











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
Familial hypercholesterolemia (FH) is a monogenic, autosomal dominant disease, associated with variants in the low-density lipoprotein receptor (LDLR), apolipoprotein B (APOB), and proprotein convertase subtilisin/kexin type 9 (PCSK9) genes1.
Patients who are carriers this pathology often are asymptomatic, with low-density lipoprotein cholesterol (LDL-C) values > 200 mg/dL, or have early cardiovascular events (< 50 years in males and < 60 years in females). Characteristics of their inheritance allow identifying affected relatives and multiple members in a single family with high levels of LDL-C or cardiovascular events2.
Initial diagnosis is based on different clinical criteria that assess the personal and family cardiovascular history and characteristic clinical stigmas, such as tendon xanthomas and early corneal arcus, and finally, LDL-C plasma values. The Dutch Lipid Clinics Network (DLCN) criteria are the most widely used. A value > 8 points qualifies the patient as “definite” for FH diagnosis3.
Considering that inheritance is autosomal dominant and that the heterozygous form of FH is an effect of a single monogenic variant in one of the three FH-related primary genes at the following proportions: LDLR, 95%; APOB, 3%; and PCSK9, 1%, in addition to increased availability of access to modern sequencing techniques, genetic diagnosis is a central tool in the FH diagnosis of FH4.
After verifying the absence of a variation or point mutation in any of these three genes, it is imperative ruling out the presence of so-called major rearrangements, essentially duplications or deletions, with molecular biology techniques or bioinformatics, which cannot be detected by gene sequencing. Different reports show that 60-80% of patients with a definite FH diagnosis by clinical criteria have FH-related genetic variants4-7.
In turn, verification of the presence of a variant in FH-linked genes allows cascade screening in the rest of the family and better patient characterization for prognosis and treatment8.
In patients with a phenotype consistent with FH and with no detected variants, polygenic causes explain most cases. Different groups of researchers have developed calculations to obtain a score based on single nucleotide polymorphisms (SNP) related to increased circulating LDL-C values5,9. Finally, pointing out other potential causes of cases phenotypically consistent with FH without genetic findings is possible, including rare mutations in other minor genes related to lipid metabolism, such as apolipoprotein E gene (APOE), ABCG5, ABCG8, lipase A gene (LIPA), CYP27A1 or STAP1, and mutations or variants that are inaccessible with exome sequencing10.
The purpose of this work is to evaluate of a group of patients with FH phenotype and definite diagnosis and negative genetic result, after complete and expanded biomolecular analysis.
Materials and methods
Two-hundred and forty-six patients with hypercholesterolemia who were included during 2016 in the FH Detection Program in Argentina (Da Vinci Study), carried out at Partido de General Pueyrredón, Provincia de Buenos Aires, were assessed. The participants had cholesterol levels > 300 mg/dL and/or LDL-C > 190 mg/dL and granted informed consent for clinical, biochemical, and genetic evaluation. The study was approved by the Ethics Committee, Res CD FFyB Res CD 1762/17.
From each patient, family and personal history was obtained, with special focus on cardiovascular atherosclerotic disease (angina pectoris, myocardial infarction, myocardial revascularization surgery, stroke, and/or peripheral or carotid vascular disease), hospitalizations, hypertension, diabetes mellitus, and current lipid-lowering treatment. Careful physical examination, with special interest in the detection of tendon xanthomas or corneal arcus was carried out. When available, these data were obtained from first-degree relatives (parents, children, and siblings). Premature cardiovascular disease was regarded as the presence of any of the previously mentioned atherosclerotic cardiovascular disease manifestations, before 55 years of age for men and 65 for women. The DLCN criteria were applied, considering the highest LDL-C recorded value3. A total of 40% of patients received lipid-lowering treatment, and when LDL-C data off medication were not available, an LDL-C correction was applied based on the dose and potency of the statin being used11.
Twenty-one subjects were not included due to the following exclusion criteria: n = 8 had a thyroid-stimulating hormone (TSH) value > 10 mIU/L, four with creatinine > 1.7 mg/dL, two with alkaline phosphatase > 200 U/L, one patient with triglycerides > 1000 mg/dL, three women were excluded due to pregnancy, and three patients who refused to having a blood sample taken. Therefore, 225 subjects with potential FH were finally included.
