 < 16:1 and SiO2:DIN < 1:1, Xu et al., 2008, Figure 4B). Following these primary productivity nutrient limitation criteria (Redfield, 1958; Reynolds, 1999; Xu et al., 2008) 13 lakes could have some kind of primary productivity nutrient limitation at the time of sampling. Phosphorus limitation was the most frequent (8 lakes), followed by nitrogen (6) and silica (5); some of these lakes were co-limited by more than one of these nutrients: one lake was co-limited by nitrogen, phosphorus and silica (Zirahuén); two by phosphorus and nitrogen (Alberca de Tacámbaro and El Sol) and one by nitrogen and silica (Alchichica) (Figure 4). Atotonilco and Sayula were notable for having very high DIN and P-PO4 concentrations but a relatively low trophic level (both were mesotrophic), suggesting that productivity in these shallow lakes could be limited by other (unknown) factors.
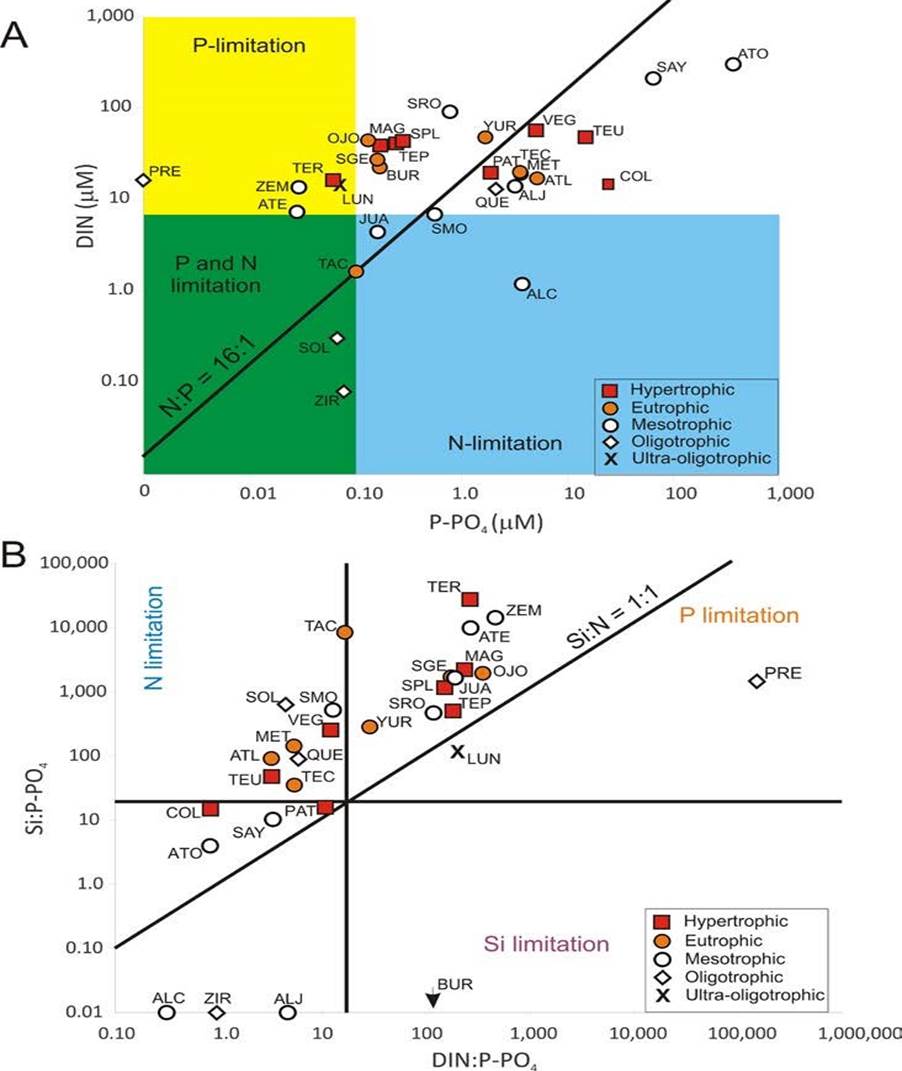
Figure 4 Nutrients in surface waters from the 30 lakes studied along the Trans-Mexican Volcanic Belt, central Mexico. A) P-PO4 and DIN molar concentrations diagram, diagonal line represents the Redfield (1958) N:P 16:1 ratio, shaded areas define nutrient limiting values according to Reynolds (1999) (P-PO4 < 0.1 µM; DIN < 7 µM). B) Nutrient molar ratios showing areas of Si, N or P limitation according to Xu et al. (2008), diatom growth could be limited by Si availability if Si:P<16 and/or Si:N<1. Abbreviations correspond to those in Table 1.
