











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introduction
The composition of forest ecosystems and genetic resources are affected by processes that threaten their conservation, such as overexploitation, land use conversion, pollution, forest fires, and especially climate change (Soni & Ansari, 2017). This leads to alterations in the distribution, structure, and abundance of tree species in forests, resulting in the reduction of locally adapted population sizes or their disappearance (Urban, 2015).
Global projections in the context of climate change suggest increases in average annual temperatures, droughts, and changes in precipitation patterns (Hammond et al., 2022; Sáenz-Romero et al., 2010). For Mexico, a 9 % reduction in average precipitation is estimated by the year 2060 (Sáenz-Romero et al., 2010). Some species in forest ecosystems are threatened by the effects of climate change, especially in humid environments, where changes towards aridity are predicted (Castellanos-Acuña et al., 2018; Sáenz-Romero et al., 2010, 2016). The evolution of climate will increase probability, frequency, and intensity of extreme weather phenomena such as heatwaves, cold spells, floods, and droughts (Hansen et al., 2012; Walsh et al., 2013). Therefore, in forests, several species and tree populations may not adapt quickly enough to the new climate conditions due to limitations in their ability to migrate through natural means and in their phenotypic plasticity (Gray & Hamann, 2013; Sáenz-Romero et al., 2016). This will be more drastic for species in geographically confined ecosystems, particularly in mountains, where several species would tend to shift their distribution towards other latitudes and altitudes (Gutiérrez & Trejo, 2014). This is especially true for tree populations located at the xeric limits of species found at the lower altitudinal or southern boundary (in the northern hemisphere) of their natural distribution (Mátyás, 2010). Therefore, climate change poses significant challenges for forest managers (Sáenz-Romero et al., 2016).
Reforestation and restoration programs are the most common actions for the recovery of vegetation cover (Castro et al., 2021). In Mexico, data from reforestation programs during the period 2004-2016 indicate that the annual survival rate per state for different ecosystems was recorded between 30 and 53 %, with an overall average of 43 % (Prieto et al., 2018). This poor survival and growth of the plants used has often been attributed to the use of germplasm from unsuitable provenances (Crow et al., 2018; O'Neill et al., 2014; Tomita et al., 2017). In the last two decades, Mexico has implemented reforestation and restoration programs such as the Programa Nacional de Reforestación, Programa de Desarrollo de Plantaciones Forestales and the current program actual ‘Sembrando Vida’, which have increased the demand for germplasm. That program is one of the largest reforestation efforts of the federal government (2018-2024) with a large investment aimed at reducing social inequality and restoring forest and agroforestry cover in degraded sites, which could provide economic income and environmental services (water sequestration, carbon and biodiversity preservation) in the future. The program 'Sembrando Vida', operated by the Welfare Secretariat of Mexico and not by the National Forestry Commission of Mexico, has not considered important technical aspects that could improve the survival and growth of reforestation and restoration efforts, and seems to ignore the expected impacts of climate change (Gallardo-Salazar et al., 2023). The latter are of greater importance in the planning of reforestation, as they are and will become progressively more severe (Astudillo-Sánchez et al., 2017; Ipinza & Gutiérrez, 2014; Sáenz-Romero et al., 2020).
The insufficient compliance with regulations for germplasm management, which guarantees seed supply and improves survival rates, are the main challenges faced by institutions and organizations dedicated to reforestation and restoration (CONAFOR, 2018; Pike et al., 2020; Prieto-Ruiz & Goche-Télles, 2016). All this, despite the fact that since 2014, criteria have been established and published in the Mexican standard NMX-AA-169-SCFI-2016 “Establishment of Forest Germplasm Production Units and Management”, which regulates the processes of collection, transportation, storage, processing, evaluation, and certification to improve the quality of plants used in reforestation (Secretaría de Economía, 2016).
Germplasm movement zones play a key role in mitigating plant adaptation issues at reforestation sites (Hamann et al., 2011; Pike et al., 2020). Germplasm movement involves seed collection (in some cases, cuttings for asexual propagation), seedling production in nurseries, and reforestation in sites where a suitable climate habitat for the seed source is anticipated in the future ( Castellanos-Acuña et al., 2014, 2015; Sáenz-Romero et al., 2016). According to St. Clair et al. (2013) and Havens et al. (2015), seed collection zones are designated by overlaying a combination of climatic variables and delineating common areas as a seed collection zone where the species of interest is distributed, to move them to areas with a suitable climate for their development. Seed transfer zones are geographical areas where the germplasm of a native population can be transferred based on environmental conditions (present or future) similar to those of the seed origin, to minimize the risks of inadequate adaptation.
The study aimed to delineate seed transfer zones for reforestation activities within the Monarch Butterfly Biosphere Reserve (MBBR) in Michoacán and Estado de México, as well as the "Sembrando Vida" program in the Meseta Purépecha in Michoacán, considering the impacts of climate change. The goal is to develop a practical tool for determining the appropriate seed sources and their target areas, considering that the suitable climatic conditions for a forest population may shift to a different location in the future. Although the primary focus is on enhancing the "Sembrando Vida" program, the zoning recommendations could be applicable to various reforestation and restoration initiatives.
Materials and Methods
Study areas
The study areas consist of the Meseta Purépecha in Michoacán, covering an approximate area of 660 000 hectares, and the Monarch Butterfly Biosphere Reserve (MBBR) on the border between Michoacán and Estado de México, with an approximate area of 56 256 hectares. Both areas are located within the Trans-Mexican Volcanic Belt (Figure 1).
