











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Mexico has a wide diversity of avocado fruit type, with at least 20 different species related to Persea americana C. K. Allen (Barrientos-Priego & López-López 2000). Persea hintonii is a wild relative of the avocado and an endemic species of Mexico, associated with the montane pine-oak and cloud forests on steep slopes. Successional patterns in these forests include the establishment of some subtropical montane cloud forest (MCF) species in moister and fertile soils under the canopy of subtropical pine-oak forest (POF) in conditions of prolonged absence of fire (Sánchez-Velásquez & Moya 1993, Jardel-Peláez et al. 2004, Saldaña-Acosta 2001). This pattern is important because species pioneering from MCF to POF potentially will have a better chance of migrate and survive in the current climate change conditions. Although many P. hintonii fruits fall under the mother tree at maturity, the birds and bats are primary seed dispersers of this species from the MCF to POF areas (Rost et al. 2015). However, high post-dispersal seed predation of P. hintonii in the POFs (Saldaña-Acosta 2001) could reduce the recruitment of new individuals and limit the propagation of this species to new areas.
After primary dispersal, the fruits or seeds can be predated or can be subject to secondary dispersal by different animal species. Secondary dispersal of fruits or seeds may end in their predation when they are consumed, or they may escape predation when they are forgotten or thrown away during transport by consumers (Dalling 2002). Although post-dispersal seed removal by vertebrates can result in a secondary favorable dispersion (López-Barrera & Manson 2006, Gómez et al. 2008, Perea 2012), the randomness of it, and the low percentage of seeds escaping predation will imply a high cost for fitness of some plant populations (Holl & Lulow 1997, Flores-Peredo et al. 2011). The basis for describing seed removal patterns is through the Janzen-Connell hypothesis, which predicts that seed predation will be higher in areas with a higher seed density close to conspecific, with respect to seed dispersed far away from the parental plant (Janzen 1970, Connell 1971). This model presupposes that the areas with high seeds density close to conspecific tress are an attractant for seed predators. Pre-dispersal and post-dispersal seed removal patterns can play an important role in plant population dynamics concerning demography, regeneration, and propagation in the community (Yu et al. 2014, DeMattia et al. 2004, García-Hernández et al. 2016). For plant species with an efficient dispersal mechanism, post-dispersal seed removal can be the main drawback factor influencing plant distribution, abundance, and successional patterns (Orrock et al. 2006). On other hand, for species with big and fresh fruits dispersed through gravity, the secondary dispersal by different animal species can facilitate the fruits/seeds to move away from the parent tree towards sites that are suitable for germination and seedling establishment (Vander Wall et al. 2005). Several factors such as seed or fruit eater preferences, seed attributes and habitat structure interact to define the pattern of seed/fruit removal in an ecosystem (Sánchez-Cordero & Martínez-Gallardo 1998, Dalling 2002, Doust 2011).
Fruits and seeds are an important seasonal food resource for a great variety of vertebrate species (Crawley 2014). Mammals are considered an efficient group in terms of post-dispersal fruit/seed removal, as they typically consume a high number of fruits and seeds (Holl & Lulow 1997, Sánchez-Cordero & Martínez-Gallardo 1998). Previous research has identified small mammals (mainly rodents) as the main fruit/seed predators in neotropical forests (Wenny 2000, DeMattia et al. 2004, García-Hernández et al. 2016) and temperate forests (Kamler et al. 2016). Habitat structure and vegetation type are important factors influencing removal of fruit or seeds by vertebrates. In general, it has been reported that in comparison to areas without vegetation cover, seed removal of broadcast sowing is higher in areas with a higher plant density (Doust 2011). Concerning some small vertebrate species, the existence of a canopy or an understory with dense shrub cover can trigger an increase in seed removal rates, compared to those in open areas (Pons & Pausas 2007, Fanson et al. 2008, Perea 2012, Yu et al. 2014, García-Hernández et al. 2016). Selection of sites with dense vegetation cover can provide an environment with a reduced remotion or predation risk for some fruit-seed consumers (Hixon 2003, Doherty et al. 2015), which allows them to allocate more time to search and consume food in those sites (DeMattia et al. 2004, Fanson et al. 2008). It is known that many species of rodents and birds feel safer and can forage more extensively under or close to a bush cover than in open microhabitats or in areas that are at a longer distance from such cover; they are aware of the risk of being hunted by aerial predators (Brown 1988, Embar et al. 2014). For fruit/seed consumers, therefore, a trade-off between time and energy exists between the amount and quality of food and the risk of predation.
