











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkHighlights:
Ecological niche modeling allows identifying areas to establish silvopastoral systems.
The ecological niche of Chloris gayana, Eragrostis curvula, Pinus devoniana, P. lawsonii, and P. montezumae was modeled.
Central and southern Mexico has a high potential for silvopastoral systems.
Agricultural areas have a larger potential area compared to forest areas.
Pinus devoniana had the greatest potential area to be intercropped with C. gayana and E. curvula.
Introduction
Silvopastoral systems consist of plots where forest and forage species are intercropped to produce feed for livestock and maintain tree cover (Mosquera-Losada, McAdam, Romero-Franco, Santiago-Freikanes, & Rigueiro-Rodríguez, 2009). These systems represent an alternative for the recovery of forest vocation of soils converted to agricultural land and provide ecosystem services such as shade for livestock, fruits, seeds, firewood, reduction of soil erosion, increased water infiltration to the subsoil, habitat for fauna, and microclimate regulation (Casanova-Lugo et al., 2015); on a regional scale, silvopastoral systems contribute to the capture of atmospheric pollutants, carbon storage, climate change mitigation, diversification of agricultural production and biodiversity recovery (Montagnini, Ibrahim, & Murgueitio-Restrepo, 2013).
Ecological niche models have been used to understand the natural distribution of species (McKenzie, Peterson, & Peterson, 2003), project the expansion of invasive species (Schussman, Geiger, Mau-Crimmins, & Ward, 2006), identify areas with high biodiversity (Newbold, Gilbert, Zalat, El-Gabbas, & Reader, 2009) and define priority areas for biological conservation (Hirzel, Hausser, Chessel, & Perrin, 2002). In the case of silvopastoral systems, the ideal areas to establish are identified through ecological niche models, because these can cartographically represent the optimal environment for the development of forage and forest species, which is delimited by filters such as temperature, precipitation, altitude, and soil type (Guisan & Zimmermann, 2000).
In the temperate region of Mexico, the establishment of silvopastoral systems is developing. The expansion of the agricultural frontier and soil management practices such as mechanized tillage, excess nitrogen fertilizer, and planting on steep slopes have caused physical and chemical degradation of soils in the center and south of the country (Cotler, Corona, & Galeana-Pizaña, 2020), especially in Estado de México, Mexico City, Michoacán, and Tlaxcala, states with the highest rates of water erosion (Bolaños-González et al., 2016). In this regard, silvopastoral systems with annual or perennial fast-growing forage species and intercropped trees can be a useful tool to stop soil erosion processes in agricultural areas (Pérez-Nieto, Valdés-Valverde, & Ordaz-Chaparro, 2012).
In forested areas of Mexico, a long-established practice is the introduction of livestock for grazing in forests or on sites with secondary vegetation; however, animals can negatively impact the flora and soil (Fuentes-Hernández et al., 2019). These impacts on the forest ecosystem can be minimized through silvopastoral systems, by rotating livestock in natural forests and planting forage species preferred by these animals (Ávila-Ramírez et al., 2019).
Chloris gayana Kunth (Rhodes grass) and Eragrostis curvula (Schrad.) Nees (weeping love grass) are two forage species native to Africa, introduced in various parts of the world for livestock feeding (palatability and high production value) and soil erosion control (Ponsens, Hanson, Shcellberg, & Moeseler, 2010). These species are adapted to various environments, including humid tropical, arid, and temperate areas (Imaz, Giménez, Grimoldi, & Striker, 2012). Both species are present in Mexico, as forage crops, along roadsides, and on disturbed lands (Valdés-Reyna 2015). Such forages have shown great potential for establishment in silvopastoral systems with timber species in some temperate regions of Mexico (Sáenz-Reyes et al., 2019).