Most of the lakes where nutrient levels showed possible limitation of primary productivity were mesotrophic (9), oligotrophic (4) or ultraoligotrohpic (1) and nearly half of the studied lakes (16) were either eutrohpic (8) or hypertrophic (8). It is worth noticing that in general the eutrophic to hypertrophic lakes were shallow (< 8 m) and/or had low relative depth (< 1 %). This raises the question of whether this high trophic level is a natural feature of the shallow lakes in the TMVB or if it is a response to antropogenic impact. Besides, shallow water bodies are probably the most sensitive to climatic changes, therefore this kind of lakes should be monitored regularly (at least once a year), to record changes in temperature, pH, salinity, major ions and nutrient levels.
4.4. Bioindicators
We report the distribution of 97 taxa of aquatic bioindicators: 40 corresponding to phytoplankton, 30 to diatoms in surface sediments, 8 testate amoebae, 11 cladocerans and 8 ostracodes (see Electronic Supplement 2).
Considering that only the most abundant taxa in each lake are reported in this work, we found that more than half of them (65) were abundant only at one site; this applies particularly to the phytoplankton and sedimentary diatom taxa. Amongst the phytoplankton, Peridinium Ehrenberg had the highest frequency of occurrence (abundant in 5 lakes) while in the sedimentary diatoms it was Aulacoseria granulata (Ehrenberg) Simonsen (abundant in 6 lakes). Testate amoeba, cladocerans and ostracoda had a higher number of taxa common to more than one lake and, taxa like Centropyxis aculeata (Ehrenberg) Stein, Alona quadrangularis (O.F. Müller), Bosmina longirostris (O.F. Müller), Daphnia longispina- group (O.F. Müller), Candona Baird and Limnocythere Brady had relatively high occurrence frequencies (abundant in ≥ 6 lakes). Surprisingly some of the lakes that shared these high frequency taxa were located at opposite extremes in the TMVB; for example La Preciosa (east of TMVB) and Atotonilco (west of TMVB, Figure 1). However, this is only a first approach on the distribution of these bioindicators, and detailed, species level identifications supported by molecular studies would be necessary.
4.5. Climatic and environmental gradients
The PCA performed using geographical as well as climatic and limnological data (Figure 4A) showed a close correlation between both sets of variables. The longest environmental vectors, which correlate closely with the first axis (PC1 27.17 %), were mean annual temperature and water temperature. Altitude on the other hand correlates negatively with PC1 and with temperature, as higher altitudes are associated with cooler climates. The second axis (PC2 20.39 %) showed a positive correlation with precipitation and a negative correlation with total dissolved solids and electric conductivity. This relationship between lower precipitation and higher water ionic concentration has already been identified in previous works (Alcocer and Hammer, 1998; Armienta et al., 2008; Hernández-Avilés et al., 2007) and is associated with the process of evaporative concentration that lake water undergoes in relatively dry climates, where evaporation exceeds precipitation (Hardie and Eugster, 1970).
Following these environmental gradients (temperature and precipitation) the PCA biplot (PC1 vs. PC2) separates the studied lakes into four quadrants (Q) according to climate and limnological conditions (Figure 5). Cool, wet climates are associated with the lakes with the lowest TDS (< 100 mg/l) in QII (e.g. La Luna and El Sol). Lakes in cool and relatively dry climates are in QIII (e.g. Alchichica, Aljojuca, Quechulac) and those in warmer and relatively dry climates are in QIV (Atotonilco, Sayula), both groups of lakes showing the highest TDS and EC values. Finally, lakes in warm, wet climates, with medium TDS are located in QI (Colorada, La Vega, San Pedro Lagunillas, Tepetiltic). Taking into account the chlorophyll a vector, another ecological gradient becomes evident, with the eutrophic or hypertrophic lakes located in QI (moist and warm climates) while the (ultra)oligotrophic lakes are located in QII or QIII (cooler climates). High trophic levels in our data set seem to be at least partially related with climatic conditions, to warmer and moister climates where chemical weathering is intense and favors faster mobilization of chemical elements, including nutrients (N, P, Si) from the basement rocks into the lakes (Lewis, 1996; Malmaeus et al., 2006; Xia et al., 2015).