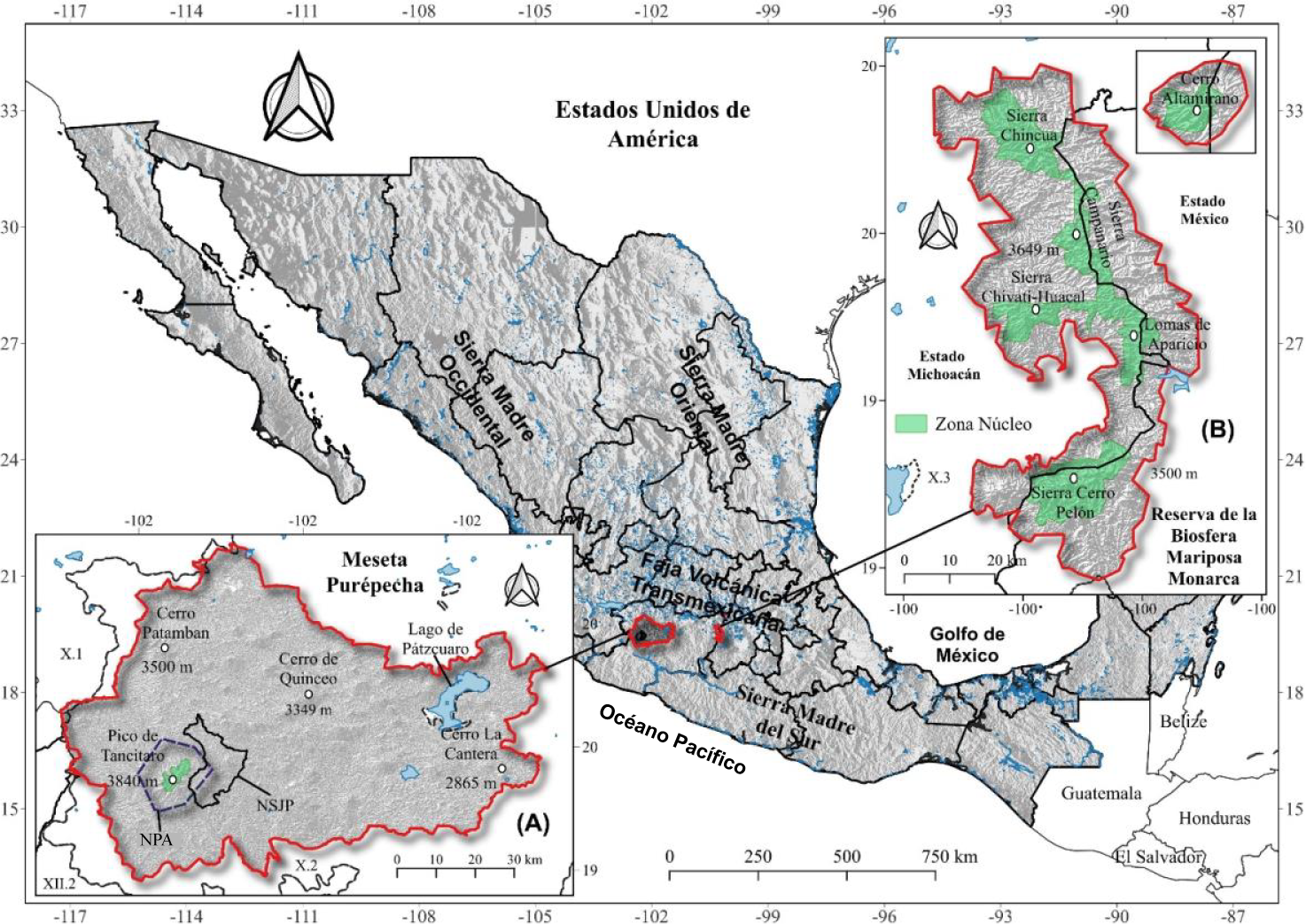
Figure 1 Political divisions of Mexico, prominent geographic regions and study areas: Meseta Purépecha in Michoacán and Monarch Butterfly Biosphere Reserve (MBBR) on the border of Michoacán and Estado de México. In the Meseta Purépecha, the indigenous community of Nuevo San Juan Parangaricutiro (NSJP) and the Pico de Tancítaro Natural Protected Area (NPA) are indicated.
We prioritized the analysis in the indigenous region of the Meseta Purépecha, comprised of the indigenous community of Nuevo San Juan Parangaricutiro (NSJP), Paracho, Cherán, Chilchota, Nahuatzen, Tancítaro and Taretan, which have economic support for beneficiaries of the federal program ‘Sembrando Vida’ (Figure 2). Elevations in this region range from 1 600 to 3 860 m. The dominant climate is temperate sub-humid with rainfall between 800 and 1 100 mm per year, although the region's rugged topography produces a wide variety of microclimates. Vegetation consists mainly of pine-oak forests, conifers and agriculture (Hall et al., 2015).
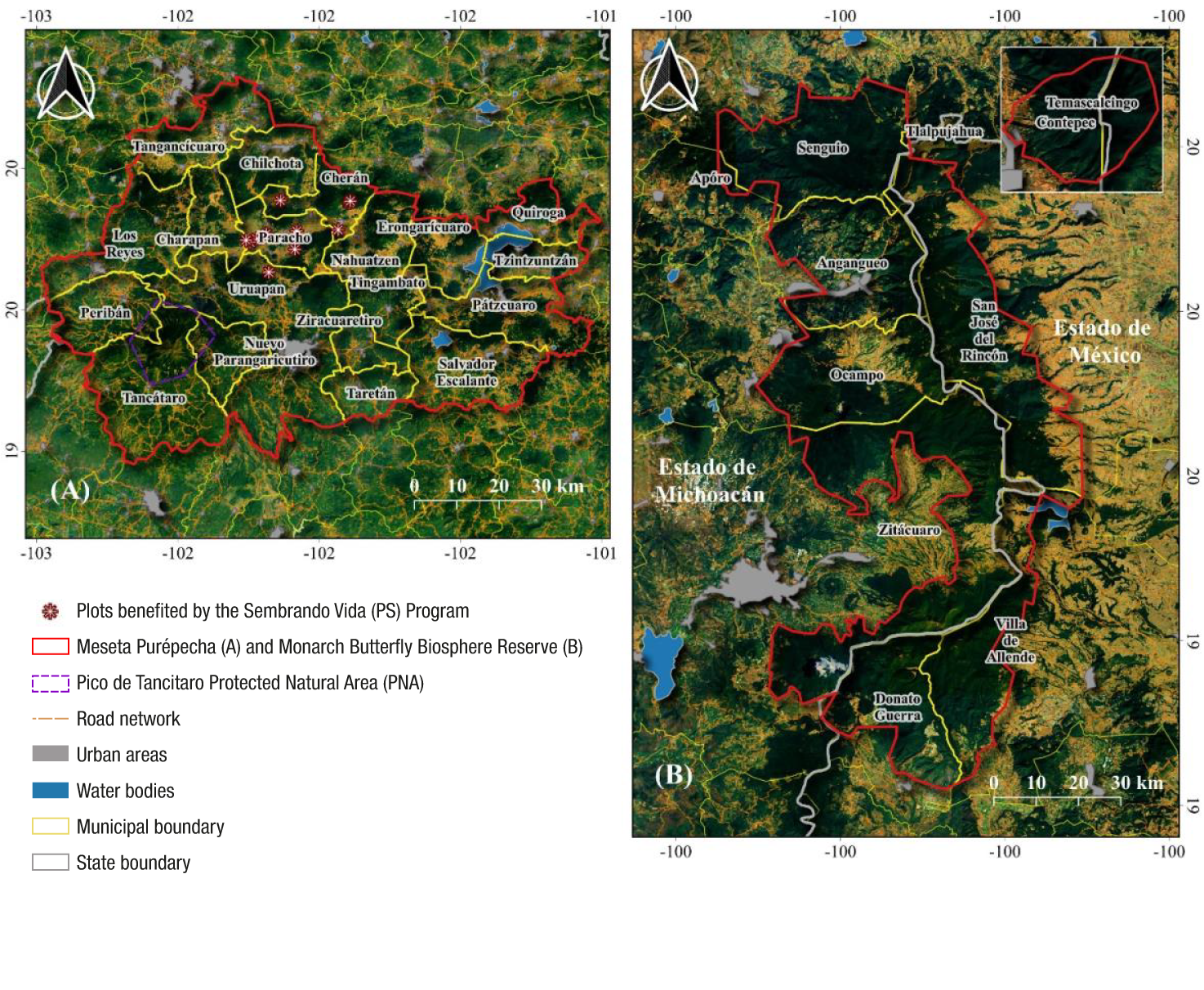
Figure 2 Description of study areas: A) Meseta Purépecha, Michoacán, and plots benefited by the 'Sembrando Vida' program and B) Monarch Butterfly Biosphere Reserve on the border of Michoacán and Estado de México.
The MBBR serves as a winter refuge for the monarch butterfly Danaus plexippus (L.) with elevations between 1 800 to 3 640 m with steep slopes (Figure 2). The climate is temperate sub-humid with rainfall in summer with mean annual precipitation of 830 mm and mean annual temperature of 15 °C. The lower elevations are characterized by temperate pine-oak forest and mainly rainfed agricultural plots. At higher elevations, there is oyamel forest (Abies religiosa [Kunth] Schltdl. & Cham.), habitat of the monarch butterfly, with patches of grassland and shrublands (de Azcárate Cornide et al., 2003).
Climate data and habitat modeling
Data were obtained from Castellanos-Acuña et al. (2018), whose vector representation model files (shapefiles) are in the digital repository available at https://doi.org/10.5281/zenodo.1052141. These provide a seed zone system for Mexico based on a reference climate (1961-1990), a recent climate (1991-2010), and the projected climate for the average of the period 2041-2070, which we will refer to for simplicity as 2050 (Wang et al., 2016; https://sites.ualberta.ca/~ahamann/data/climatena.html). Climate zones were generated from mean coldest month temperature (MCMT) and an aridity index [AI = (mean annual temperature + 10) / (precipitation/1 000)], to represent the relationship between temperature and plant-available moisture (Bower et al., 2014). Higher values indicate greater aridity.
