











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
Digitaria eriantha Steud cv. Survenola, named recently for the standardization of scientific writing1, was once known as Digitaria umfolozi and is popularly identified in Brazil as ‘faixa-branca’ or ‘pangolão’. It is a low-fertility hybrid plant obtained from the cross between D. setivalva Stent and D. valida Stent that has a tussock form of growth and is propagated via stolons. It is somewhat tolerant to low-rainfall periods by reason of its ability to capture dew, that condense together forming water droplets. The plant has a high regrowth capacity, which favors its use for grazing and harvesting2. It is also the most largely used grass species in northeast Brazil, notably in the states of Sergipe and Bahia, mainly because of its persistence in the soil and climatic conditions of that region, which has aroused the interest of producers, however, information on the production of this forage plant is still incipient, and the existing results often originate from regions of different environmental conditions3.
Net herbage accumulation in a pasture is the result of the balance between growth, senescence, and death of tissues - which are a consequence of the growth of new structures4 like leaves and stems on individual tillers - and of the number of existing tillers (density), associated with tillering patterns (appearance, mortality, and survival)5. However, other factors such as the amount and quality of light, temperature, water, and nutrients available in the local environment and the management strategy adopted interfere with photosynthesis and with the growth and senescence dynamics of a pasture6.
Increases in forage mass influence the leaf area index (LAI) and consequently light interception. Canopy height can be used as an indicator of the right time for defoliation, since LAIs above the ‘critical limit’ - when the canopy intercepts 95% of the incident light - favor stem and senescent-material accumulation7. However, adopting a fixed or pre-determined rest period for a grass species is not an effective management strategy to monitor the pasture growth and structure, given the physical and structural alterations the plant undergoes throughout the year.
In view of the above considerations, the present study was proposed to evaluate the herbage-accumulation dynamics of Digitaria eriantha cv. Survenola subjected to different canopy heights at two seasons of the year (rainy and dry).
Material and methods
The study was conducted in the Forage Plants Section of the Federal University of Sergipe (UFS), located in the municipality of São Cristóvão - SE, Brazil (10°55'53.7" S, longitude 37°06'18.8" W, 5 m asl). According to the Köppen classification, the climate in the region is a tropical Awa type. The average annual precipitation, temperature, and air relative humidity in São Cristóvão are 1,200 mm, 25.5 ºC, and 75 %, respectively8.
The National Institute of Meteorology (Instituto Nacional de Meteorologia) delivered the climatic data recorded during this study (Figures 1 and 2), where rainy and dry seasons are shown.
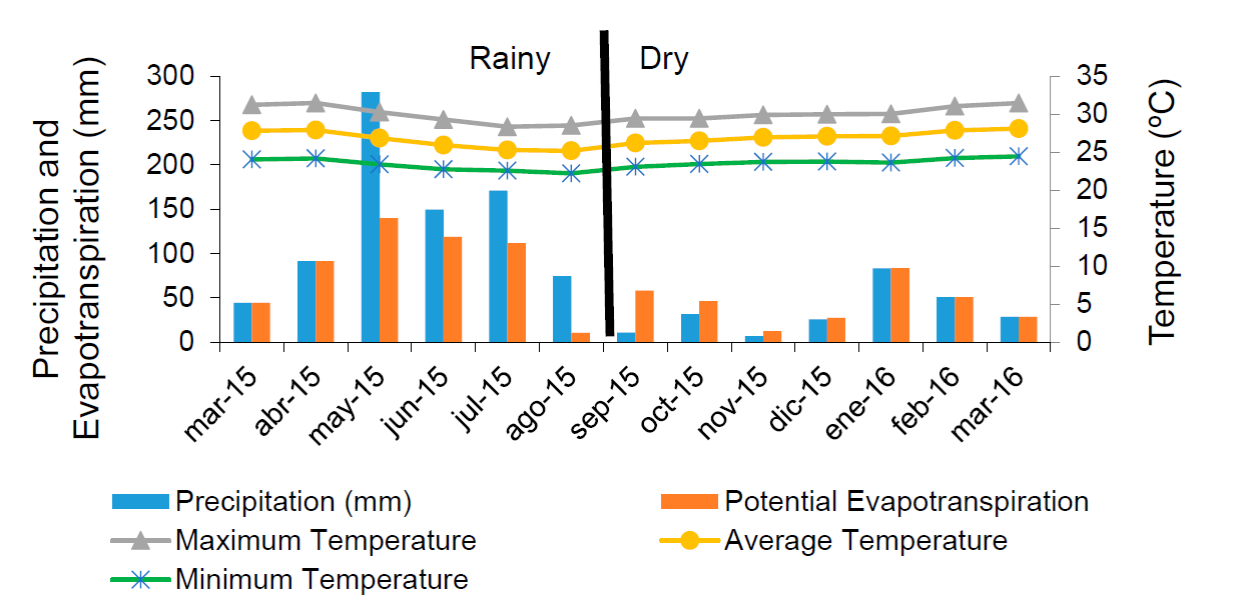