The potential of silvopastoral systems with C. gayana and E. curvula in temperate areas of Mexico could be increased through the use of commercially important conifer species, widely propagated in forest nurseries and with soil retention and erosion prevention capacity, such as Pinus devoniana Lindley (Michoacán pine), Pinus lawsonii Roezl ex Gordon (Lawson’s pine) and Pinus montezumae Lambert (Montezuma pine) (Gómez-Romero et al., 2012; Sáenz-Reyes, Muñoz-Flores, & Rueda-Sánchez, 2011). Therefore, the objectives of this study were to model the distribution area of C. gayana, E. curvula, P. devoniana, P. lawsonii, and P. montezumae in Mexico and identify potential areas for the establishment of silvopastoral systems with these species in forest and agricultural areas of the country.
Materials and methods
Forage component
C. gayana is a perennial grass (Figure 1a), develops canes that measure between 50 to 130 cm in height, grows in the rainy season, flowers all year round, and is present in 22 states of Mexico (Valdés-Reyna, 2015). The species adapts well to loamy and saline soils, its optimal development is achieved in subtropical and tropical climates and it is shade tolerant when established intercropped with trees (Guevara-Escobar, Cervantes-Jiménez, Suzán-Azpiri, González-Sosa, & Saavedra, 2012). This grass is used for direct grazing and responds to chemical fertilization with diammonium phosphate and urea, organic fertilizers, liquid biofertilizers, and vermicompost (Sáenz-Reyes et al., 2016), achieving yields between 5 and 15 Mg∙ha-1 of dry matter in soils of volcanic origin in central Mexico (Imaz et al., 2012).
E. curvula is a perennial grass (Figure 1b), it develops deep roots and tufted canes that measure between 60 and 150 cm in height; it grows in the spring-summer cycle, flowers in summer and resprouts abundantly after being defoliated by livestock; it adapts well to clay loam, sandy loam soils with low humidity; it responds to chemical fertilization with N, K, Mg, Si, NH4NO3 and organic fertilization with manure, fishmeal, and lime (Ward, Kirkman, & Tsvuura, 2017). The species is drought resistant, adapts to temperate climates and is shade tolerant (Ghebrehiwot, Fynn, Morris, & Kirkman, 2006); it can reach up to 15 % crude protein content in the spring-summer cycle, it produces yields of 4 Mg∙ha-1 in sandy soils of western Mexico (Esqueda-Coronado & Carrillo-Romo, 2012) and is present in 10 states of the country (Valdés-Reyna, 2015).
Forest component
P. devoniana (Figure 1c) is a species distributed mainly in the Neovolcanic Axis and Sierra Madre del Sur. Trees can reach 20 to 30 m in height (Farjon, Pérez de la Rosa, & Styles, 1997); they grow in deep, volcanic, sandy-loam soils and form monospecific stands or stands associated with other pine or oak species. This species is used for timber, resin extraction, and pulp for paper (Sáenz-Reyes et al., 2011).
P. lawsonii (Figure 1d) is a conifer restricted to the Neovolcanic Axis and Sierra Madre del Sur. Trees reach up to 30 m in height (Farjon et al., 1997); this species grows in deep, sandy soils with a silty-clay texture and acid pH. This species is used as firewood, for making fence posts, beams, and rustic furniture, and sometimes for resin production (Ramos-Uvilla et al., 2014).
P. montezumae (Figure 1e) is distributed in the Neovolcanic Axis, Sierra Madre del Sur and small portions of the Sierra Madre Oriental. Trees reach 20 to 35 m in height (Farjon et al., 1997). This species grows in well-drained volcanic soils with a clay-loam texture, forming monospecific stands or in co-dominance with other pine and Abies species; it is mainly used for timber production and has been widely propagated for commercial forestry plantations (Centre for Agricultural Bioscience International [CABI], 2002; Pérez-Miranda, Moreno-Sánchez, González-Hernández, & Arriola-Padilla, 2014).

