











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
Treeline ecotones represent the highest point at which an arboreal stratum can be found (Körner, 1998; Lutz, Larson, Freund, Swanson, & Bible, 2013); they are conspicuous zones of transition between forest ecosystems and peatlands, high moorland or alpine grasslands (Camarero & Gutiérrez, 2004; Körner, 1998). Since extreme low temperatures are the main regulator of physiological and ecological processes of trees at high altitudes (Tranquillini, 1964), these ecotones are considered highly sensitive to climate change (Holtmeier & Broll, 2005; Körner, 1998). In this context, the presence of trees of various species (e. g., Larix sibirica Ledeb., Pinus uncinata Raymond ex A. DC., P. hartwegii Lindl.) above their altitudinal limits (enclaves) is associated with their need to find similar environmental conditions to their original niche (Camarero & Gutiérrez, 2004; Holtmeier & Broll, 2005; Jobbágy & Jackson, 2000). Therefore, altitudinal migration, in addition to adaptation and extinction, is one possible response of alpine vegetation to environmental temperature increase (Aitken, Yeaman, Holliday, Wang, & Curtis-McLane, 2008). This response will depend on species and the extent to which its distribution is controlled by temperature (Jobbágy & Jackson, 2000). However, if the ecotone is not able to migrate at the same rate as the species on an individual basis, the rate of movement will decrease and even migration will be inhibited in its entirety, so the complexity of the ecotone could function as a barrier to altitudinal migration (Lutz et al., 2013).
In general, the treeline ecotone presents additional stress conditions to those present in the forest core (Viveros-Viveros et al., 2009). Low temperatures reduce the activity of apical meristems and new tissue formation (Tranquillini, 1964), generating trees with structural deformation or krummholz (Camarero & Gutiérrez, 2004; Körner, 1998) or reducing seed production, germination and seedling survival (Coop & Givnish, 2008), which modifies the structure of treeline ecotones. In this respect, based on the dominant regulatory mechanism in the ecotone, Harsch and Bader (2011) classify treeline ecotones in four types: diffuse, abrupt, island and krummholz. Diffuse type, the most common of the four, is characterized by growth limitations (Harsch & Bader, 2011); the abrupt ecotones are formed due to an increase in seedling mortality, while dieback produces islands or groups of trees isolated from each other (Bader, Rietkerk, & Bregt, 2007; Harsch, Hulme, McGlone, & Duncan, 2009). Shape of the trees can also be altered due to the incidence of intense winds or snow presence on trees, producing krummholz (Körner, 1998). Therefore, tree height and density are key attributes to understand structure and processes that regulate establishment in treeline ecotones (Bader et al., 2007), and the local factors that could limit altitudinal migration of forests, in the climate change context (Harsch & Bader, 2011).
Pinus hartwegii is the highest-altitude (2,800 to 4,200 m) pine species in the world (Perry, 1991). This species forms monospecific forests that form ecotones with high mountain grasslands (Challenger & Soberón, 2008); it presents great adaptation to extreme low temperatures down to -30 °C (Perry, 1991), supporting snowfall during the months of December to February (Challenger & Soberón, 2008). However, the adaptation of this species to low temperatures also makes it one of the most vulnerable to climate change (Ricker, Gutierrez-García, & Daly 2007).
In Mexico, changes in altitudinal distribution of P. hartwegii in Izta-Popo National Park (Beaman, 1962), Nevado de Toluca (Lauer, 1973) and Cofre de Perote (Iglesias & Tivo, 2006), associated with changes in local temperature, have been reported. However, such reports are focused on P. hartwegii as a species, without considering the structure, composition and dynamics of the ecotone as a whole (Harsch & Bader, 2011). Based on the above, it is important to know P. hartwegii structure through the transition zone to the alpine grassland established at higher altitude and under more adverse environmental conditions. Therefore, the objectives of the present study were to characterize the structure of P. hartwegii treeline ecotone at Nevado de Toluca and determine if any changes in its altitudinal limits occurred during the last 25 years. This study was conducted with the expectation that it could serve as a baseline to monitor potential effect of climate change on altitudinal distribution of P. hartwegii.