Figure 1 Precipitation, potential evapotranspiration and average, maximum and minimum temperatures during the experimental period9

The soil in the experimental area was classified as a Quartzipsamment of clayey texture10 with a flat relief. Before the experiment was established soil samples with a soil auger from three areas in each plot, at the 0-20 cm layer, and mixed to generate a composite sample. Subsequently, these were sent to the Technological and Research Institute of Sergipe State (Instituto Tecnológico e de Pesquisas do Estado de Sergipe - ITPS) for an analysis of chemical and particle-size properties that revealed the following composition: organic matter= 10.6 g.dm‒3; pH in H20= 6.15; P= 45.4 mg.dm‾3; Na= 0.053 cmolc.dm⁻3; Ca= 3.78 cmolc.dm‒3; Mg= 1.925 cmolc.dm‒3; Al= 0.08 cmolc.dm⁻3; H+Al= 0.67 cmolc.dm‒3; sum of bases= 5.77 cmolc.dm⁻3; CEC= 6.44 cmolc.dm‒3; base saturation= 89.60 %, and clay content 9.97 %.
According to the results of the soil analysis and in conformity with the recommendations of the Soil Fertility Committee of Minas Gerais State (Comissão de Fertilidade do Solo do Estado de Minas Gerais), no liming was required, since the base saturation content of the soil was 89.6 %. However, maintenance fertilization was necessary, and so 200 kg ha‒1 K2O in the form of potassium chloride were applied, split in three fractions11.
During the experimental period, fertilization was performed using 300 kg N ha‒1 yr‒1 in the form of ammonium sulfate. Because the harvest intervals depended on the time the plants reached the target canopy heights were imposed as treatments, different quantities of nitrogen fertilizer were applied after each harvest.
The grass was established in a total area of 90 m2 in June 2014. The area was divided into sixteen 5.2 m-2-experimental plots. In March 2015, a uniformity cut was made in all plots at 10 cm-stubble to start the study. Evaluations began on 03/26/2015 and ended on 03/28/2016, totaling 368 d of experimental period. Plots were irrigated with 5 mm of water the periods of water stress each two days due to the soil low water-retention capacity and the need for improving fertilizer utilization and maximizing herbage growth.
Treatments consisted of four canopy heights (25, 35, 45, and 55 cm) of harvest evaluated at two times of the year: rainy (March 26 to September 20, 2015), and dry seasons (September 21 to March 20, 2016). After every evaluation finished, were clipped the plants in all the experimental plots to a 10 cm of residual herbage. A randomized-complete-block experimental design was adopted, with four replicates, with 16 experimental units.
Canopy height was measured twice weekly, at five points per experimental unit, using a centimeter-graduated ruler12. Whenever the canopies reached the pre-established heights, was monitored the rest period (PD) (time required for the canopy to reach the established heights), their leaf area index and light interception using a SunScan® canopy analyzer (Delta Devices Ltd., Cambridge, United Kingdom) at three random points per experimental unit (plots). Evaluations corresponded to a reading performed above the canopy and another at the soil level (below the canopy).