The subtropical MCF is one of the most diverse ecosystems in Mexico; its distribution is fragmented and discontinuous, and the tree species occurring in this vegetation type are generally present in low densities (Santiago-Pérez et al. 2009, Cuevas-Guzmán et al. 2010). In this context, it is important to evaluate how fruit/seed post dispersal removal could impact the propagation of MCF species. The spatial pattern of diaspores’ removal throughout the landscape can play an important role in the seedling establishment patterns of MCF species. We hypothesized that (i) due to the small seed size, the main post-dispersal fruit removers will be small and medium vertebrate, either mammals or birds. (ii) According to escape hypothesis (Janzen 1970, Conell 1971), post-dispersal fruit removal of Persea hintonii will be higher in areas close to individuals fruiting at MCF, compared to more distant sites with no conspecifics fruiting at POF or OA. And (iii) regardless of the type of vegetation, areas with higher vegetation density will be associated with a higher removal of P. hintonii fruits. The objective of this study was to determine the influence of vegetation density and vegetation type in the fruit removal patterns at potential areas for new individuals’ establishment and to identify the most frequent removers of P. hintonii fruits in MCF. We discuss the role of possible predation risk for fruit-eating vertebrates in the removal pattern. This study will increase our understanding of fruit/seed removal as part of the successional patterns in subtropical forests, to identify sites with a lower risk of seed predation, as well as its possible implications for management and conservation.
Materials and methods
Study area. Las Joyas Scientific Station (LJSS) is part of the Sierra de Manantlán Biosphere Reserve (SMBR) at 19° 34' 14''; 19° 37' 30'' N; 104° 14' 49'', 104° 18' 16'' W, in western Mexico. It has a territory of 1,257 ha and an altitudinal range from 1,560 to 2,242 m asl. The climate is temperate-subhumid with a warm summer (Martínez-Rivera et al. 1991); annual average temperature is 15.72 ± 1.52 °C (average ± sd), and annual average rainfall is 1758.16 ± 203.42 mm (Zuloaga-Aguilar 2021). The vegetation in the study area are subtropical forests; LJSS is mostly covered by pine-oak and pine forests, and a small proportion (18 %) of the area is MCF. In the area, the MCF grow in guillies (moist and fertile soil) and the pine-oak and pine grow in midslopes and upper slopes (low moisture and poor fertility soils) (Cuevas & Jardel 2004). The most frequent species in the MCF canopy are Dendropanax arboreus (L.) Decne. & Planch., Carpinus caroliniana Walter, Aiouea pachypoda (Nees) R. Rohde, Persea hintonii, Magnolia iltisiana A. Vázquez and Quercus nixoniana Valencia y Lozada. In the understory, Solanum nigricans M.Martens & Galeotti, Euphorbia schlechtendalii Boiss and Parathesis villosa Lundell are fairly common. The dominant species in POF are Pinus douglasiana Martínez, Quercus candicans née and Q. scytophylla Liebm; Arbutus xalapensis Kunth is also present. The POF understory contains saplings of MCF trees and shrubs of Solanum aphyodendron S.Knapp, Cestrum aurantiacum Lindl and Parathesis villosa Lundell as well as several species of Salvia, among others (Jardel-Peláez et al. 2004). Small areas of abandoned agriculture (˃ 30 yr) persist in the POF. These open areas (OA) are mainly covered by shrubs, grasses, and, at very low densities, seedlings of pioneer shrub and tree species such as Budleja parviflora Kunth, Senecio salignus DC, P. hintonii and P. douglasiana. Persea hintonii is a dominant species in the MCF; however, its seedlings can be established in sites with moister and fertile soils under the POF canopy. The fruit of P. hintonii is an oval drupe (11.7 × 11.4 mm), with a dry weight of 0.41 ± 7-03 g, thin green exocarp (≈ 1 mm thickness) and a mesocarp and endocarp are almost absent. The seeds of creamy color; the main nutrients of the scarce flesh had lipids accounting for 1.9 % of the dry weight, and proteins are 0.086 µg/mg of the fruits (Rojo-Cruz 2016). Due to the fruit size of P. hintonii, many of the fruits fall to the ground by gravity as they ripen, but birds and bats are also dispersers of this species from MCF to POF (Rost et al. 2015). The most abundant medium and large mammals in the study area are the collared peccary (Pecari tajacu Linnaeus), Virginia opossum (Didelphis virginiana Kerr), white-tailed deer (Odocoileus virginianus Zimmermann) and puma (Puma concolor Linnaeus) (Moreno-Arzate 2009). In the case of small mammals, rice rats (Oryzomys couesi Alston), deer mice (Peromyscus spicilegus J. A. Allen), harvest mice (Reithrodontomys fulvescens J. A. Allen), Allen’s woodrat (Hodomys alleni Merriam) and Allen’s cotton rat (Sigmodon alleni Bailey) are among the most abundant species (Vázquez et al. 2000). All vertebrate species used different vegetation types in the area.