The blood samples were collected in a non-fasting state and sent to the Lipid and Atherosclerosis Laboratory of the University of Buenos Aires for biochemical analysis. Serum total cholesterol, triglycerides, high-density lipoprotein cholesterol (HDL-C), direct LDL-C, creatinine, and alkaline phosphatase were determined using standardized enzymatic methods (Roche Diagnostics, Mannheim, Germany) in a Cobas C-501 autoanalyzer. Average intra-analysis coefficient of variation (CV) for all parameters was < 2.3% and inter-analysis CV < 3.0%. Apoprotein B was determined by immunoturbidimetric method in the same autoanalyzer, with an inter-analysis CV < 2.5%. TSH was measured by chemiluminescence (DPC, Immulite, Los Angeles, CA, USA) with intra-analysis and inter-analysis CV < 3.5%.
In all subjects with a DLCN score > 8, genetic study was carried out. Patient DNA was extracted from the whole blood sample at the Centro Nacional de Genética Médica12. The genetic study was carried out at Boston Heart Diagnosis, Massachusetts, USA, using next-generation sequencing, as described in a previous study13, to identify genetic variants in classical genes related to elevated LDL-C: LDLR, APOB, PCSK9, and in other genes associated with dyslipidemia: LDLRAP1, STAP1, ABCG5, ABCG8, APOE, LIPA, CYP27A1, LIPC, LIPG, and DHCR24. In addition, a panel of ten common variants (SNP) was studied in the following genes: rs6544713 in ABCG8, rs515135 in APOB, rs12740374 in CELSR2, rs3846663 in HMGCR, rs2650000 in HNF1, rs6511720 in LDLR, rs6102059 in MAFB, rs10401969 in NCAN, rs11206510 in PCSK9, and rs1501908 in TIMD4. Based on the detected SNPs, the genetic risk score (GRS) was calculated to assess the polygenic contribution to increased LDL-C14. Finally, bioinformatics analysis of the copy number variations (CNV) was carried out to identify large rearrangements in the structure of genes that are not detected by genetic sequencing15.
Data are expressed as median (range) according to their distribution and percentage of prevalence. Statistical analysis was carried out with SPSS 19.0.
Results
Twenty-one of the 225 studied patients with hypercholesterolemia had a DLCN score > 8, with an interval of 9 to 14. Out of these 21 patients with definite FH clinical diagnosis, two had first-degree relatives with cardiovascular disease, median (range) LDL-C of 250 mg/dL (181-431), which corresponded to the highest value eventually recorded or corrected at the end for the received treatment. Nine patients had arcus corneal arcus before 45 years of age; eight, tendon xanthomas; three, premature coronary heart disease; and two, early cerebral arterial disease. The rest of the lipid profile showed a median (range) of triglycerides: 121 mg/dL (62-314); HDL-C: 50 mg/dL (36-68); and APOB: 164 mg/dL (91-278).
Ten of the 21 patients with definite FH phenotype showed variants in LDLR and one in APOB, together with a variant in APOE, which accounted for 52.4% of patients who have point variants in classical genes. The study of the expanded panel of genes in patients with definite phenotype also revealed one case with mutation in LIPC, one with variant in LIPA, and another in CYP27A1. Table 1 shows the detected variants, as well as their classification with regard to their pathogenicity.