The forage accumulation dynamics was evaluated weekly in five basal and five aerial tillers randomly selected and marked (after emergence) per experimental unit measuring the elongation of leaf blades and stems and senescence of leaf blades. On the last day of each evaluation, all marked tillers were cut at the soil level (basal tillers) or at the insertion point (aerial tillers), wrapped in a plastic bag, and immediately transported to a cold room to minimize moisture losses. With these data leaf and stem elongation rates and the leaf senescence rate (cm tillers‒1 day‒1) were estimated the, used to calculate the forage accumulation rate13.
Then, were dried the basal and aerial tillers in a forced-air oven at 65 ºC for 72 h and weighted them. Were calculated a length-mass conversion factor dividing the mass of each component by its respective growth, which was used to transform all field readings from cm tiller‒1 d‒1 into mg tiller‒1 d‒1. Finally, were multiplied these values by their respective tiller density in each experimental unit in order to compute the forage dry matter (DM) in kg ha‒1 d‒1.
Were assessed the live tillers density using 0.25-m2-metal-frame placed at random per experimental unit, always pre-harvest. Thus, the values for leaf and stem elongation rates and leaf senescence rate were transformed into leaf and stem growth rates and leaf senescence rate (kg ha‒1 d‒1 dry matter), respectively, for basal and aerial tillers. The sum of leaf growth and stem growth resulted in the total growth rate, whereas forage accumulation rate was calculated as the difference between total growth rate and leaf senescence rate14.
The variance analyzes were performed, considering a randomized block design with four replicates, and orthogonal decomposition of the sum of treatment squares into linear, quadratic, and cubic effects to probe the best fit of the model. The significance of effects was analyzed by the F test, at α=0.05, using the Computational Package Statistical Analysis System (SAS), version 9.0.
Results and discussion
Tiller density (TD) decreased linearly (P<0.05) for basal (Figure 3A) and aerial (Figure 3B) tillers in the dry season as the canopy heights increased. The same was not influenced (P>0.05) during the rainy season.
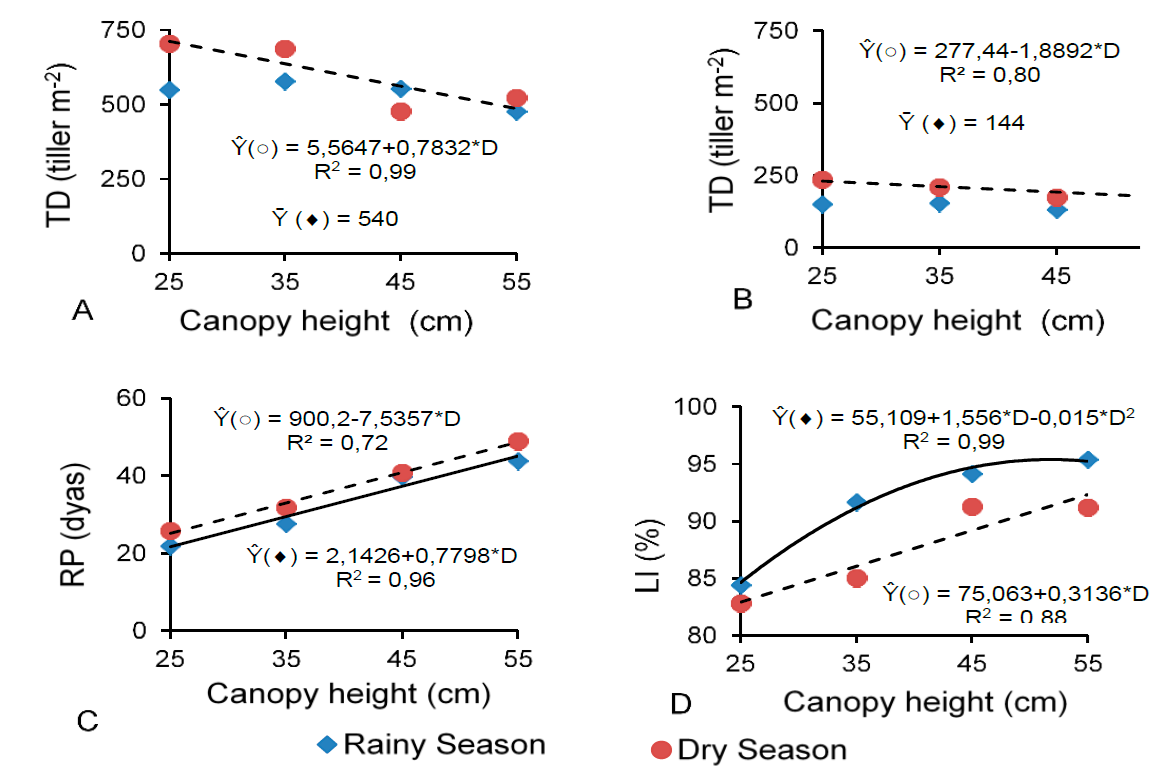
Figure 3 Tiller density (TD, A and B), rest period (RP; C) and light interception (LI; D) of basal (A) and aerial (B) tillers of digitgrass as a function of canopy heights during the rainy (() and dry (○) seasons
The rest period (Figure 3C) of the basal tillers, in the two seasons; and light interception (LI) (Figure 3D) in the dry period increased linearly (P<0.05) with the canopy heights. In the rainy season, this last variable responded quadratically (P<0.05) with a maximum LI value of 95.5 % at the canopy height of 55 cm (Figure 3D).