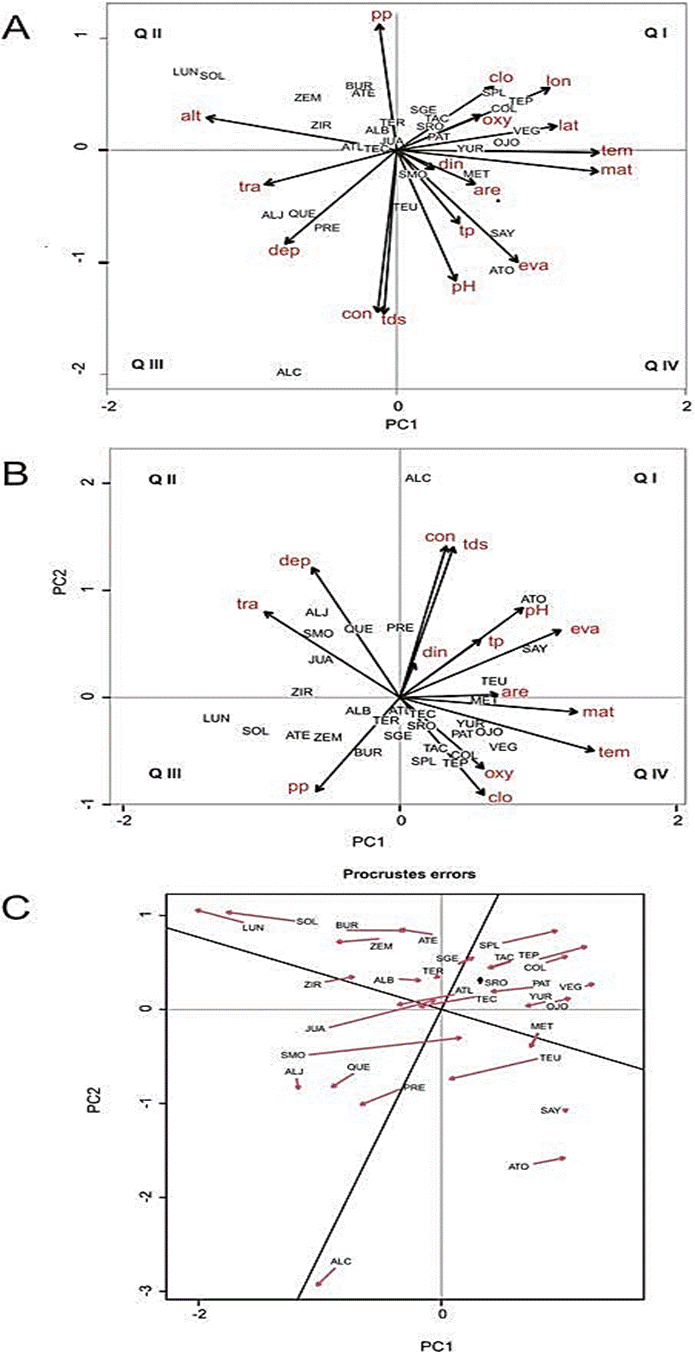
Figure 5 Principal component analysis (PCA) ordination of the 30 lakes studied along the Trans- Mexican Volcanic Belt, central Mexico; arrows indicate the geographic, climatic and limnological variables. A) PCA including latitude, longitude and altitude. B) PCA without latitude, longitude and altitude. C) Procrustes analysis showing the displacement of the sites between both ordinations. Abbreviations of waterbodies correspond to those in the chart of each waterbody (Figure 2 - 3). Abbreviations of environmental variables: alt, altitude; are, area; clo, chlorophyll a; con, electric conductivity; dep, maximum depth; din, dissolved inorganic nitrogen; eva, mean annual evaporation; lat, latitude; lon, longitude; mat, mean annual temperature; oxy, dissolved oxygen; pH, pH; pp, mean annual precipitation; tds, total dissolved solids; tem, water temperature; tp, total phosphorus; tra, transparency. Abbreviations correspond to those in Table 1.
It is interesting to note that in the PC1 vs. PC2 graph the lakes from opposite sides of the TMVB lie at adjacent quadrants (QIII and QIV) as they share high electric conductivities and total dissolved solids, associated to relatively dry climatic conditions. The bioindicator distribution previously discussed could be explained, therefore, as geographically distant lakes such as La Preciosa and Atotonilco share some physicochemical characteristics.
When the PCA is performed excluding geographical location (latitude, longitude and altitude) (Figure 4B) the variables associated to each axis (PC1 25.32 %, PC2 23.69 %) do not change, even though the PC2 orientation does. The Procrustes test allows us to determine which lakes are influenced the most by their geographic location, results are represented by arrows with their base indicating the location of the site in the first ordination, whereas the tip of the arrow indicates the localization of the same site in the second ordination. The lakes with the longest arrows (Figure 5C, e.g. Santa María del Oro, Juanacatlán and Tecuitlapa) are the ones that are more influenced by regional variables associated with their location (latitude, longitude and altitude) and would be the best choices for palaeoclimatic research while the shorter arrows (Alberca de Teremendo, Sayula, Santa Rosa, La Vega) indicate lakes which characteristics depend more on their local rather than regional variables.
It is only through the knowledge of the basic limnology and biodiversity of our lakes, generated via unified methodologies that guarantee comparable datasets, that more complex or detail studies can be proposed and developed leading to a better understanding, management and conservation of our country’s water resources.