The corresponding intervals for the MCMT are set every 3 °C. The aridity index intervals are variable in absolute values but have approximately the same width under a logarithmic transformation. The superposition of these two climate variables for the reference period (1961-1990) resulted in 63 climate zones (details in Castellanos-Acuña et al., 2018). Subsequently, that same zoning criterion was used for a projected forward-looking scenario of the Representative Radiative Forcing Greenhouse Gas Concentration Trajectory (RCP) 4.5 W∙m-2. This scenario is considered relatively optimistic; a pessimistic scenario (RCP 8.5 W∙m-2) was not used, because it would imply excessive germplasm movement at present, which would generate a risk of frost damage to reforestation seedlings. Although the climate data grids under the reference and projected scenario of Castellanos-Acuña et al. (2018) have a spatial resolution of 1 km2, for this study it was reduced to 400 x 400 m in order to improve it.
As a complement to decide seed sources and destination areas, we used climate habitat modeling data for P. pseudostrobus Lindl. and A. religiosa from the reference period (1961-1990) and the future (decade centered on 2060) generated by Gómez-Pineda et al. (2020). The species under study have ecological, economic and conservation importance, which are distributed from the high and low parts of the physiographic provinces of Mexico. This is the case of the economic importance of P. pseudostrobus (2 200-3 000 m; Farjon, 1997) for sawtimber production; also, the dense forest of A. religiosa (2 400-3 600 m) is of great importance for the conservation of the Monarch butterfly that spends the winter in the MBBR.
The objective of the zones was to find a coupling with the climate of the decade centered on the year 2050 or 2060. If a closer scenario were chosen (2030, for example) there is a risk that the transfer of seed sources will be an insufficient climate transfer and the planted trees will be stressed when they reach reproductive age or commercial size. A more distant scenario (2090, for example) involves the risk that seedlings will be planted on a site that is too cold at present and suffer severe frost damage.
Climatic habitat
The percentage change in climatic habitat suitable for P. pseudostrobus and A. religiosa was estimated from Gómez-Pineda et al. (2020) modeling of the projected seed collection area and reforestation area (deployment) for each tree species, based on the current and projected forward scenario for the average of the period 2056-2065, which we will refer to for simplicity as the decade centered on 2060 (Sáenz-Romero et al 2010; https://charcoal2.cnre.vt.edu/climate/). Habitat change was determined with the formula: % change = [(S1-S0) / S0] * 100 %; where, S0 is the total area occupied by suitable climatic habitat in the study area according to the reference period (1961-1990) and S1 is the total area projected to occupy in the decade centered on 2060. This to visualize the reduction of the climatic habitat of the species under study in the Meseta Purépecha and MBBR.
