











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkMost ecophysiological studies in epiphytes have focused on getting explanations about plant geographical distribution and how they respond to changes in their habitat (Schmidt et al., 2001). Three of the most important ecological factors influencing plant development and distribution are water, irradiance and temperature (Adams et al., 1997; Lambers et al., 2008; Smith et al., 1997; Zotz 2005).
Water is the most limiting factor for growth and survival in arid environments (Nobel, 1988). Similarly to terrestrial plants in arid environments, epiphytes grow in habitats with dry microclimate, mainly due by intermittent water supply (Benzing, 1978). Epiphytes show modifications to offset drought. For example, many bromeliads present an elaborate phyllotaxis and cavities that hold water, phytotelmata, that in combination create efficient water-storage tanks (Benzing, 1980). Crassulacean acid metabolism (CAM) is a physiological adaptation which confers three to six times higher water use efficiency because stomatal opening occurs at night, when evaporative demand is lower (Nobel, 1988; Ehleringer and Monson, 1993; Winter and Smith, 1996). The atmospheric epiphyte Tillandsia ionantha Planch. after 6 months of drought was still green and maintained a constant level of osmotic and water potential and a positive carbon dioxide uptake (Ohrui et al., 2007).
Temperature is the factor which best explains the distribution of a species (Gibson and Nobel, 1986; Sanford, 1969). Temperature regulates most of the basic aspects of cell metabolism; high temperatures denature enzymes and lower temperatures freeze tissues and damage cell membranes (Palta et al., 1993; Pastenes and Horton, 1996). Furthermore, most plants grow best when temperatures fluctuate at least a 6 to 7 ºC between day and night (Lüttge, 2004). This is marked in CAM plants because the CO2-binding enzymes, phosphoenolpyruvate carboxylase (PEPC) is stabilized at low temperatures, and Ribulosa 1-5 biphosphate carboxylase (Rubisco) activity is favored by warm temperatures (Carter et al., 1995; Israel and Nobel, 1995). Cold temperature is the major limiting factor that affects the distribution of tropical epiphytes (Benzing, 2000). However, for the atmospheric Tillandsia usneoides (L.) L. low temperature is not limiting (Garth, 1964). Similarly, for Polypodium vulgare L., an epiphytic fern, long severe expositions to frost don’t lead to visible damage (Zotz, 2005). Studies of tolerance to high temperatures in epiphytes are lacking. One of the few studies shows that the epiphytic lichen Evernia prunastri (L.) Ach. can tolerate air temperatures of 40 ºC for short periods and exhibits low acclimation to increasing temperatures (Pisani et al., 2007).
Some studies have explored the importance of size-related physiological responses in epiphytes. Almost all studies related to bromeliad size have focused on the measurement of the responses of the photosynthetic apparatus, gas exchange, transpiration rates and morphological changes in relation to environmental factors (Adams and Martin, 1986a, b; Schmidt et al., 2001; Schmidt and Zotz, 2001; Zotz, 1997; Zotz and Thomas, 1999). For example, adult tank-forming bromeliads under well-watered conditions show higher rates of CO2 exchange and transpiration than smaller ones (Adams and Martin, 1986a). Many bromeliads with a rosette growth pattern have the ability to form a tank and store water to avoid drought. However, many tank-forming bromeliads do not have water storage capacity during early stages that could suggest a higher mortality for desiccation than adult plants (Benzing, 2000).
This study compares drought and extreme temperature tolerance for two Tillandsia dasyliriifolia Baker populations and the effect of plant size in these parameters. Tillandsia dasyliriifolia is a CAM bromeliad (Ricalde et al., 2010) that grows along costal dune scrubland vegetation in the northern Yucatan Peninsula. We measured the tolerance of T. dasyliriifolia to drought and extreme temperature under controlled conditions. For this experiment, individuals of different sizes and from different populations were tested. Since there is a temperature gradient that decreases to east, while the percentage of winter precipitation increases in the same direction, it is expected that the larger plants from more arid and warmer conditions will be more tolerant to drought and extreme temperature.
Materials and methods
Study site. The climate of the northern coast of the Yucatan Peninsula is semi-arid and presents a precipitation gradient increasing from west to east (Orellana et al., 2009) San Benito has average mean air temperature of 26 ºC, a maximal of 45 ºC and a minimum of 10 ºC and receives less than 500 mm average annual rainfall, with a dry season from March to May (Orellana, 1999). Ria Lagartos presents an average mean air temperature of 25.6 ºC, a maximal of 39.1 ºC and a minimum temperature of 10.8 ºC, with 656.1 mm of average annual rainfall, most of which falls during the rainfall season (June - October) and some precipitation (20 - 60 mm) occurs in the northwind season from November to February (Orellana, 1999).