Materials and methods
Study area
The study was carried out in Nevado de Toluca Flora and Fauna Protection Area (APFF for its initials in Spanish) in the State of Mexico, between the Toluca and Tenango valleys (Figure 1). Study area is located between 18° 51' - 19° 19' NL and 99° 38' - 100° 09' WL, with an altitudinal range between 3,000 and 4,680 m (Körner & Paulsen, 2004). The semicold-subhumid C(E)wig and cold E(T)Hwig climates predominate (García, 1990); mean annual temperature ranges between -2 and 7 °C and mean annual precipitation between 200 and 1,800 mm, with snowfall in the months of December to February (Challenger & Soberón, 2008). The dominant soil group is Andosol, with Phaeozem, Regosol, Cambisol and Leptosol groups occurring to a lesser extent (Körner & Paulsen, 2004). The main ecosystems are temperate forests consisting of species of Abies, Pinus and Quercus, located at an altitude of 3,000 to 4,100 m, and high mountain grassland composed of Festuca and Calamagrostis, at altitudes of 4,100 to 4,500 m (Calderón de Rzedowski & Rzedowski, 2005).

Species under study
Pinus hartwegii is a pine adapted to extreme low temperatures that is distributed on the tops of the highest mountains, from Nuevo León in northeastern Mexico to El Salvador (Calderón de Rzedowski & Rzedowski, 2005; Perry, 1991), at elevations of 3,000 to 4,200 m (Beaman, 1962; Lauer, 1973; Perry, 1991). Its natural distribution is discontinuous and restricted to high-elevation sites such as Nevado de Colima, Pico de Tancítaro, Nevado de Toluca, Ajusco, Popocatépetl, Iztaccíhuatl, Malinche and Pico de Orizaba, in the Trans-Mexican Volcanic Belt, where it constitutes the treeline (Calderón de Rzedowski & Rzedowski, 2005; Vera-Vilchis & Rodríguez-Trejo, 2007). This species is of great ecological importance due to its limited geographical distribution and its tolerance to extreme low temperatures, which can reach as low as -30 °C (Beaman, 1962; Perry, 1991).
Sampling design
In this study stratified-systematic sampling (Matteucci & Colma, 1982) was used. Treeline was taken as a starting point and five perpendicular transects of 220 m in length were established from there. Two transects were located in a northwest direction and three in an east-southeast direction with respect to the volcanic crater. Accessibility, state of treeline conservation and slope similarity were considered to establish transects. In each transect six plots of 20 m x 20 m were formed based on the model of Camarero and Fortin (2006); three plots were located below treeline (forest area) and three above it (grassland area), with a distance of 40 m between the center of two adjacent plots, as shown in Figure 2. Plots located at the same altitudinal level of a different transect were considered as repetitions in tree density calculations, while for the height and DBH (1.3 m above the ground) calculations, trees were considered individually.

Structure of treeline ecotone of P. hartwegii forest
The structure of P. hartwegii treeline ecotone was characterized in the months of August to October 2014, by recording height and DBH of each tree in the plots (Cárdenas, Martínez, Bravo, Núñez, & Caballero, 2008) and tree density in the altitudinal gradient (3,980 to 4,120 m). Dimension of trees was compared by establishing diameter classes by altitudinal level and identifying the strata that make up the ecotone. The percentage of proportional reduction in tree density between altitudinal levels was calculated in order to identify the form presented by the Nevado de Toluca treeline ecotone, according to Harsch and Bader (2011) classification. Forest regeneration was evaluated by counting and calculating the density of seedlings and individuals less than 1.3 m in height and 10 cm in DBH.
Altitudinal delimitation of P. hartwegii treeline ecotone
Upper and lower altitudinal limits between which the transition zone of the P. hartwegii forest to alpine grassland is distributed at Nevado de Toluca were determined by adapting the moving split window method of Camarero and Fortin (2006). This was done by taking as reference the pixels for the 2014 Landsat image, the spatial resolution of which was adjusted to 15 m by fusion with a panchromatic image of the same year. The tree density averages per pixel and the ecotone boundaries in the pixels that presented significant statistical differences were obtained with IDRISI Selva software (Clarck Labs, 2012). These boundaries were corroborated by the supervised classification of 30 georeferenced points within the ecotone and 30 points in each cover that forms the transition zone (forest and high mountain grassland), which were obtained in the field with an altimeter GPS (Garmín brand, model V, USA). Altitudinal limits were extrapolated to the whole ecotone on Nevado de Toluca; based on the delimitation, an analysis of altitudinal changes was made to determine the modifications in altitudinal distribution of P. hartwegii treeline ecotone in the last 25 years. Two Landsat images were used, one from 1989 and the other from 2014, adjusted to 15 m resolution, which were compared through the IDRISI Selva’s Land Change Modeler module (Clarck Labs, 2012).