Seed Transfer Zones
The area where forest vegetation is actually present (because climatic habitat usually overestimates the area actually occupied) was obtained from the temperate forest occurrence information layer published in Latorre-Cárdenas et al. (2023). The presence of P. pseudostrobus and A. religiosa was identified from the National Forest and Soil Inventory (INFyS) database produced by CONAFOR 2015-2020 (https://snmf.cnf.gob.mx/datos-del-inventario/). Based on the aforementioned data and the altitudinal range of the distribution of each species in the field, these layers were overlaid on the climatic habitat of each species, estimated by Gómez-Pineda et al. (2020), in the QGIS program version 3.32.1 (QGIS, 2023), for the location of sites with potentially seed-producing trees. The layer of Germplasm Movement Zones, indicated in the NMX-AA-169-SCFI-2016 standard (Secretaría de Economía, 2016), which are defined as areas with relatively similar ecological and climatic characteristics, was overlaid.
Finally, climatype maps were created, a subdivision of the climatic habitat suitable for each species resulting from superimposing on it the climate zones based on the MCMT and aridity index intervals. This to account for (at least partially) the fact that populations are adapted to a portion of the species' suitable climatic habitat and not to the whole habitat (see Sáenz-Romero et al., 2012b and Ortiz-Bibian et al., 2017 for P. pseudostrobus and A. religiosa, respectively). The climatype concept used is similar to that developed by Tchebakova et al. (2005) where the subdivision is determined by intervals of climatic values; however, in the present study, the size of the intervals was not based on the minimum significant difference between provenances, obtained in garden trials by those authors, but on zoning of Castellanos-Acuña et al. (2018).
Results and Discussion
Climate zones for seed collection and climate zones for reforestation
The boundaries of seed collection zones (1961-1990 reference climate) and deployment zones (projected climate for the 2050s, RCP 4.5 scenario) in the Meseta Purépecha and the MBBR resulted in 19 climate zones out of 63 reported for Mexico (Castellanos-Acuña et al. 2018; Figure 3). The MBBR reflects altitudinal gradients with warm areas at lower altitudes (in the direction of the Balsas Depression) and cooler areas in the mountains.
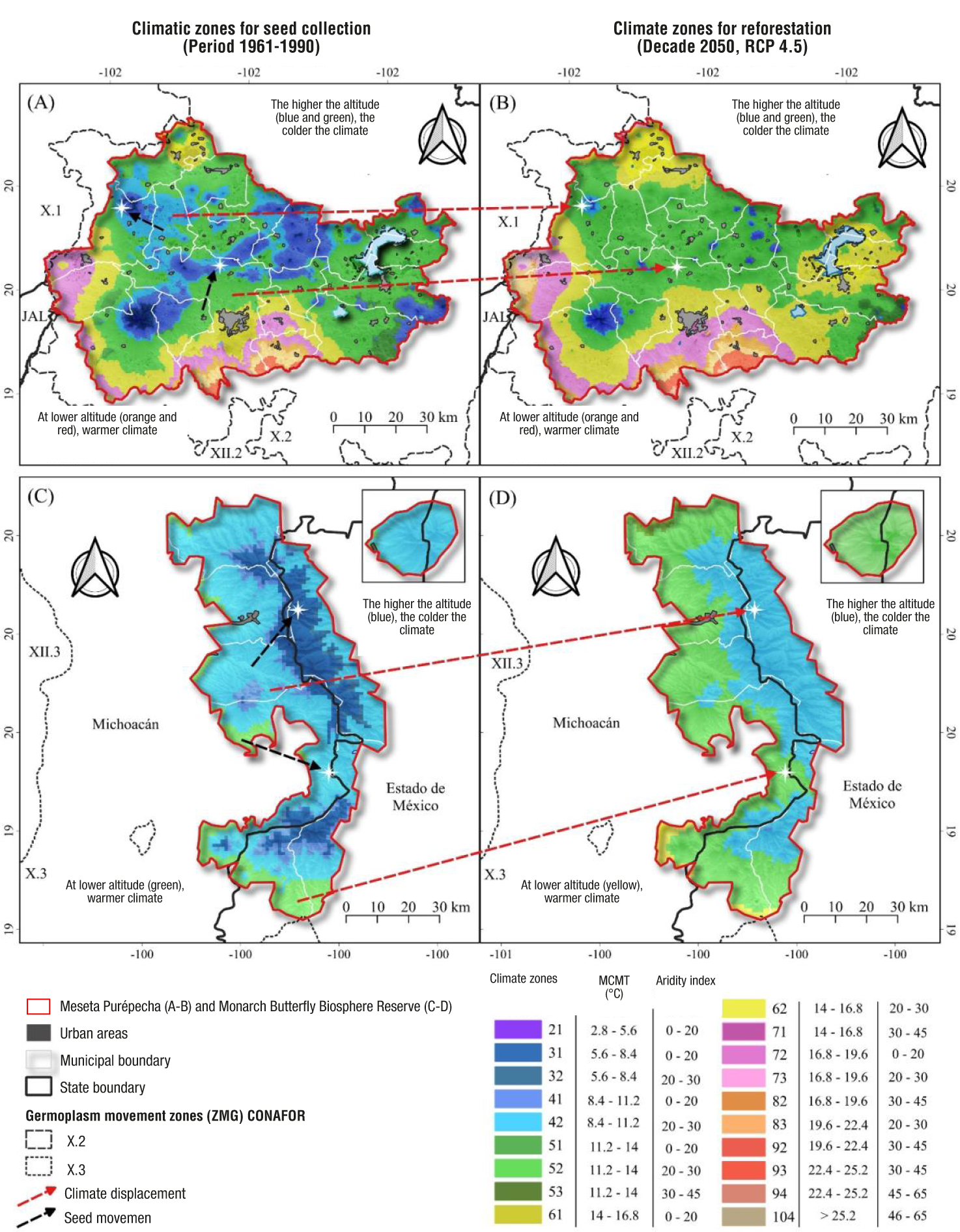
Figure 3 Climate zones for seed collection (defined under a 1961-1990 reference climate) and target zones for reforestation (deployment) under a future climate (2050, RCP 4.5), resulting from the intersection of the mean temperature of the coldest month (MCMT) and the aridity index, for the Meseta Purépecha and the Monarch Butterfly Biosphere Reserve (MBBR). The arrows indicate where the climate currently occurring at a given site will occur (red arrows) and what the movement of germplasm might look like in the present to compensate for projected climate change (black arrows).