Ecological niche modeling
The raster format layers of the 19 bioclimatic variables, at a spatial resolution of 1 km2, were obtained from the WorldClim portal (Hijmans, Cameron, Parra, Jones, & Jarvis, 2005); the altitude, slope, and slope orientation layers from the Hydro1k portal (Hydro1k Elevation Derivative Database) (United States Geological Survey [USGS], 2000); and the soil types from the Geoinformation Portal of the National Biodiversity Information System (INEGI, 2014). All layers were cut to the limits of the Mexican territory (Table 1).
Table 1 Bioclimatic and topographic variables used to model the ecological niche of Chloris gayana, Eragrostis curvula, Pinus devoniana, P. lawsonii, and P. montezumae in Mexico.
| Key | Variable |
|---|---|
| Bio01 | Mean anual temperature (°C) |
| Bio02 | Diurnal temperature range (°C) |
| Bio03 | Isothermality (°C) |
| Bio04 | Temperature seasonality (%) |
| Bio05 | Maximum temperature of the warmest period (°C) |
| Bio06 | Minimum temperature of the coldest period (°C) |
| Bio07 | Annual temperature range (°C) |
| Bio08 | Average temperature of the wettest quarter (°C) |
| Bio09 | Average temperature of the driest quarter (°C) |
| Bio10 | Average temperature of the warmest quarter (°C) |
| Bio11 | Average temperature of the coldest quarter (°C) |
| Bio12 | Average annual precipitation (mm) |
| Bio13 | Precipitation of the wettest period (mm) |
| Bio14 | Precipitation of the driest period (mm) |
| Bio15 | Precipitation seasonality (%) |
| Bio16 | Precipitation of the wettest quarter (mm) |
| Bio17 | Precipitation of the driest quarter (mm) |
| Bio18 | Precipitation of the warmest quarter (mm) |
| Bio19 | Precipitation of the coldest quarter (mm) |
| Altitude | Meters above sea level (m) |
| Slope | Degrees (°) |
| Hillside | North, South, West, East |
| Type of soil | FAO nomenclature |
The information on the presence of the study species in Mexican territory was downloaded from the Global Biodiversity Information Facility (GBIF, 2020) database. Data were cleaned by eliminating repeated points and making boxplots to identify outliers according to the bioclimatic and topographic variables evaluated. Principal component analysis (PCA) was then performed to identify the variables that most influence the distribution of collection points for each species, selecting those with the greatest magnitude and easiest to interpret. The boxplots and PCAs were run using the R-3.5.1 program (R-Core Team, 2017).
Once the most important variables for each species were identified, the ecological niche was modeled using Maxent 3.3.3 (Phillips, Anderson, & Schapire, 2006), using 75 % of the collection points for modeling and the rest as validation points. Models with area under the curve (AUC) values between 7 and 9 were considered acceptable, while those with values greater than 9 were considered to have a high predictive capacity (Peterson et al., 2011).
Furthermore, a partial ROC (Receiver Operating Characteristics) analysis was performed in the NicheToolBox program (Osorio-Olvera et al., 2020). This analysis minimizes omission and commission errors of the AUC, using 50 % of the collection points as validation data, 1 000 iterations, and a commission error of 5 %. Partial ROC values were validated by Z-tests to determine if they were different at random (ROC = 1); those close to 2 and statistically significant (P < 0.05) indicated models that fit the actual species distribution very well (Slater & Michel, 2012). The maps obtained for each species were reclassified, where cells with a probability of the presence of species lower than the first quartile were discarded (Peterson et al., 2011).
Potential areas for the establishment of silvopastoral systems
Areas with water bodies and urban settlements were eliminated from the distribution maps of each species, according to the Land Use and Vegetation Map Series VI for Mexico, scale 1:250,000 (INEGI, 2017). Subsequently, maps were projected to the Lambert 1992 Conic coordinate system and the potential area of the distribution of each species (km2) was calculated.
The maps of C. gayana and E. curvula were superimposed with the maps of P. devoniana, P. lawsonii and P. montezumae to obtain the potential areas for the establishment of silvopastoral systems in forest areas, agricultural areas, and by the federal state, according to the distribution of primary forest and agricultural areas of the Land Use and Vegetation Map Series VI. Finally, estimated extent (ha) of potential silvopastoral areas with these species were calculated.