Statistical analysis
Tree height, DBH and individual density were used as the main factors in a one-way analysis of variance (ANOVA), followed by a Tukey’s multiple comparison test. On the other hand, in order to establish the relationship between height and DBH of trees along the altitudinal gradient, a linear regression was applied for the following association of variables: height vs. DBH, height vs. altitude, DBH vs. altitude, height-DBH vs. altitude. The results of the ANOVA applied to the density of the trees were used to obtain the points in altitude between which the differences in density were statistically significant; these points were used as reference in the extrapolation of the altitudinal limits. All analyses were performed in JMP8 (Statistical Analysis System [SAS], 2008) with a confidence level of 95 %.
Results and discussion
Structure of P. hartwegii treeline ecotone
The P. hartwegii treeline ecotone of the Nevado de Toluca conforms to the diffuse form of the Harsch and Bader (2011) classification. This form describes the gradual reduction in tree height and density from the forest center to higher altitude.
The trees of P. hartwegii treeline ecotone ranged from 0.3 to 22.0 m in height with a mean of 3.88 m (SD = 4.5). Below the forest line (Bos1, Bos2 and Bos3), trees had an average height of 4.09 m (SD = 4.7), while above the line (Pas4, Pas5 and Pas6) the mean was 2.2 m (SD = 1.4). Trees at 4,043 m (Bos1) recorded the highest and lowest height.
Regarding DBH, trees measured between 1.0 and 72.0 cm, with a mean of 11.82 cm (SD = 11.85). Figure 3 shows that the largest diameters were recorded in the trees of the Bos1 (4,043 m) and Bos2 (4,050 m) levels. Below the forest line (Bos1, Bos2 and Bos3), trees measured between 1.0 and 72.0 cm in DBH, with a mean of 12.05 cm (SD = 12.3), while above the line (Pas4, Pas5 and Pas6) they measured between 1.5 and 26.5 cm, with a mean of 9.96 cm (SD = 5.63).
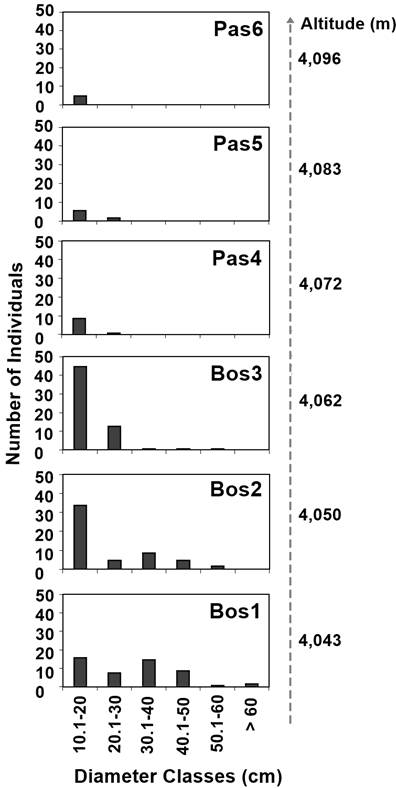
Figure 3 Number of Pinus hartwegii individuals per diameter classes in the altitudinal levels above (Pas4, Pas5 and Pas6) and below (Bos1, Bos2 and Bos3) the forest line.
Separation by diameter classes allowed identifying the height range of the regeneration trees (DBH ≤ 10 cm), which was between 0.3 and 4.0 m; for juvenile- and adult-size individuals (DBH > 10 cm), height ranged between 1.5 and 22 m. Both height (r2 = 0.144, SE = 4.23) and DBH (r2 = 0.06, SE = 11.39) presented significant negative relationships (P = 0.0001) with increased altitude.
According to Wong and Lertzman (2001), reduction in tree height and DBH along the altitudinal gradient does not necessarily imply a lower age in individuals. At high altitudes, the growth rate of P. hartwegii is lower due to limitations imposed by low temperatures (Tranquillini, 1964). At these sites, mature individuals (62 years old) can be found above 4,000 m with significantly smaller sizes compared to the forest core trees (Beaman, 1962); the great adaptation of this species to freezing temperatures also makes it vulnerable to changes in environmental temperature ranges, affecting growth. In this regard, Ricker et al. (2007) predict a 10.6 % reduction in growth rate of P. hartwegii associated with an increase of 0.6 °C in average environmental temperature in the next 100 years. In addition to the above, P. hartwegii can present a caespitose state during the first years; that is, its growth is reduced or practically zero, with this state being able to last up to seven years (González-Rosales & Rodríguez-Trejo, 2004). The above makes it even more difficult to determine the time of establishment and the relationship it might have with biotic and abiotic variables. Therefore, it is essential to develop and apply techniques to determine the precise age of individuals in the treeline ecotone and enclaves to elucidate the effect of increased environmental temperature on the altitudinal distribution of the species in question.