Under the current reference climate, in the region of the Meseta Purépecha, the coldest zones are found in the higher parts of the municipalities of Los Reyes and Tancítaro with elevations of 2 794 to 3 840 m; in contrast, the warmest regions are located in the south of Uruapan and Taretan, with altitudinal ranges of 630 to 1 800 m (Figure 3A). In the case of the MBBR, the coldest areas are located in San José del Rincón and Villa de Allende, and the warmest areas are in the southern part of the municipality of Donato de Guerra in Estado de México (Figure 3C). However, both regions will be warmer and drier in the future according to projections for the year 2050 (Figure 3B and 3D). These changes can be seen by comparing the zoning under the reference climates (1961-1990) and the projected future climate (2050). For example, in the Meseta Purépecha, in the future, the green and blue areas (at their margins corresponding to lower altitude) that generally correspond to forests dominated by P. pseudostrobus are significantly reduced (Molina-Sánchez et al., 2019). Similarly, within the MBBR, dark blue areas are reduced, which generally correspond to oyamel fir forests (de Azcárate Cornide et al., 2003; Sáenz-Romero et al., 2012a).
The reduction of climate zones suitable for conifers in the Meseta Purépecha and in the MBBR will generate decoupling between the areas currently occupied by forest populations and the site where the climate for which they have adapted through a long evolutionary process have occurred (Sáenz-Romero et al., 2016). In other words, the climate in which forest populations have evolved will occur in the future in a different location (at a higher altitude in our case) or may even disappear altogether (Rehfeldt et al., 2012). This displacement of the suitable climate will occur at a much faster rate than the eventual displacement of forest populations by natural means (seed dispersal and colonization of new sites). Thus, trees that will remain on a site with adverse climate will progressively suffer more and more stress from warmer droughts, predisposing them to pest and disease attack and, eventually, death (Gómez-Pineda et al., 2020; Hammond et al., 2022; Rehfeldt et al., 2014; Sáenz-Romero et al., 2016, 2023; Seda & Ömer, 2019).
The effects of the decoupling between forest populations and favorable climate are already observable. In the Meseta Purépecha, branch defoliation has been detected during the dry season in the xeric limit of P. pseudostrobus. Tree branches that often no longer recover during the following rainy season accumulate dead branches to a threshold where they become severely weakened, and these are candidates for attack by bark beetles, causing total tree death (López-Toledo et al., 2017). Similarly, defoliation was observed in A. religiosa in the MBBR (Sáenz-Romero, 2015) and on Cerro Tláloc (Flores-Nieves et al., 2011), apparently related to climate change.
This situation calls for proactive management for the re-coupling of forest populations to a favorable climate, even if it means they will occur in a different location. As a general strategy, regardless of the forest species, it is possible to use the climate zoning in Figure 3 as a guide for seed transfer aimed at approximating such a coupling. The way to proceed would be as follows:
Select the site to be reforested on the current climate zoning map (1961-1990 reference climate) (Figures 3A or 3C).
Locate the same site to reforest, but on the 2050 climate zoning map (Figures 3B or 3D) in order to determine the possible future climate.
Return to the current climate map (Figures 3A or 3C) and locate the zones where the climate that will occur in the future is currently found at the site to be reforested. These zones with current climate are the zones of present-day seed collection.
Figure 3 shows the above procedure with reforestation sites indicated with a white star. For example, the upper star of the planting site, both within the Meseta Purépecha (Figure 3A) and the MBBR (Figure 3C), is in a dark blue climatic zone (MCMT = 5 to 8 °C; IA = 2 to 20) and becomes warmer in the future (light blue, MCMT = 8 to 11 °C) and drier (IA = 20 to 30). For the lower star example (Figures 3B and 3D), the area to be reforested is light blue and becomes warmer (dark green, MCMT = 11 to 14 °C, AI = 20 to 30). For this second case, it is suggested to collect seeds under the current climate (green color), produce seedlings in the nursery and, subsequently, reforest sites where that same green climate will occur, but projected for the year 2050 (according to the maps based on the climate zones of Castellanos-Acuña et al., 2018). In practice, it would be advisable that nursery production be done in the climatic region of the site to be reforested to facilitate plant acclimatization, even if the seed is imported from warmer sites. In other words, the transfer of germplasm from the site chosen as seed source to the planting (deployment) site is preferable, as far as possible, to transport the seed for nursery supply rather than transporting the plant once it's already grown.
Location of collection and reforestation zones using the suitable climatic habitat for reforestation
Although the use of the previously described climatic zoning is considered an advancement, it's important to note that in the climatic zones currently compatible with the future climate of the site to be reforested, the species of interest desired for reforestation may or may not be available. This is because the climatic zones compatible with the future climate are usually located at lower altitudes, where a different forest species could be distributed or even the original arboreal vegetation could no longer exist due to land use change.
While the climatic habitat projection can guide the location of potential collection sites for a species of current interest, the fact that the suitable climatic habitat is in a location does not guarantee the presence of the species. This can be due to a multitude of factors that prevent it: competition effects from other species, lack of propagules that have previously colonized the site, illegal logging, and others such as the change in land use from temperate forest to avocado orchards in Michoacán (Latorre-Cárdenas et al., 2023).
One way to have a more realistic approach to identifying potential seed collection sites is to overlay the climatic habitat of the species of interest with a vegetation map, in this case, of pine-oak, pine, and conifer forests (obtained in Latorre-Cárdenas et al., 2023). The result is shown in Figure 4, where it is possible to notice that the area occupied by the suitable climatic habitat, both for P. pseudostrobus and A. religiosa, is much larger (particularly for the former species) than the area with forest effectively available for seed collection.