Results
Ecological niche of the study species
According to Table 2, the PCAs showed that altitude and annual temperature range (Bio07) were the most important variables for Pinus species. The third most important variable for P. devoniana was the average temperature of the warmest period (Bio05), for P. lawsonii, mean annual precipitation (Bio12), and for P. montezumae, mean annual temperature (Bio01). In general, C. gayana and E. curvula responded most to altitude, followed by annual temperature range (Bio07) in the case of E. curvula, and the average temperature of the wettest quarter (Bio08) for C. gayana; for both species, the variable average temperature of the coldest quarter (Bio11) was the third most important. Figure 2 shows the contribution of bioclimatic and topographic variables on species distribution.
Table 2 Contribution of environmental variables in the potential distribution models of two forage species and three pine species in Mexico.
| Environmental variable | Contribution to distribution models (%) | ||||
|---|---|---|---|---|---|
| Chloris gayana | Eragrostis curvula | Pinus devoniana | Pinus lawsonii | Pinus montezumae | |
| Altitude | 44.9 | 62.9 | 32.3 | 36.8 | 58.2 |
| Mean annual temperature (Bio01) | 0.1 | 0.1 | 2.3 | 6.2 | 6.8 |
| Isothermality (Bio03) | 1.7 | 2.7 | - | - | - |
| Maximum temperature of the warmest period (Bio05) | 1.2 | 0.1 | 15.9 | - | - |
| Annual temperature range (Bio07) | - | 27 | 39.7 | 35.3 | 31.2 |
| Average temperature of the wettest quarter (Bio08) | 34.8 | - | - | 5.3 | - |
| Average temperature of the coldest quarter (Bio11) | 9 | 5.4 | 8.1 | - | 1.8 |
| Average annual precipitation (Bio12) | 2.7 | 0.3 | - | 14.8 | - |
| Precipitation of the driest period (Bio14) | 5.7 | - | 2.4 | - | 2 |
| Precipitation of the driest quarter (Bi017) | - | 1.5 | - | 1.5 | - |
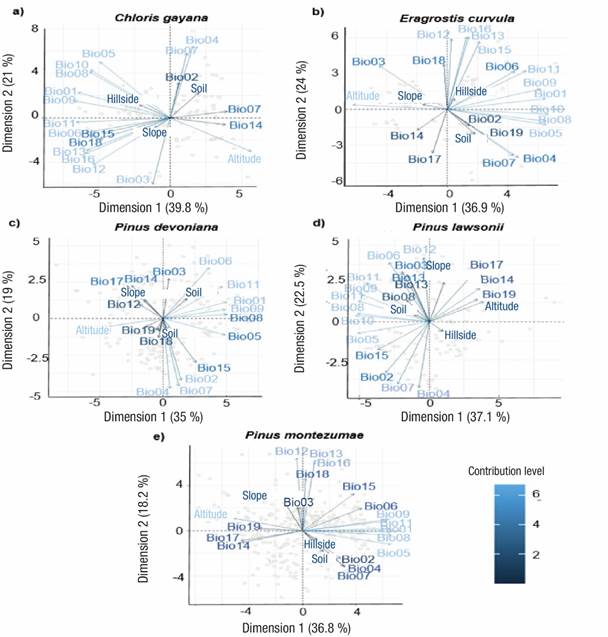
Figure 2 Principal component analysis showing the contribution of bioclimatic and topographic variables on the distribution of two forage species and three Pinus species in Mexico. The keys of the variables are described in Table 1.