The reduction in the density of Basal (Figure 3A) and aerial (Figure 3B) tillers in the dry season, as the canopy heights increased can be explained by the longer rest period (Figure 3C); i.e., during the time to reach the target canopy heights, leaf production and stem elongation enlarged, which initially contributed to increasing light interception (Figure 3D), but ultimately reducing the incidence of light under the canopy. Reduced incidence of light at the base of the canopy caused the decrease of tiller density, as the activation of basal and axillary buds for the production of new tillers was inhibited15, as the rest time was longer for the greater canopy heights. In an experiment with Marandu Palisadegrass, longer harvest intervals negatively affected tiller densities, which was possibly due to the amount and quality of light reaching the canopy16. The reduction of light at the base of the lawn with increasing pasture height, influences the reduction of the population density of tillers17.
In the dry period, the intervals between harvests for the canopy to reach the pre-defined heights was on average 11.5 % longer than in the rainy period. Even though the plots were irrigated, evapotranspiration (Figure 1) and limited soil water retention capacity may explain these results; i.e., the amount of water was not sufficient to provide the ideal plant-growth conditions. In the rainy season, the climatic conditions (e.g., higher precipitation (Figure 1) and solar radiation of 1197.3 kj m2 (Figure 2) positively influenced plant growth. These results are likely due to the higher photosynthesis rates, which may culminate in elevated production of photo assimilates, thereby providing better conditions for plant growth. In an evaluation of the structural traits of Tanzania grass, more leaves had to be produced for the canopy to reach 95 or 97 % of LI, so grass plants could intercept more light and required longer rest periods15. The longer the canopy needs to attain a pre-established height, the larger the forage accumulation will be, especially under favorable conditions, such as higher precipitation, temperature, and light. However, this larger forage mass may result from the growth of stem and dead material, since leaf production increase is stabilized and stem growth and senescence processes are accelerated7.
Leaf growth rate (LGR) in basal (Figure 4A) and aerial (Figure 4B) tillers was not influenced (P>0.05) by the increasing canopy heights during the rainy and dry seasons. However, the leaf senescence rate of basal (Figure 4C) and aerial (Figure 4D) tillers, and the stem growth rate (SGR) of basal (Figure 4E) and aerial (Figure 4F) tillers in both seasons increased linearly (P<0.05) as canopy heights were greater.