Drought tolerance. Plant size was defined as maximal leaf length (Schmidt y Zotz, 2002). Maximal leaf length for small plants (S1) was 9.6 ± 0.4 cm (mean ± standard deviation) and tank volume was 13.8 ± 0.5 ml. Maximal leaf length for larger plants (S2) was 35.6 ± 1.2 cm and tank volume was 75 ± 3.1 ml. Ten plants per size and site were acclimated for 4 weeks to 22.5 mol m-2 d-1 PPFD, 33.0 ± 0.5 / 23.1 ± 0.7 ºC diurnal/nocturnal air temperature and well-watered conditions. PPFD and temperature were similar to those in the microhabitats where Tillandsia dasyliriifolia grows (R.J. Castillo unpublished data). The impact of drought was measured using the visual wilting stages method developed by Engelbrecht and Kursar (2003), based on leaf angle, leaf rolling and we included a necrosis percent value.
Leaf measurements. Thickness was measured in the middle part of the leaf of 30 well hydrates plants of Tillandsia dasyliriifolia using a digital caliper. To determinate the parenchyma percentage in completely expanded leaves were transversally sectioned in the middle of the blade using a razor blade. Temporary slides were made for study using light microscopy. Digital photographs were taken with an objective lens of 4×, photographs were analyzed with ImageJ 1.48c software (Wayne Rasband, National Institutes of Health, USA).
Extreme temperature tolerance. For this experiment, San Benito and Ria Lagartos plants were acclimated to day/night ambient temperatures of 30/20 °C (mean temperature of the rainy season in the Yucatan Peninsula, (Cervera et al., 2006), 50 % of total daily PPFD and tanks were refilled every second day. For each site, we selected seven individuals for each plant size, S1 and S2. Tolerance was determined using the vacuolar uptake of the vital stain neutral red (3-amino-7-dimethylamino-2- methylphenazine hydrochloride, Sigma, Didden-Zopfy and Nobel, 1982). Stain uptake only occurs in living cells (Nobel and De la Barrera, 2003).
Five leaves per treatment were cut; we choose the middle leaves of the rosette, tissues samples were wrapped in moistened tissue paper and aluminum foil to avoid desiccation. Samples were exposed to air temperatures from 5 ºC to 65 ºC, with 5 ºC increases, for one hour. Temperature was monitored using a copper-constantan thermocouples in contact with the leaf sample. Low temperatures were obtained in a freezer, intermediate temperatures were obtained using a growth chamber and high temperatures were obtained with a heat dry incubator (EI40-AID, NOVATECH, Guadalajara, Mexico).
After the exposure to temperature treatments, samples were sliced with a razor blade, and were placed in 0.01 % w/v neutral red in distillate water during ten minutes and washed for ten minutes in distillate water. Then 130 intact chlorenchyma cells were examined at 100× using a BH-2 phase-contrast light microscope (Axioplan - Zeiss, Germany) to check for stained cells. The Lethal low-temperature (LLT50) and lethal high temperature (LHT50) were defined as the lower and higher temperatures, respectively, which halved stain uptake from the maximum occurring for the control at 25 °C (Didden-Zopfy and Nobel, 1982).
Statistical analysis. A two-way analysis of variance (ANOVA), followed by Tukey’s test were used to test differences between sites and sizes. Arcsine square root transformation was used to achieve normality (Sokal and Rohlf, 1969). Drought tolerance was measured as the number of days needed for the 50 % of the leaves to pass to the next phase of wilting. Statistical analyses were performed using STATGRAPHICS Centurion XV, version 15.2.06 (StatPoint, Inc.).
Results
Drought tolerance. After 61 days of drought, 100 % of the leaves of the smaller plants (S1) from Ria Lagartos were dead (Fig. 1a). At the same time, for San Benito individuals 95 % of the leaves were dead (Fig. 1b). The first responses to drought were observed at the ninth day of drought for plants from Ria Lagartos and for plants from San Benito, one day later. Both spent most time in the normal stage without showing any wilting responses and they rapidly progressed through the other wilting stages. For plants of Ria Lagartos and San Benito 50 % of the leaves reached the dead stage after 43 and 46 days, respectively.