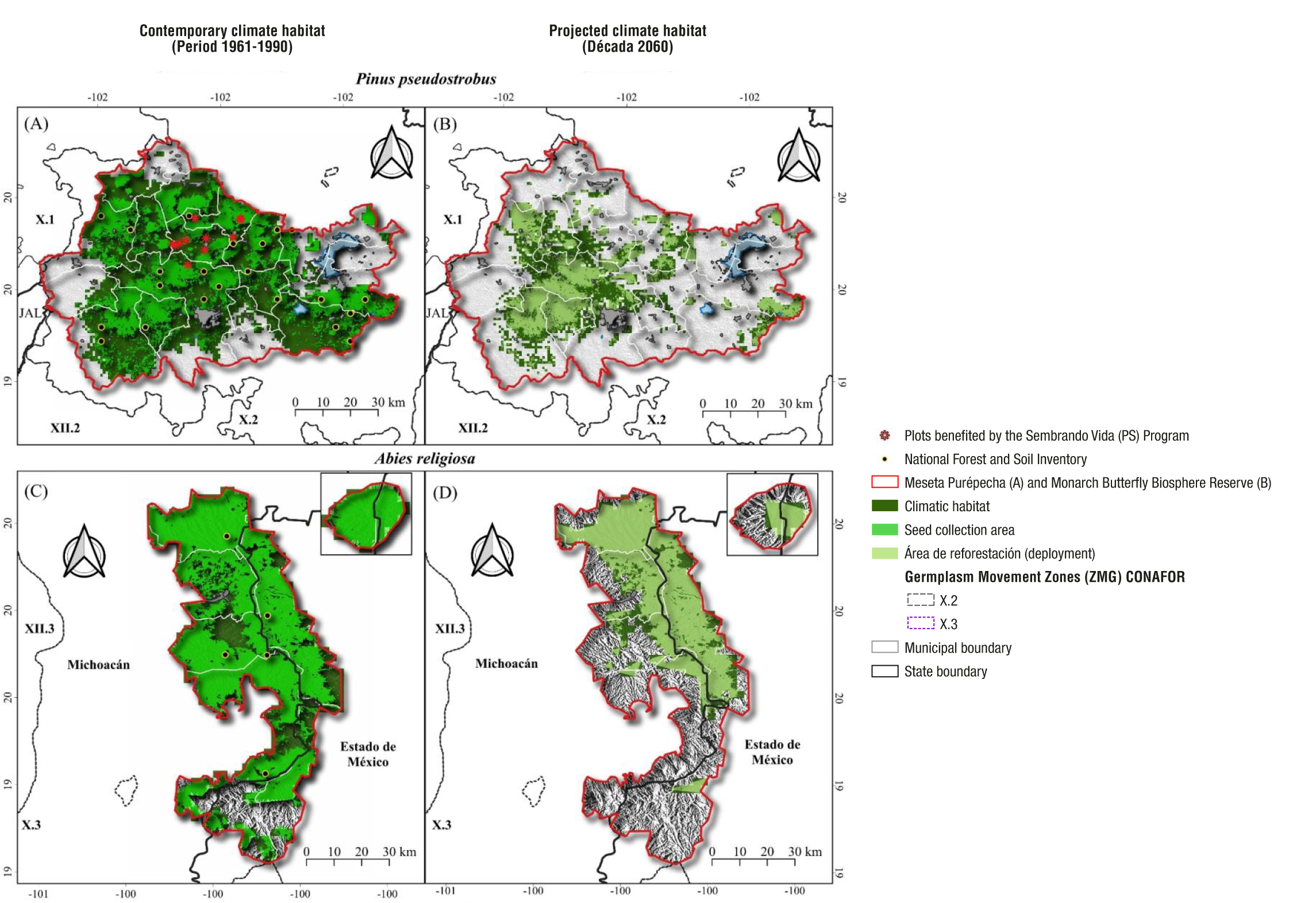
Figure 4 Suitable climatic habitat for P. pseudostrobus and A. religiosa, seed collection areas of the current period (1961-1990) and reforestation areas (deployment) to match the future climate (2060s) in the Meseta Purépecha and Monarch Butterfly Biosphere Reserve in Michoacán and Estado de México, respectively. The seed collection area and reforestation layer overlapped with the current and future climate habitat layer, respectively. Suitable climatic habitat based on Gómez-Pineda et al. (2020) and area effectively covered by forest according to Latorre-Cárdenas et al. (2023).
It is also important to note that the area for reforestation with the species of interest is significantly reduced when considering the expected effects of climate change, compared to the current area potentially useful for seed collection at present (Figures 4A and 4C), due to the progressive reduction of climatic habitat suitable for conifers, as a result of climate change (Figures 4B and 4D). Site options for planting a conifer species of interest are reduced because of a projected 48 % decrease in favorable climatic habitat for P. pseudostrobus in the Meseta Purépecha and 49 % in the MBBR for A. religiosa (Figure 5). This is not exclusive to the two areas studied, but a general trend for temperate forests in Mexico (Rehfeldt et al., 2012).
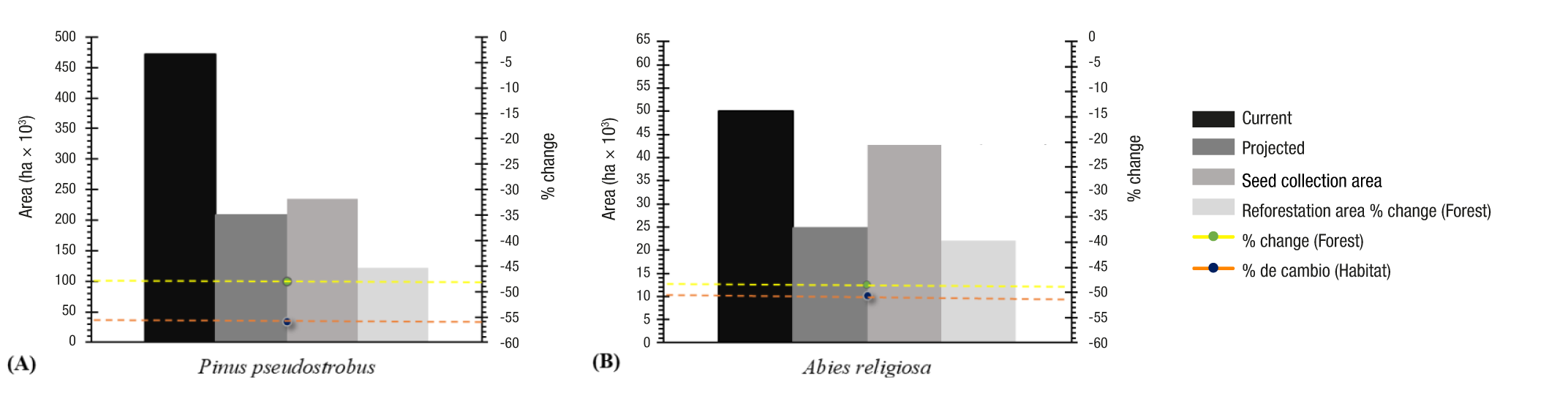
Figure 5 Climatic habitat, seed collection areas and reforestation areas (deployment) of the analyzed species according to the current scenario (1961-1990) and projected scenario (2060s) in the Meseta Purépecha (A) and Monarch Butterfly Biosphere Reserve (B). The total area (ha) is presented with percentages (%) of change with respect to the present. There is a significant reduction in the area of climatic habitat for each taxon in the study area.