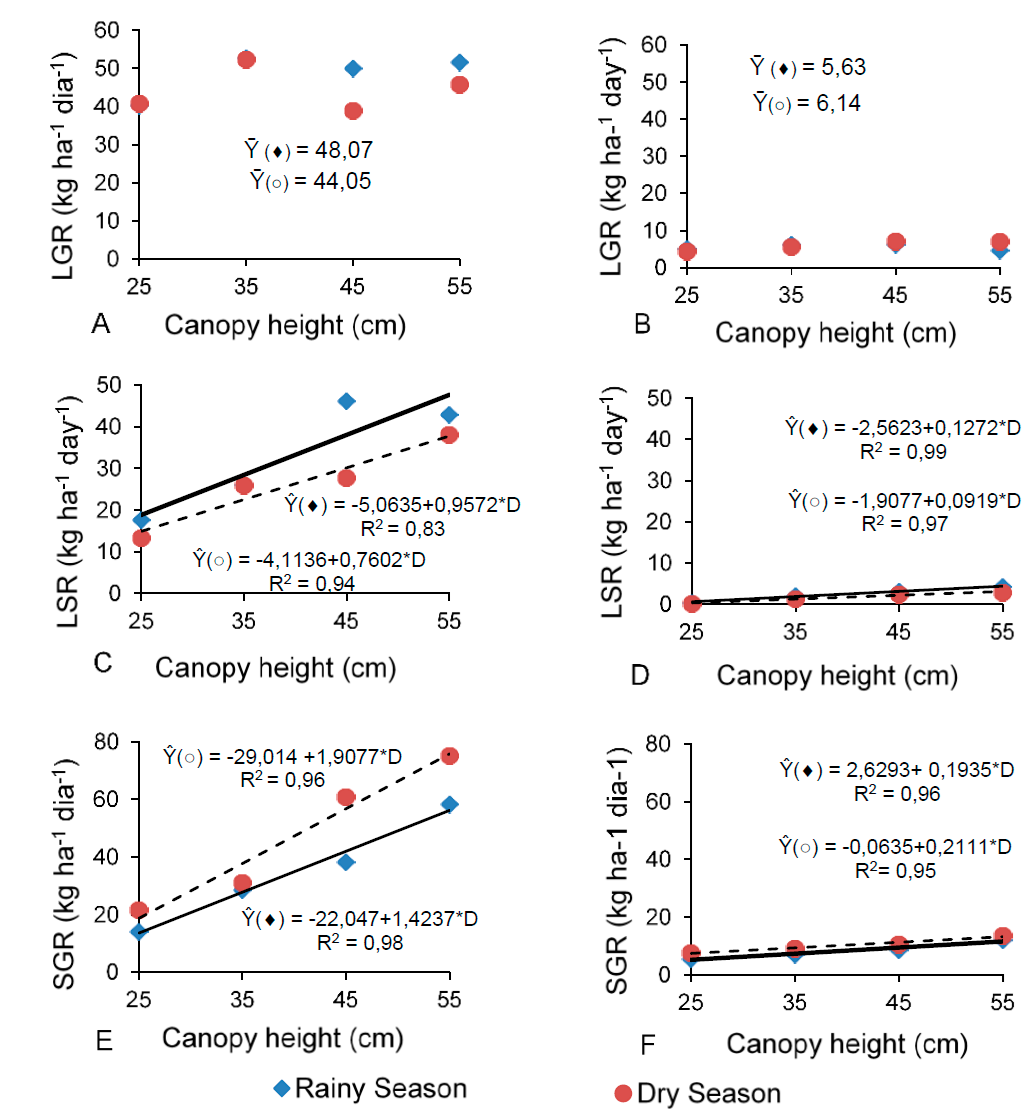
Figure 4 Leaf growth (LGR, A and B), leaf senescence (LSR, C and D) and stem growth rates (SGR, E and F) of basal (A,C and E) and aerial (B, D and F) tillers of digitgrass as a function of canopy heights during the rainy (() and dry (○) seasons
As a result, because of its longer rest period (Figure 3C), the highest canopy height (55 cm) would reduce the grass productivity, since no record higher daily growth rates under this treatment. The results for leaf senescence rate (Figures 4C and D) may be a consequence of the longer rest periods (Figure 3C), which led the leaves to reach their maximum lifespan and increase senescence rate. Higher canopy heights granted further plant development, as a consequence, more leaves completed their lifespan and senescent tissue accumulated 18.
The senescence rates of basal (Figure 4C) and aerial (Figure 4D) tillers while raining were 43 % and 26 % higher, on average, than in the dry season, respectively. This is explained by the higher rainfall (Figure 1) in the wet period. Solar radiation and temperature in this last season (Figure 2) averaged 1197.3 kj m2 and 27.8 ºC, in that order. Despite the irrigation during the months with sparser precipitation, evapotranspiration (Figure 1) might have influenced these results.