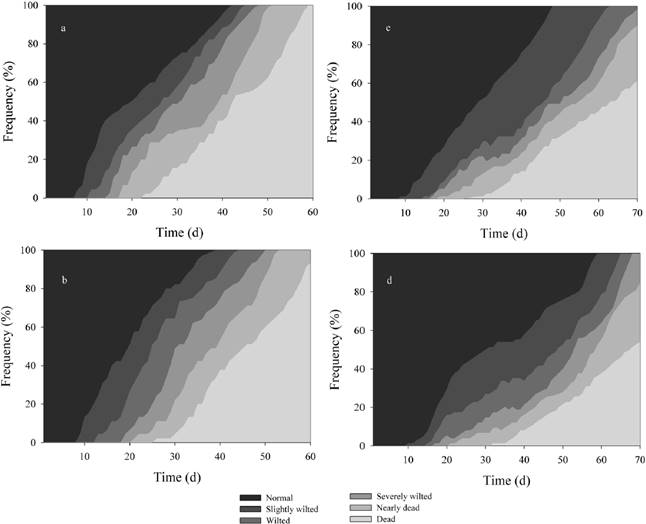
Figure 1 Time course of wilting of Tillandsia dasyliriifolia, on left small individual from Ria Lagartos a), form San Benito b). On the right larger plants form Ria Lagartos c) and from d) San Benito. Growing in controlled conditions, 22.57 mol m-2 d-1 PPFD, 33.02 ± 0.49 / 23.12 ± 0.66 ºC diurnal/nocturnal temperature (n = 10 per treatment). Different shading refers to different wilting stages.
For S2 plants, about 50 % of the leaves were dead after 65 and 67 days of drought for Ria Lagartos and San Benito respectively. In general plants passed more gradually from the normal stage to the slightly wilted stage, but San Benito plants passed more time in the wilted stage than Ria Lagartos plants, after the wilted stage they process was faster (Fig. 1c, d). We found differences between plant sizes but not between sites (P < 0.05, Fig. 1). However, there was a 2.5 ºC difference in tolerance between plant sizes (P < 0.05).
Leaf measurements. Tillandsia dasyliriifolia leaves had an average thickness of 0.95 ± 0.031 mm. The leaf blade is equifacial and it consists of epidermis, sclerenchyma, vascular bundles, chlorenchyma and hydrenchyma. As we expected the hydrenchyma tissue occupied almost of the 70 % of the area of the T. dasyliriifolia blade and a 31 % were chorenchyma (P < 0.05).
Extreme temperature tolerance. Extreme low temperature was similar for smaller plants (S1), lethal low temperatures (LLT50) were 11.95 ± 0.064 ºC for San Benito and 11.92 ± 0.047 ºC for Ria Lagartos. A strong reduction on the uptake of the stain was observed when tissues were exposed at 15 ºC and a decrement of the 93 % on the uptake occurred at 5 ºC. Lethal high temperatures (LHT50) for S1 plants were similar for both sites, 40.85 ± 0.095 ºC (P > 0.05).
For larger plants (S2) the LLT50 was 9.125 ± 0.047 ºC for San Benito and 9.15 ± 0.288 ºC for Ria Lagartos. In contrast the LHT50 showed for S2 plants was higher in comparison with S1 plants, 43.2 ± 0.081 ºC and 43.1 ± 0.057 ºC, San Benito and Ria Lagartos respectively. The LLT50 and LHT50 were similar between sites.
Discussion
Differences in drought tolerance have been found in different life stages in some bromeliads (Benzing and Dahle, 1971). The primary reason of this could be the anatomical advantages related to plant size, such as larger tank. For example for Catopsis sessiliflora (Ruiz & Pav.) Mez, an epiphytic bromeliad, exhibit a strong correlation between tank water capacity plant sizes. Larger individuals can withhold more water than smaller ones (Zotz and Laube, 2005). Also, the time in which the tank is empties is related to plant size. Smaller individuals of Tillandsia fasciculata Sw. and Guzmania monostachya (L.) Rusby ex Mez passed more time without water in the tank than larger individuals (Zotz and Thomas, 1999).
The differences related to the plant size were observed in early ontogeny. For example, six-month old germinated seedlings of Tillandsia flexuosa Sw. exhibited more tolerance than recently germinated seedlings (Bader et al., 2009). In some bromeliads was observed an inverse relation of rate of water loss with plant size. Smaller plants of Guzmania monostachya (L.) Rusby ex Mez loss more water by transpiration than larger plants. However to Tillandsia fasciculata Sw. water loss if not affected by the plant size (Zotz and Andrade, 1998).