The double reduction of options (scarce trees effectively present within the current climatically favorable habitat and the reduction of this in the future) affects, for example, the beneficiaries of the federal program ‘Sembrando Vida’ (plots shown in Figures 4A and 4B). For them, the priority species for reforestation is P. pseudostrobus in the Meseta Purépecha. In the case of the MBBR, the options of sites for obtaining seed at low altitude and sites for reforestation with A. religiosa are reduced, due to the change of land use in the lower limit of the A. religiosa forest and the reduction of its favorable habitat in the future, particularly in the southern part (Figures 4C and 4D).
For P. pseudostrobus and A. religiosa, the reduction of suitable habitat occurs at the lower altitudinal limit, which has been called the 'xeric limit' because it is warmer and, therefore, usually drier than the rest of the climatic habitat (Mátyás, 2010). Therefore, in the lower altitudinal band of climatic habitat that will disappear, it will be necessary to consider the replacement of species by one adapted to warmer sites: P. devoniana could replace P. pseudostrobus at low altitudes in the Meseta Purépecha (Sáenz-Romero et al., 2012b) and P. pseudostrobus would replace A. religiosa in the lower parts of the MBBR buffer zone (Ortiz-Bibian et al., 2017).
Subdivision of climatic habitat into climatypes
Subdivision of the climatic habitat of P. pseudostrobus and A. religiosa by overlaying the climatic zoning of Castellanos-Acuña et al. (2018) with the present and future climatic habitat developed by Gómez-Pineda et al. (2020), generated 10 ‘climatypes’ (climatically defined, not based on provenance trials in the sense of Rehfeldt & Jaquish [2010] or Tchebakova et al. [2005]), under current climate in the Meseta Purépecha (Figure 6A) and five in the MBBR (Figure 6C). The projection of these same climatypes for the year 2050 shows that the colder ones will severely decrease or disappear, while the warmer ones will expand their area (Figures 6B and 6D). For example, the sites currently occupied by the climatypes indicated as ‘dark blue’ and ‘light blue’ will be replaced by warmer ones represented by ‘light blue’ and ‘green’, respectively (Figure 6).
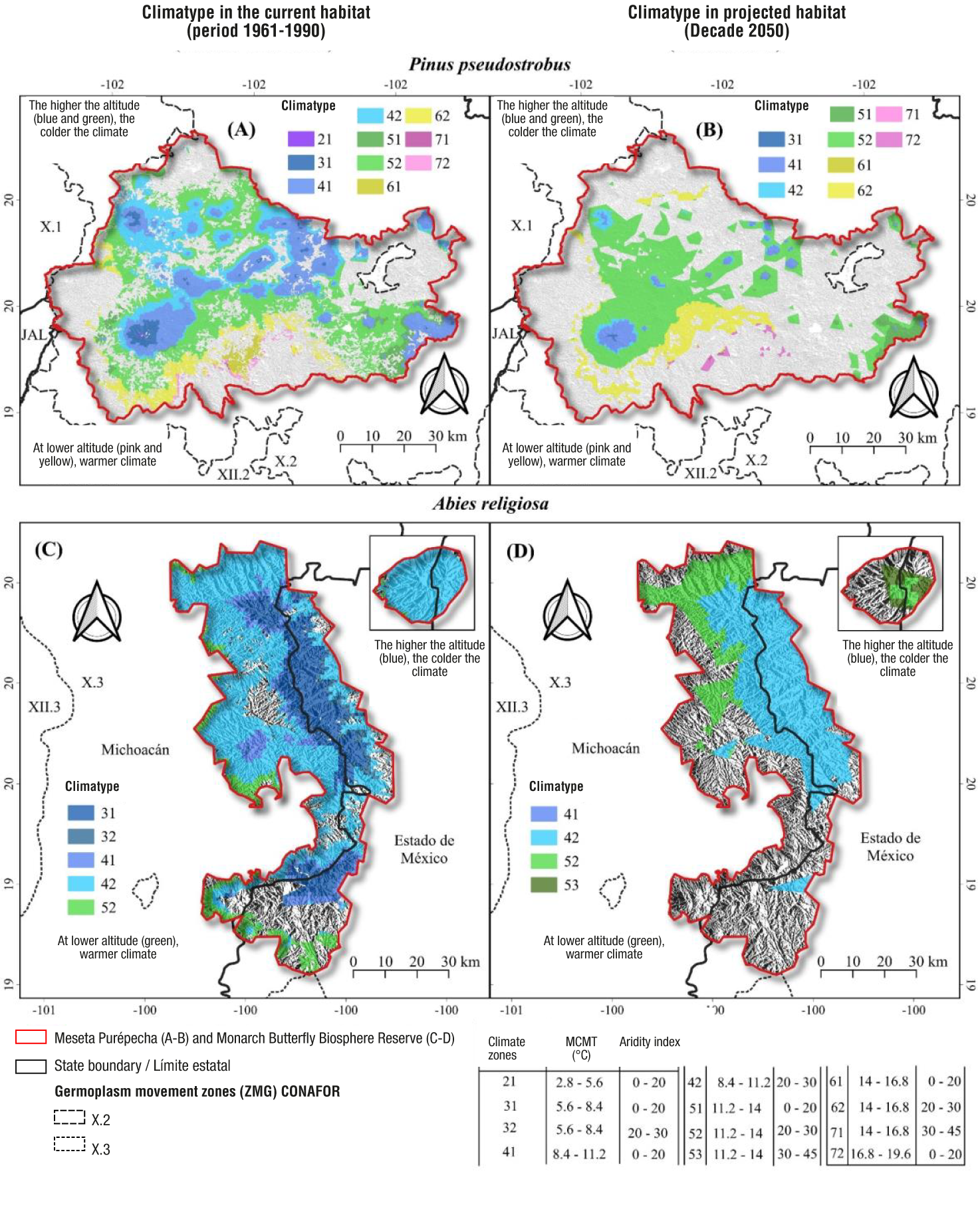
Figure 6 Climatypes (subdivision of the favorable climatic habitat based on mean coldest month temperature [MCMT] intervals and aridity index) of Pinus pseudostrobus in the Meseta Purépecha and Monarch Butterfly Biosphere Reserve in the state of Michoacán and Estado de México in the current (1961-1990) and projected climate for the decade to 2050.