The increasing canopy height provided a longer rest period (Figure 3C), and favored the stem and dead material to build up, which was likely due to the shading of basal leaves. At the greatest canopy heights, stem elongation might occur as an attempt to rise the younger leaves in the upper stratum of the pasture, where canopy receives more photosynthetically active radiation. Increasing stem elongation and growth rates should reduce the pasture quality, because stems are the most fibrous fraction of forage, restricting its digestibility15. The shading of leaves at the base of the canopy may reduce their photosynthetic efficiency and tiller density in a mechanism known as population size/density compensation, in addition to elevating the accumulation of old tillers, which have leaf appearance and elongation rates reduced and senescence rate increased19.
Total growth rate (TGR) (Figure 5A) of basal tillers rose linearly (P<0.05) at both seasons, but the net herbage accumulation rate (NHAR) (Figure 5C) just did it during the dry season (P<0.05) as canopy heights were greater. About aerial tillers, the TGR (Figure 5B) improved during the dry season, but remained with no change during the rainy season.
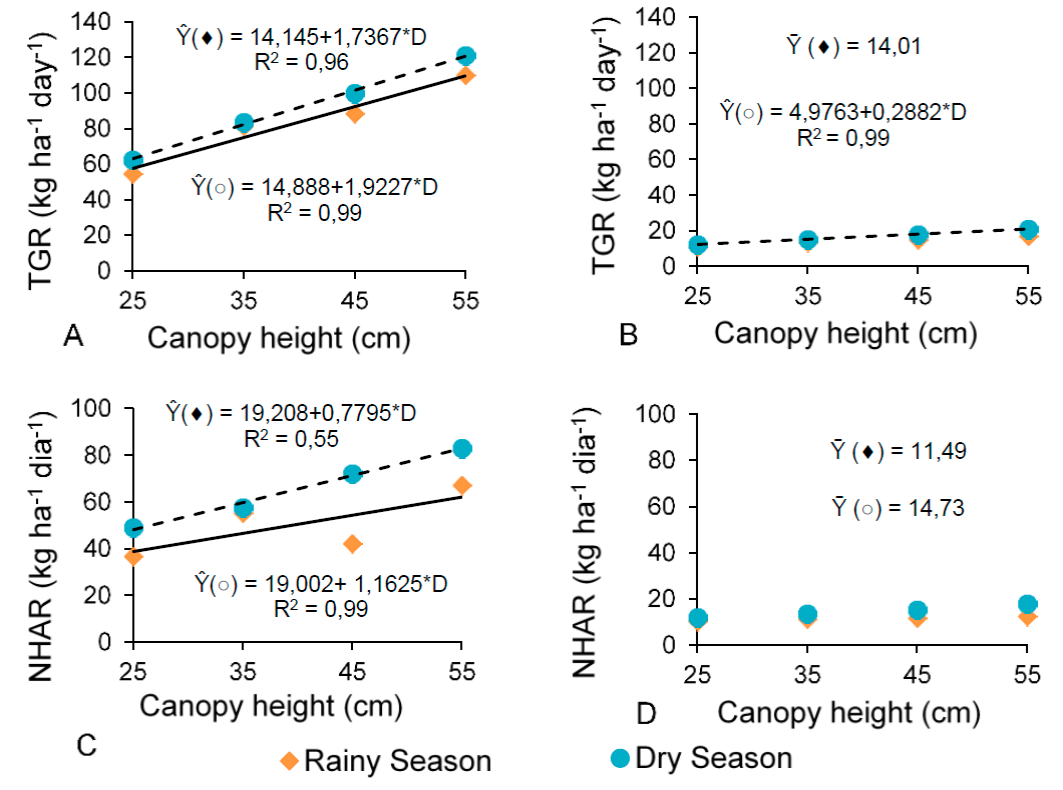
Figure 5 Total growth rate (TGR; A and B) and net herbage accumulation rate (NHAR; C and D) in basal (A and C) and aerial (B and D) tillers of digitgrass as a function of defoliation heights during the rainy (() and dry (○) seasons
NHAR (Figure 5D) did not change at the two seasons (P>0.05) because of the canopy heights. The population density of basal and aerial tillers (Figures 3A and 3B), leaf senescence rate (Figures 4C and 4D) and stem growth rate (Figures 4E and 4F) influenced, at different magnitudes, the TGR and NHAR of basal and aerial tillers during the dry and rainy seasons.