Ecological niche models had high predictive capacity with AUC values greater than 0.93, while partial ROC values were very close to 2, as shown in Table 3. Species distribution areas were located at altitudes between 1 620 and 2 940 m, corresponding to temperate regions, with mean annual temperature between 12.2 and 19.5 °C and mean annual precipitation between 450 and 1 200 mm. The grass E. curvula occupied higher elevations and with lower mean annual temperature compared to C. gayana. On the other hand, P. montezumae was found on higher slopes and sites with lower mean annual temperature compared to P. devoniana and P. lawsonii, species that shared elevation and mean annual temperature ranges.
Table 3 Ecological niche values of species evaluated for the establishment of silvopastoral systems in Mexico.
| Descriptor | Chloris gayana | Eragrostis curvula | Pinus devoniana | Pinus lawsonii | Pinus montezumae |
|---|---|---|---|---|---|
| Area under the curve (AUC) | 0.9345 | 0.9418 | 0.9577 | 0.9901 | 0.9814 |
| Partial ROC | 1.8691 | 1.8836 | 1.9156 | 1.9802 | 1.9628 |
| Standard deviation | 0.0201 | 0.0415 | 0.0096 | 0.0017 | 0.0053 |
| Z value | 2 939.1 | 1 432.1 | 6 270.3 | 34 836 | 11 599 |
| P value | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 |
| Altitude (m) | 1 620-2 070 | 2 082-2 385 | 1 720-2 340 | 1 930-2 220 | 2 340-2 940 |
| Mean annual temperatura (°C) | 16.7-19.5 | 14.9-16.8 | 15.6-18.7 | 16.4-19.1 | 12.2-15.5 |
| Mean annual precipitation(mm) | 567-840 | 450-813 | 957-1 164 | 876-1 153 | 965-1 200 |
| Estimated surface área (km2) | 302 400 | 85 720 | 103 900 | 27 340 | 46 580 |
Figure 3 illustrates the potential distribution of the species evaluated. Chloris gayana was found in an area of 302 400 km2, occupying extensive areas of the Neovolcanic Axis, of the Mexican Plateau and some portions of the Sierra Madre Occidental, Sierra Madre Oriental, and Sierra Madre del Sur; while the distribution area of E. curvula was smaller, occupying 85 720 km2, especially located in the Neovolcanic Axis and the southern portion of the Mexican Plateau. In the case of conifers, P. devoniana occupied a larger area (103 900 km2) compared to the other two species along the Neovolcanic Axis, Sierra Madre del Sur, and the Chiapas highlands; P. montezumae (46 580 km2) was found mainly in the eastern portion of the Neovolcanic Axis and some portions of the Sierra Madre Oriental, Sierra Madre del Sur, and Chiapas; while P. lawsonii had a more restricted distribution (27 340 km2), mainly in the Neovolcanic Axis and Sierra Madre del Sur.

Potential areas for silvopastoral systems
According to Figure 4, potential areas were located mainly in the Neovolcanic Axis and Sierra Madre del Sur. With C. gayana and P. devoniana, potential areas in forest zones were found mostly in the western portion of the Neovolcanic Axis (4 730 024 ha), while in agricultural areas they were found in the eastern portion of this mountain range (1 559 539 ha). The distribution of P. lawsonii was more discontinuous with portions mainly in agricultural areas of Michoacán, Puebla, and Guerrero (635 154 ha), while potential areas with P. montezumae were concentrated in agricultural areas in the central states of the country (2 536 042 ha). On average, the area in forest zones represented 25 % of the total area for each species.
The distribution of areas suitable for silvopastoral systems with E. curvula and P. devoniana had a pattern similar to that of C. gayana, with areas of preserved forest in the western portion of the Neovolcanic Axis (676 321 ha), while agricultural areas were located in the eastern portion of this mountain range (2 761 068 ha). The areas for silvopastoral systems with P. lawsonii were smaller (128 344 ha), with small portions in agricultural areas of the Neovolcanic Axis. Suitable areas with P. montezumae were also concentrated in the Neovolcanic Axis, with forest areas (432 801 ha) in Jalisco, Michoacán, Guanajuato and Querétaro, while agricultural areas (1 228 405 ha) were located in Estado de México, Mexico City, Hidalgo, Tlaxcala and Puebla.