The guidelines for the use of the climatypes shown in Figure 6 are essentially the same as those described for the climate zones in Figure 3. Seed collected within a climatype should be used to produce seedlings to be used in reforestation sites that will be within the same climatype in the future. For example, seed collected from sites within the ‘light blue’ climatype (Figure 6A or 6C) should be used in reforestation of sites occupied by the future ‘light blue’ climatype (Figure 6B or 6D). This subdivision of suitable climatic habitat allows accounting, at least in part, for genetic differentiation among populations within the species, which has been demonstrated with provenance trials from climatically contrasting sites, planted at sites with climates different from the seed origin (Leites et al., 2012a, 2012b).
Implications of Management Adaptive to Climate Change
The use of current climatic zones for seed collection and climatic zones for reforestation purposes, which partially overlap but are not the same, implies the change of a central paradigm. Before climate change, it was assumed that local seed sources would provide seedlings with the ability to adapt to a reforestation site close to the collection site (Giencke et al., 2018). The climate change that is taking place, with the consequent mismatch between seed source climate and the site with a favorable climate for the planted trees when they reach adulthood, requires that adaptive management to a changing climate include partial abandonment of the idea that the local seed source is the desirable one (Ipinza & Gutiérrez, 2014; Sáenz-Romero et al., 2016). Recent evidence of catastrophic forest fires linked to climate change (e.g. 18 million ha in the summer of 2023 in Canada), and the exceeded threshold of +1.5 °C mean annual temperature compared to the pre-industrial period, indicate a clear acceleration of climate change, which makes it even more urgent to take management measures other than the usual ones (Sáenz-Romero, 2024).
A central decision is to determine how far it is acceptable to move a seed source from the collection site to the reforestation site. This involves determining which future scenario is the desired outcome for which alignment is sought, which in turn implies choosing a future time horizon and a greenhouse gas concentration scenario (RCP 4.5, 6.0 or 8.5). It also requires knowledge of the adaptive phenotypic plasticity that will allow a plant to survive in a site that is initially cooler than the one to which it is transferred, but which will be close to its optimum in the future. At least for shade tolerant species, such as A. religiosa, an alternative for the protection of newly established seedlings at a higher altitude than their origin is the use of shrubs as nurse plants, which has been successfully experimented in the MBBR core zone (Carbajal-Navarro et al., 2019).
Experiments of assisted migration in North America indicate from accumulated experience that a climatic transfer (difference between destination and origin climate) of the order of 2 °C to 3 °C to colder sites (in this case to sites of higher altitude) is acceptable (Sáenz-Romero et al., 2021). In the present proposal for the use of present and future climate zones, the transfer from one climate zone to the contiguous colder one, implies a movement of 3 °C on average, due to the definition of 3 °C intervals that was used for the zoning of Castellanos-Acuña et al. (2018). The transfer movement is recommended to be done as soon as possible so that when P. pseudostrobus and A. religiosa trees are approximately 25 years old in 2050 (reproductive age; Madrigal-Sánchez, 1967), they have the conditions to reproduce successfully, being in a favorable climatic habitat.
Reforestation with seedlings originating from seed collected in a site with a climate similar to the future climate of the target area should be carried out prioritizing sites that are already disturbed (by deforestation, forest fires or pest outbreaks) and, preferably, not replacing natural stands that are still healthy.
In the species and regions analyzed in this study, the movement of seed sources to destination sites in reforestations is possible, since there are still sites high enough in altitude to achieve at least some degree of adaptation to future climates. There are other much more difficult cases, such as Picea mexicana Martínez, where the best option would be an assisted transcontinental migration to save it from extinction (Mendoza-Maya et al., 2022).
Limitations of the climate zoning proposal
While the use of the proposed current and future climatic zoning will contribute to reducing the risks of seedling maladaptation and subsequently, adult trees to future climates, there are other factors that could influence the success of reforestation efforts. For example, the availability of viable seeds in sufficient quantities for reforesting priority areas, quality of nursery-produced plants, planting date, competition between the planted species and existing ones at the destination (deployment) site, as well as soil conditions, exposure, and slope degree. These factors could decrease the expected survival rate; therefore, it is essential that, in addition to the proposed zoning to decide on seed source and destination site, there is input from forestry experts, biologists, and local stakeholders who can assess whether the planting site has soil and vegetation conditions compatible with the selected species and origin chosen for reforestation.
Conclusions
Climate is causing a mismatch between the site occupied by forest species populations and the location of the climate to which they are adapted. This generates the need for more proactive management of forest genetic resources, aimed at achieving re-alignment once again. It is recommended to move seed sources from warmer and drier locations to currently cooler and wetter (on average 3 °C cooler) planting sites, to compensate for the climate change that has already occurred and is expected to continue over the next decades. To facilitate such a decision, the use of a climatic zoning (3 °C intervals and a variable interval aridity index) is proposed, in which the seed source under current climate (1961-1990) belongs to the same climatic zone as the site to be reforested, regarding a future scenario (RCP 4.5, year 2050).