Because the canopy heights did not influence leaf growth rate (Figures 4A and 4B), then, the stem growth rate (Figure 4E and 4F) and hence possibly the tiller size:density ratio17 may explain the higher total growth rate (Figures 5A and B) and consequently NHAR (Figure 5C) as canopy heights increased. Results probed that the density of basal (Figure 4A) and aerial (Figure 4B) tillers in the dry season declined, as the canopy heights were greater; whereas was did not find any influence in the rainy season. However, the increasing canopy height may require heavier tillers to support its structure, which influenced the stem growth (Figures 4 E and 4F) and senescence rates (Figures 4C and 4D). Other authors found that seasons of the year and canopy heights also modified the tiller size:density ratio: Decreasing tiller densities are compensated by an increase in tiller weight, which results from higher stem- and leaf-elongation rates20. They observed that seasonal variations influenced the results. In an study with Tanzania grass subjected to three grazing intervals (90, 95, and 100 % LI) and two defoliation intensities (25 and 50 cm height), long rest periods caused changes in the canopy structure, with greater contribution of stem and dead material21. In this experiment, the increasing canopy heights elicited responses similar to those reported in this last study, with larger proportions of stem and greater senescence.
At the lowest canopy height (25 cm), results indicated an increase in the density of basal (Figure 3A) and aerial (Figure 3B) tillers during the dry season. For basal and aerial tillers in both seasons, however, there was a decrease in the rest period (Figure 3C), light interception percentage (Figure 3D), leaf senescence rate (Figures 4C and 4D), and stem growth rate (Figures 4E and 4F). On the other hand, TGR (Figures 5A and 5B) for basal tillers in the two seasons and aerial tillers in the dry season was lower at the 25-cm-canopy height. The same was true for NHAR (Figure 5C) of basal tillers in both seasons. Based on these results, it is suggested that, at the lowest height, senescence and stem elongation were lessened, implying a better forage nutritional quality. Nevertheless, higher defoliation frequencies, with shorter rest periods, may deplete the energy reserve of grass plants for canopy growth and, at the end, reduce the persistence of this grass over time. The greatest defoliation height (55 cm) provided the lowest tiller density (Figures 3A and 3B) for basal and aerial tillers in the dry season; and the highest leaf senescence rate (Figures 4C and 4D) and stem growth rate (Figures 4E and 4F) for basal and aerial tillers in the two periods, which may negatively influence the herbage digestibility. A longer rest period may bring about an increase in stem and senescent-material growth rates, which may reduce the pasture quality15. Therefore, letting the canopy to reach between 35 and 45 cm can benefit herbage accumulation as well as provide better quality to digit grass, and ultimately improving its productivity. Depending on the time of the year and growth conditions, a shorter rest period with lower defoliation heights may lead to yield losses, whereas longer rest periods may induce nutritional and quantity losses. Moreover, both cases can result in pasture degradation7.
As previously discussed, the tiller size:density ratio influenced the results; stem elongation was higher possibly because the rest period to reach the greater canopy heights was longer. Other research probed that the weight of tillers in the vegetative stage is lower than in the reproductive stage for Brachiaria decumbens, which also bore inflorescences22. The same authors reported that taller plants had heavier vegetative tillers, but a lower tiller density. These findings can explain the present results, where the increasing canopy heights provided an increase in TGR and NHAR. In the dry season, the vegetative tillers changed to a reproductive stage that resulted in higher stem growth rates. In an evaluation of the nutritional value of tillers from Brachiaria decumbens cv. Basilisk plants, researchers found a greater stem elongation rate when tillers progressed from the vegetative to the reproductive state, contributing to a lower leaf:stem ratio23.
Conclusions and implications
Digit grass should be managed in a defoliation-height interval of 35 to 45 cm, because when defoliated above this range, it reaches a greater accumulation of forage, to the detriment of greater accumulation of stem and dead material. Further studies should be undertaken focusing on the residual height with animals involved in the evaluation procedures, under different environmental conditions.