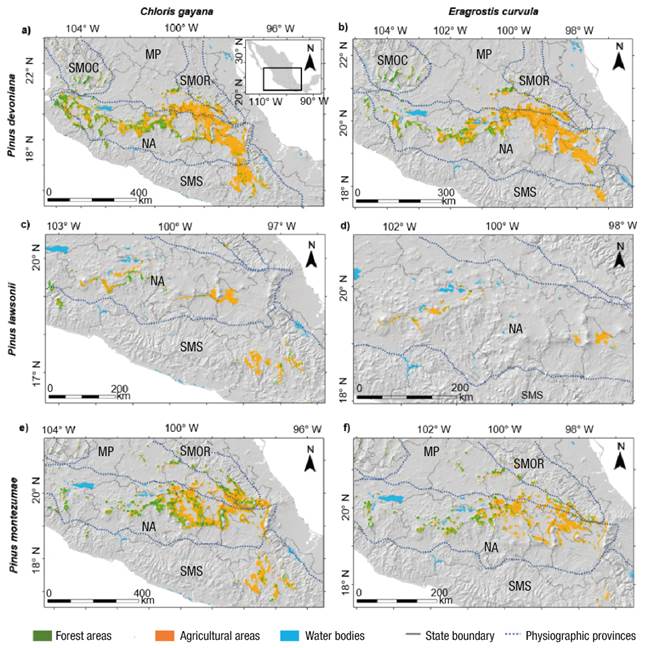
Figure 4 Potential areas for the establishment of silvopastoral systems of two forage species and three pine species in forest and agricultural areas of Mexico. Physiographic provinces: SMOC = Sierra Madre Occidental, SMOR = Sierra Madre Oriental, MP = Mexican Plateau, NA = Neovolcanic Axis, SMS = Sierra Madre del Sur.
As for the distribution by state, Table 4 shows that potential areas with C. gayana and at least one pine species were found in 20 states. For agricultural areas, 48.5 % of the suitable area with P. devoniana and 84.6 % of the area with P. lawsonii were concentrated in Michoacán, Puebla and Oaxaca, while 56.7 % of the area with P. montezumae was located in Puebla, Oaxaca and Estado de México. In the case of forested areas, Michoacán was first in the area with P. lawsonii and P. montezumae, and second with P. devoniana. Jalisco, Michoacán and Oaxaca concentrated 67.2 % of the area with P. devoniana and 87.4 % with P. lawsonii, while Michoacán, Oaxaca and Estado de México had 55.8 % of the area with P. montezumae.
Table 4 Potential area for silvopastoral systems with Chloris gayana and three pine species in Mexico.
| State | Pinus devoniana | Pinus lawsonii | Pinus montezumae | |||
|---|---|---|---|---|---|---|
| Agricultural area (ha) | Forest (ha) | Agricultural area (ha) | Forest (ha) | Agricultural area (ha) | Forest (ha) | |
| Chiapas | 67 370 | 21 800 | 8 737 | 1 433 | 66 040 | 22 070 |
| Coahuila | 236 | 21 | - | - | 15 590 | 6 020 |
| Colima | 8 037 | 6 633 | 255 | - | 2 292 | 2 288 |
| Mexico City | 35 840 | 4 550 | 416 | - | 56 470 | 12 790 |
| Durango | 8 241 | 30 169 | - | - | 679 | 7 051 |
| Guanajuato | 130 600 | 51 500 | 1 571 | 1 732 | 36 300 | 42 020 |
| Hidalgo | 684 400 | 50 600 | 6 161 | 1 392 | 286 500 | 88 500 |
| Jalisco | 508 900 | 549 100 | 2 970 | 18 620 | 24 990 | 80 310 |
| Estado de México | 523 600 | 79 000 | 52 700 | 7 770 | 660 300 | 212 900 |
| Michoacán | 734 800 | 402 200 | 157 500 | 91 200 | 226 600 | 257 500 |
| Morelos | 29 330 | 18 080 | 18 050 | 5 730 | 22 800 | 20 970 |
| Nayarit | 24 030 | 79 570 | - | - | 356 | 5 350 |
| Nuevo León | 5 063 | 8 897 | - | - | 40 070 | 24 680 |
| Oaxaca | 760 700 | 97 500 | 261 700 | 33 000 | 421 400 | 68 500 |
| Puebla | 797 600 | 61 600 | 118 100 | 2 100 | 355 800 | 110 700 |
| Querétaro | 166 900 | 35 800 | 3 191 | - | 62 850 | 31 150 |
| Sinaloa | 977 | 15 573 | - | - | - | - |
| Tlaxcala | 198 900 | 10 500 | - | - | 193 700 | 22 600 |
| Veracruz | 19 050 | 5 450 | - | - | 57 930 | 14 660 |
| Zacatecas | 21 940 | 28 030 | - | - | 1 763 | 8 477 |