Other advantage against drought and desiccation is the amount of hydrenchyma tissues (Barcikowski and Nobel, 1984). For example, after 50 days of drought the metabolizing region of Tillandsia ionantha Planch. leaf not shows a strong desiccation, allowing a positive assimilation of CO2; consequence of water movement from the hydrenchyma to the chlorenchyma (Nowak and Martin, 1997). Moreover, the presence of hydrenchyma in Tillandsia usneoides allowed an increase of 700 % in tissue water content after a rain and a reduction of 300 % after a severe drought (Penfound and Deiler, 1947).
Furthermore, a dense layer of trichomes have the inverse effect in water loss; they help regulate leaf temperature and transpiration through increased light reflectance (Ehleringer, 1984; Ohrui et al., 2007). Low stomatal density and crassulacean acid metabolism (CAM, Martin, 1994; Ricalde et al., 2010) are other adaptations of Tillandsia dasyliriifolia to arid environments to avoid desiccation.
As a general rule, low temperatures are used to explain species abundance in colder regions mainly because of damage by frost (Zotz, 2005). Low temperature causes change in carbohydrate metabolism, hormonal regulation and inhibit or decreases plant growth (Nievola et al., 2005; Guy et al., 2008; Hua, 2009; Mollo et al., 2011). Only a few bromeliads can tolerate low temperatures. For example, some Puyas and Tillandsia usneoides can survive and grow in cold regions such as northern USA (Benzing, 1980; 2000). In this way is not surprising that Tillandsia dasyliriifolia individuals were less tolerant to low temperatures than high temperatures. Low lethal temperature (LLT50) for adult individuals was similar to the LLT50 of Mammillaria gaumeri (Britton & Rose) Orcutt seedlings, a rare cactus endemic to northern Yucatan; acclimate at air temperature of 30/20 ºC (Cervera et al. 2006).
High temperature in epiphytes alters cellular and metabolic processes damages cell membranes, inhibiting of chlorophyll synthesis and affects chlorophyll degradation, reduce biomass and flowering furthermore some of them exhibit low acclimation (Vaz et al., 2004; Pisani et al., 2007). Like Vriesea ‘cathy’ an epiphytic bromeliad shows a decrease in growth, when the temperature increases 5 ºC (Pelligrini, 2012).
Compared with other succulent plants, such as agaves and cacti that growth in North American deserts, which can tolerate temperatures below 0 ºC and above 65 ºC, Tillandsia dasyliriifolia exhibits a moderate tolerance temperature (Nobel and De la Barrera, 2003; Nobel and Zutta, 2007; Drennan, 2009). Even the LHT50 of Hylocereus undatus (Haw.) Britton & Rose, a co-ocurring hemiepiphytic cactus is higher, similar Mammillaria gaumeri which exhibits a LHT50 above 5 ºC (Nobel and De la Barrera, 2002; Cervera et al., 2006).
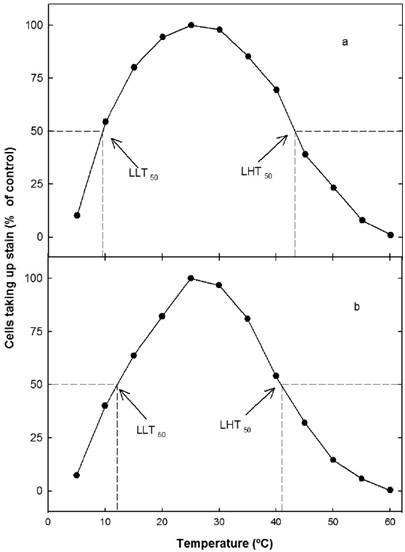
Figure 2 Effect of high and low temperature treatments on cell viability of Tillandsia dasyliriifolia adult larger (a) and smaller (b). Acclimated in controlled conditions to a diurnal/nocturnal temperature of 30/20 ºC and 50 % of total daily PPFD. Data are expressed relative to the control at 25 ºC (for which 100 % of the chlorenchyma cells took up stain).
Our results suggest that Tillandsia dasyliriifolia exhibited a moderate tolerance to drought and extreme temperatures. We confirmed the effect of plant size on drought tolerance and demonstrated that extreme temperature tolerance is also affected. Although differences in precipitation and microclimate of populations of origin were not significant, it is recommended that more studies with populations with more marked differences. Future research must focus on the responses of acclimation with respect to rising air temperatures and changes in the precipitation regiments accompanying global climate change.