In terms of tree density, some authors (Holtmeier & Broll, 2005; Jobbágy & Jackson, 2000) point out that this variable has important effects on seed production and germination, as well as on the establishment and development of new individuals. According to Figure 4, along the treeline ecotone, tree density showed a gradual reduction between 8 and 70 % in the altitudinal levels. On the other hand, Figure 5 shows that the highest density was recorded in the forest area; the Bos1 altitudinal level (4,043 m) had the highest average with 37 trees per plot (925 trees·ha-1; SD = 37.3), while the Pas6 level (4.096 m) had the lowest density, being one tree per plot (25 trees·ha-1). Based on this reduction along the altitudinal gradient, it can be said that the P. hartwegii treeline ecotone corresponds to the diffuse form of the Harsch and Bader (2011) classification, being relevant that the diffuse ecotones have recorded altitudinal advances with the greatest frequency at the global level (Harsch & Bader, 2011; Holtmeier & Broll, 2005; Körner, 1998). This confirms that environmental conditions limit the growth of P. hartwegii above 3,980 m, resulting in a well-defined treeline (Harsch & Bader, 2011).
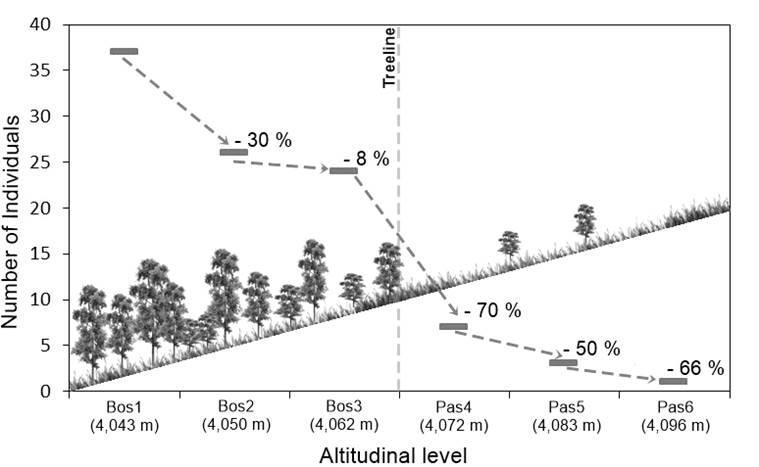
Figure 4 Proportional reduction in Pinus hartwegii tree density with respect to the immediately preceding altitudinal level.
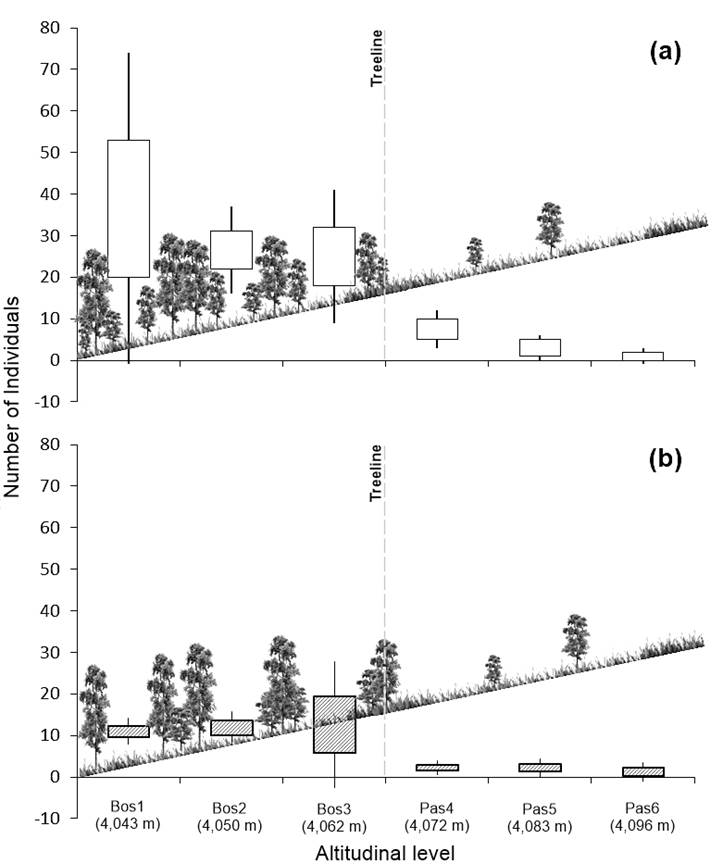
Figure 5 a) Total density of trees in the forest’s treeline ecotone and b) density of trees larger than 10 cm in DBH and 1.3 m in height.