| Total | 4 726 515 | 1 556 572 | 631 351 | 162 977 | 2 532 430 | 1 038 536 |
In the case of E. curvula, potential areas were found in 15 states (Table 5). The states with the largest areas suitable for P. devoniana in agricultural areas were Hidalgo, Michoacán, and Oaxaca (39.5 % of the total), while in forest areas were Michoacán, Jalisco, and Guanajuato (70.5 %). Only five states were suitable for P. lawsonii, of which Puebla, Estado de México, and Michoacán concentrated 95.7 % of the potential area in agricultural areas, while in forest areas were Michoacán, Guanajuato, and Puebla with 97.7 % of the area. Estado de México, Hidalgo and Michoacán accounted for 58.3 % of the potential area with P. montezumae in agricultural areas, while in forest areas were Michoacán, Jalisco and Guanajuato, with 60.6 % of the total.
Table 5 Potential area for silvopastoral systems with Eragrostis curvula and three pine species in Mexico.
| State | Pinus devoniana | Pinus lawsonii | Pinus montezumae | |||
|---|---|---|---|---|---|---|
| Agricultural area (ha) | Forest (ha) | Agricultural area (ha) | Forest (ha) | Agricultural area (ha) | Forest (ha) | |
| Mexico City | 29 280 | 1 770 | - | - | 24 440 | 2 600 |
| Durango | 3 443 | 4 360 | - | - | 410 | 1 558 |
| Guanajuato | 128 800 | 51 100 | 1 571 | 1 732 | 36 240 | 41 960 |
| Hidalgo | 657 100 | 36 600 | - | - | 230 800 | 39 200 |
| Jalisco | 113 300 | 186 100 | - | - | 19 900 | 59 270 |
| Estado de México | 385 600 | 9 800 | 11 200 | 280 | 301 300 | 34 800 |
| Michoacán | 511 400 | 239 900 | 59 650 | 9 480 | 184 500 | 161 200 |
| Nayarit | 9 181 | 40 939 | - | - | 273 | 4 982 |
| Nuevo León | - | - | - | - | 6 702 | 4 318 |
| Oaxaca | 99 430 | 4 870 | - | - | 26 140 | 2 330 |
| Puebla | 435 600 | 27 800 | 40 530 | 710 | 150 900 | 31 300 |
| Querétaro | 166 300 | 35 600 | - | - | 62 580 | 30 900 |
| Tlaxcala | 188 400 | 7 300 | 3 191 | - | 160 200 | 8 600 |
| Veracruz | 9 592 | 99 | - | - | 19 280 | 100 |
| Zacatecas | 20 320 | 27 550 | - | - | 1 746 | 7 676 |
| Total | 2 757 746 | 673 789 | 116 142 | 12 202 | 1 225 411 | 430 795 |
Discussion
Ecological niche models had high predictive power and were better than random modeling in both AUC and partial ROC values, indicating that these models fit the actual distribution of the species (Slater & Michel, 2012). AUC values were higher (0.93) compared to other studies on potential distribution of C. gayana (0.62) (Lastiri-Hernández, Cruz-Cárdenas, Álvarez-Bernal, Vázquez-Sánchez, & Bermúdez-Torres, 2020), as well as P. devoniana, P. lawsonii, and P. montezumae (Cruz-Cárdenas et al., 2016).