Table 1 Identified genetic variants. Clinical-biochemical, and polygenic score and phenotypic characteristics of each case
| DLCN | Genetic variant | Zygosity | Gene | Classification* | Polygenic score | LDL-C | Xanthoma | Arcus cornealis | Age | Gender |
|---|---|---|---|---|---|---|---|---|---|---|
| 9 | c.977C > G (p.S326C) | het | LDLR | VUS | No | 311 | No | No | 40 | F |
| 14 | c.646T > C (p.C216R) | het | LDLR | LP | No | 401 | Yes | No | 49 | F |
| 9 | c.1027G > A (p.G343S) | het | LDLR | P | No | 260 | No | Yes | 53 | F |
| 9 | c.1567G > A (p.V523M) | het | LDLR | LP | No | 251 | No | Yes | 52 | F |
| 9 | c.1895A > T (p.N632I) | het | LDLR | LP | Yes | 244 | No | Yes | 61 | M |
| 10 | c.1027G > A (p.G343S) | het | LDLR | P | No | 222 | No | No | 49 | M |
| 10 | c.1567G > A (p.V523M) | het | LDLR | LP | No | 273 | No | Yes | 36 | F |
| 11 | c.971A > G (p.D492G) | het | LDLR | LP | Yes | 321 | No | Yes | 52 | F |
| 11 | c.1118G > A (p.G373D) | het | LDLR | P | No | 251 | No | No | 47 | F |
| 11 | c.2312-?_2583+? del (p.781_?*del) | het | LDLR | P | No | 400 | No | No | 35 | F |
| 14 | c.428G > T (p.C143F) | het | LDLR | LP | No | 361 | Yes | No | 66 | F |
| 9 | c.68-?_1186+? dup (pV23_V395dup) (;) c.1141C > G (p.L381V) | het (;) het | LDLR (;)LIPA | P (;) VUS | No | 407 | Yes | Yes | 59 | F |
| 10 | c.10163C > T (p.T3388I) (;) c.968T > C (p.L323P) | het (;) het | APOB (;) APOE | LP (;) LP | Yes | 258 | No | No | 45 | F |
| 12 | c.720C > T (p.F240F) | het | CYP27A1 | VUS | Yes | 213 | No | Yes | 55 | F |
| 10 | c.724G > A (p.D242N) | het | LIPC | VUS | Yes | 288 | Yes | No | 66 | F |
| 9 | No mutation | - | - | - | No | 237 | No | Yes | 50 | F |
| 9 | No mutation | - | - | - | No | 221 | Yes | No | 49 | F |
| 9 | No mutation | - | - | - | No | 224 | Yes | Yes | 50 | F |
| 9 | No mutation | - | - | - | No | 300 | Yes | Yes | 53 | F |
| 9 | No mutation | - | - | - | No | 221 | Yes | Yes | 54 | F |
| 10 | No mutation | - | - | - | No | 209 | No | No | 61 | F |
*Classification according to the ACMG 2015 criteria for genetic variants in anomalies with monogenic Mendelian inheritance.
DLCN: Dutch lipid clinic network criteria; het: heterozygosity; VUS: variant of uncertain significance; P: pathogenic variant; LP: pathogenic type variant; F: female; M: male.
The CNV a posteriori bioinformatics study to detect large rearrangements in the gene structure showed that one of the cases with a negative genetic study and the carrier of the genetic variant in the LIPA gene were carriers of a deletion of exons 16-18 and one duplication of exons 2-8, respectively, in the LDLR gene (Table 1).
Regarding the calculation of GRS, five of the 21 patients had a higher GRS at the cutoff value of 1.96 established at the 90th percentile, although these cases were also carriers of variants in LDLR (n = 2) and cases with variants in APOB/APOE, CYP27A1 and LIPC. It should be noted that 6 (28%) of the 21 patients with a DLCN score > 8 did not show any type of genetic alteration after the extended genetic study and the detection of polygenic causes (Fig. 1).
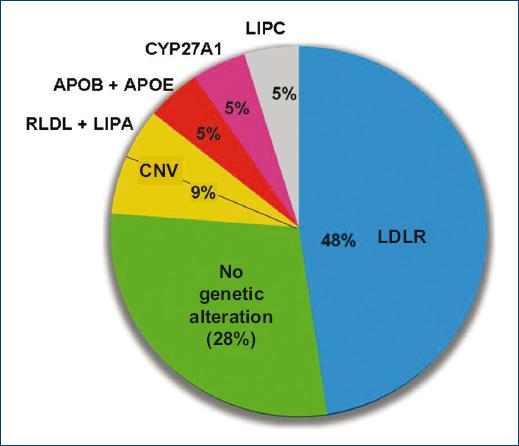
Figure 1 Distribution of the genetic variants detected in the 21 patients with a DLCN score > 8. CNV: copy number variations; LDLR: lipoprotein receptor gene; LIPC: lipase C gene; CYP27A1: cytochrome P450 family 27 type A1 gene; APOB: apolipoprotein b gene; APOE: apolipoprotein E gene; LIPA: lipase A gene.