A low density of adult-size individuals (300 to 700 trees·ha-1) favors tree regeneration (Camarero & Gutiérrez, 2004) because the presence of other trees mitigates the effects of extreme low temperatures and increases microsite availability (Coop & Givnish, 2008). Low density also provides sufficient space for the establishment and development of new individuals (Viveros-Viveros et al., 2009); by contrast, with a density of more than 1,000 trees·ha-1, development decreases considerably due to reduced survival capacity of the regeneration individuals (Vera-Vilchis & Rodríguez-Trejo, 2007). This is reflected in the presence of enclaves, which is considered an indicator of the altitudinal advance of P. hartwegii; however, it does not imply an advance of the forest itself, rather it represents the success of the more genetically resistant individuals, which were able to overcome the adverse conditions and find a favorable microsite for seedling establishment.
Under high mountain conditions, a higher density can even lower the soil temperature under the trees due to shading, reducing survival and development capacity of the root system and seedlings in general. In this regard, Körner and Paulsen (2004) reported that temperatures lower than 5 °C and higher than 12 °C reduce germination success and initial growth of P. hartwegii at Nevado de Toluca and Pico de Orizaba. Such a narrow optimal temperature range suggests that a minimal change in environmental temperature could significantly impact the population dynamics of the species. This demonstrates the need to know, in a more precise way, the effect of environmental conditions (temperature and soil characteristics) and the altitudinal gradient on the mechanisms that restrict the growth of P. hartwegii at the limit of its altitudinal distribution. This, in turn, will enable obtaining more precise predictions about the potential distribution of P. hartwegii in the long term and determining whether the species is actually able to migrate altitudinally as a result of increased environmental temperature.
Altitudinal delimitation of P. hartwegii treeline ecotone
Altitudinal limits of P. hartwegii treeline ecotone identified in the 1989 image were found in a range of 3,990 to 4,040 m as the lower limit and between 4,055 and 4,110 m as the upper limit. In 2014 images, the lower limit was identified between 3,980 and 4,090 m, and the upper between 4,030 and 4,130 m (Figure 6). These ranges are congruent with those reported 55 years ago by Beaman (1962) on Iztaccíhuatl (3,920 to 4,111 m) and Popocatépetl (3,753 to 4,076 m). These similarities show not only a behavior pattern of the species, but a convergence of conditions under which it is distributed and the effect of historical changes that environmental conditions have had on it (Jobbágy & Jackson, 2000; Körner & Paulsen, 2004). The upper limit identified for 2014 was 20 m higher than in 1989; however, no significant differences (P = 0.07) were found between two sampled years. No significant differences (P = 0.11) were found in the treeline altitude between the northwest and east-southeast exposures sampled in the field; the northern volcanic cone face recorded the highest altitudinal variation of treeline ecotone (Figure 6). The upward shift on the north face was between 25 and 60 linear meters, equivalent to an average increase of 6.5 m in altitude (SD = 4.5) on the upper boundary of treeline ecotone. On the other hand, the south side showed no visible changes in altitudinal distribution of the ecotone with the images used. It is important to consider that the growth rates of arboreal life forms, particularly when they develop in high-mountain altitudinal limits, are extremely slow. Therefore, the monitoring of species and their long-term altitudinal distribution, both in the ecotone and enclaves located above the transition zone, is fundamental to determine if there are regeneration processes associated with the maintenance of the ecotone, that is, if it is a stable or dynamic ecotone with constant ascents and descents (Harsch et al., 2009; Harsch & Bader, 2011).

Conclusions
At global level, the diffuse form in ecotones is considered the most frequent, with limitations on tree growth being dominant. The diffuse form of the P. hartwegii treeline ecotone reflects adverse conditions to which it is subjected. Considering that this type of ecotone can have constant altitudinal ascent and descent dynamics, this study provides a baseline for later comparisons. Since no differences were found in altitudinal distribution of the P. hartwegii ecotone between the periods evaluated, it is important to establish long-term monitoring to determine the dynamics of the species, and to know if it is actually able to migrate altitudinally as a result of an increase in environmental temperature. This would allow, at the time, the application of efficient adaptive management techniques.