The distribution of the five species was correlated with altitude and annual temperature range (Bio07), interrelated variables because temperature conditions change rapidly concerning altitude; the annual temperature range is more marked in mountainous areas (Winkler, Lin, Delgadillo, Chapin, & Huxman, 2019). The tropical species C. gayana and E. curvula have adapted to the temperate conditions of the Neovolcanic Axis, Sierra Madre del Sur, and the Mexican Plateau, due to high capacity for acclimatization in ruderal and degraded environments, which has allowed their expansion in these regions of Mexico (Imaz et al., 2005), while E. curvula has shown the ability to adapt to higher slopes and lower temperatures (Schussman et al., 2006).
In the case of pines, the ecological niche was adjusted to temperate mountain areas, where P. devoniana had the largest potential area, followed by P. montezumae, while P. lawsonii recorded a more restricted distribution, which coincides with the previous modeling for these species (Aceves-Rangel, Méndez-González, García-Aranda, & Nájera-Luna, 2018). The conifers P. devoniana and P. lawsonii shared elevations, while P. montezumae was found on higher slopes, where it can even form co-dominant stands with species more akin to cold conditions, such as Abies religiosa (Kunth) Schltdl. et Cham (Pérez-Miranda et al., 2014), which confirms that P. montezumae has a distinct altitudinal ecological niche.
In the present study, the great potential was noted for the establishment of silvopastoral systems using C. gayana and E. curvula with the conifer species evaluated, mainly in the temperate region of the center and south of the country, which coincides with the high potential found for these two forage species on a regional scale (Sáenz-Reyes et al., 2019). Michoacán, Jalisco, Puebla, Oaxaca, Estado de México, Guanajuato, and Hidalgo had the largest suitable area, mainly in agricultural areas, where it is feasible to intercrop forage crops with conifers widely propagated in forest nurseries (Rueda-Sánchez et al., 2010), which increases the potential of silvopastoral systems with these species; in addition, different altitudinal zones could be covered, because P. devoniana and P. lawsonii occupy low slopes with warmer temperatures, while P. montezumae inhabits colder zones.
It is important to highlight that the use of these species in silvopastoral systems has been evaluated in localities of Michoacán, with chemical and organic fertilization, having forage production of 11.9 Mg∙ha-1, sufficient to feed up to 6 animal units∙ha-1∙year-1, and with an adequate development in height for conifers (Sáenz-Reyes et al., 2016). Therefore, it is feasible to implement this technology in regions whose soils have been severely degraded and have a large livestock production, such as the Morelia-Queréndaro valley, the Bajío region, and the Cuitzeo lake basin (Mendoza, López-Granados, Geneletti, Pérez-Salicrup, & Salinas, 2011).
The temperate region of central and southern Mexico had the greatest potential for silvopastoral systems with the species studied, because, in the case of conifers, they represent the regions of their natural distribution (Farjon et al., 1997). Forages have a great capacity for acclimatization to Mexican soils (Beltrán-López et al., 2018), but could also be suitable for use with other forest species, for which the modeling of the ecological niche of the species represents a useful tool to identify potential areas.
Conclusions
Mexico has a high potential for silvopastoral systems with forage crops C. gayana and E. curvula and conifers P. lawsonii, P. devoniana, and P. montezumae, mainly in the central and southern regions of the country. Most of the potential areas were located in agricultural areas, where the implementation of these systems could contribute to the recovery of forest cover and prevent soil erosion, while in forest areas they could generate a continuous source of food for livestock and prevent animal damage to the ecosystem. Ecological niche modeling is recommended to know the potential areas for establishing silvopastoral systems with forage and forest species.