Discussion
FH is a monogenic disease with high penetrance, classically associated with variants in the LDLR, APOB, and PCSK9 genes1. Current wide availability of genetic studies, in part due to cost reduction, has allowed deepening the study of patients with FH phenotype and establishing a correlation between clinical suspicion and the results of the sequencing of genes potentially participating in cholesterol metabolism.
After FH is clinically suspected, the procedure that follows is the application of a clinical score or diagnostic criteria to establish FH clinical diagnosis. The most widely used clinical-biochemical criteria are DLCN, where, based on five items, it is concluded if the patient has a definite (> 8 points), probable (6-8), possible (3-5), or improbable diagnosis (< 3)3.
Genetic analysis, looking for the genetic variant, is the next step, provided there are the resources and instruments necessary to carry out the sequencing15.
Identification of the genetic variant allows to better characterize the patient; the different mutations guide about disease prognosis; and it is possible to carry out a family cascade screening and, in selected cases, support specific therapeutics16,17.
During the development of the Da Vinci Study (Estudio Argentino Prevalencia en Hipercolesterolemia Familiar), 246 patients with increased levels of cholesterol and/or LDL-C were assessed, of which 21 had a DLCN score > 8, i.e., a FH definite diagnosis. These patients were studied with genetic sequencing for the detection of monogenic variants, with an expanded panel of 23 genes, added to an analysis of large rearrangements; finally, a polygenic score (PGS) of 10 SNPs related to LDL-C increase was applied. Of the 21 patients, 10 had variants in LDLR, one in APOB together with APOE, one in LIPC, and one in CYP27A1; they had also an elevated PGS, and two patients had one deletion and one duplication (genetic rearrangements) in LDLR, the latter with one variant in LIPA.
It should be noted that six of the 21 patients with a score > 8 showed no genetic alterations. Therefore, this 28% of patients comprises the category of subjects with FH phenotype and negative genetic study, keeping in mind that, in the present study, the evaluated gene panel was expanded and included polygenic causes, which further adds value to the finding18.
Possible explanations for this finding would be the following: (a) existence of variations or mutations in genes not yet recognized as causative of FH; (b) presence of mutations in regions not detected by currently available technologies, such as mutations mapped in deep intronic or promoter distal zones; (c) effects of the environment on genes (epigenetic phenomena), and (d) interactions between different genes in a single individual19-21.
On the other hand, DLCN score validation in this population for clinical diagnosis, as well as GRS, is necessary to confirm its usefulness in this setting and ensure its specificity and diagnostic sensitivity10,14.
Implementation of the expanded gene panel beyond those typically associated with FH has allowed recognizing unusual variants related to severe hypercholesterolemia13. In this study, one patient showed a variant in the lysosomal acid enzyme gene (LIPA), the deficiency of which has been linked to dyslipidemia and premature mortality22. This patient is a double heterozygote, since she also showed a duplication in LDLR; either way, the variant in LIPA is of uncertain significance. Other variants in uncommon genes that were identified in patients with a definite FH diagnosis were detected in the hepatic lipase gene (LIPC) and in CYP27A1 in association with cerebrotendinous xanthomatosis. However, those variants also coexisted with high GRS, which indicates polygenic causes. Therefore, the alteration mostly that causes elevated LDL-C remains to be elucidated, since polygenic causes or variants in unusual genes whose pathogenicity is still uncertain can coexist10.
Current usefulness of genetic studies for therapeutic decision-making is not established. It would be useful in cases of patients with homozygous FH, to quantify LDL receptor residual activity, but not in most FH cases. Therefore, LDL-c level and patient overall risk should be the determining factors regarding the therapeutic behavior in patients with FH18.20.
It can be concluded that, despite an exhaustive and expanded search for genetic causes in patients with a definite FH clinical diagnosis, 28% (6/21) of patients showed no genetic alteration. Future and more complex studies, such as performing exomes to different members of a single family, perhaps will finish demonstrating and explaining the referred findings.
Conclusion
Finally, in the analyzed cohort in a specific region of the Province of Buenos Aires, 28% of patients with a FH clinical diagnosis (DLCN) had no genetic alterations after having carried out an extensive analysis, not only of the variants in common genes related to FH but also after having studied genes considered non-classical and having applied a GRS to assess the polygenic contribution of elevated LDL-C and, finally, after a bioinformatics analysis to identify large rearrangements in the structure of genes not recognized by genetic sequencing.