Record of seed consumers. To identify the vertebrate species that remove post-dispersal P. hintonii fruits in the MCF, trap cameras (either Bushnell Trophy 119537 or Moultrie MCG-12631) were set up under the canopy of three P. hintonii trees separated by ≈ 100 m, with a high number of ripe fruits (one camera per tree, n = 3). Under each tree, the camera was set 30 cm above the ground, with the focal point on where most of the fresh fruits (≈ 100) get accumulated under the tree canopy. The creation of photo records was carried out during the fructification peak of P. hintonii (15 March to 15 April 2015). Cameras were set up to operate 24 hr with an interval of one minute lapse between each photo. The definition of independence in photo-records varies among studies; in activity patterns and abundance studies, the most common interval ranges from 0.5 h to 1 h (Linkie & Ridout 2011, Harmsen et al. 2011, Wearn & Glover-Kapfer 2017). Due our setting configuration, we were able to evaluate the performance of different periods to consider an event as independent (30 minutes, 12 hours, and 24 hours), to identify the differences caused by the specific activity for different animal species. The relative frequency of visits was calculated using the relative activity index
Where IPR is the independent photo-records number for the species, and SE is the sampling effort (total days with the trap camera in operation) (O’Brien 2011). In addition, during the fructification period, all scats within a 10 m radius from the focal point were collected every day to search for P. hintonii seeds. Each mammal excreta were identified based on a field guide (Aranda-Sánchez 2012).
Exclusion treatments. To determine the significance of different-sized animals on post-dispersal fruit removal, we carry out an experiment using exclusion cages built with 2 cm wire mesh (see Supplementary material, Figure S1). Three different exclusion treatments were used in relation to the possible size of the removers: 1) the exclusion of medium and large round drupe removers, mostly mammals, allowing access for small removers such as rodents (access of 5 cm height × 25 cm width and an area of 35 × 25 cm; treatment name: Small); 2) the exclusion of large removers, allowing access for small- and medium-sized animals (access of 35 cm height × 30 cm width and an area of 60 × 45 cm; treatment name: Medium = small + medium); and 3) without exclusion, allowing access to all removers (i.e., treatment name: Control, or without exclusion, = small + medium + large).
To evaluate the effect of the vegetation type and density on fruit removal by vertebrates, experiments were conducted in two forest types, avoiding to locate the treatments under fruiting trees of any species: MCF and POF, with two understory conditions, namely dense (du, ˃ 30 %) and sparse (su, ˂ 10 %), and open area of abandoned crop (OA) with only one condition: dense shrub cover with no tree canopy. These vegetation types represent potential areas for the establishment of new P. hintonii individuals through fruit dispersal. For each forest type (n = 2), cover condition (n = 2), and OA, we selected two sites that are at least 100 m apart (total n = 10 sites). Vegetation cover density in each site was described as the average number of individuals/m2. Canopy trees were recorded in one circular 500 m2 plot (a radius of 12.61 m); the understory shrubs and saplings (< 5 cm DBH) were recorded in three 16 m2 plots; and the cover of grasses and herbaceous plants (including seedlings) was recorded as percentage in three 1 m2 plots.
In each site, four replicates of each exclusion treatment (small, medium, and control) were established. These were distributed randomly in a grid pattern with a minimum distance of 20 m between any two points (total n = 120 points). The exclusion cages were fixed to the ground with stakes; before, we eliminated the litter at each point. The fruits were deposited on flat paper dishes in each exclusion cage (small and medium = 17-25 fruit/cage, control = 8-50 fruit/cage). The number of fruits per experimental unit varied along the fruiting season according to the availability of fresh and mature fruit (see Supplementary material, Table S1). As the fruits do not ripen at the same time, the fresh fruit were collected daily to ensure that all the exclusion plots were sufficiently baited for three days. Each exclusion was baited only once. All of the experiments were carried out at the end of the dry season, and the number of fruits removed in each experimental unit was recorded each day for 15 days (28 April to 12 May of 2015). We estimated the uniformity index (UI) of fruit removal based on a germination uniformity coefficient (see González-Zertuche & Orozco-Segovia 1996) using the following
Statistical analysis. To find the differences in the visits’ frequency (photo-records) caused by specific activities of fruit-removing species, accounting 30 minutes, 12 hours, or 24 hours among independent records, we carried out a non-parametric analysis of longitudinal data (NPFALD) and ANOVA-type tests (ATS) using the function ex.f1f1$Wald.test (Brunner et al. 2002). For the next tests, we selected the time interval with the highest relative frequency of occurrence. The differences in the relative frequency of visits (the number of photo-records in three cameras) among P. hintonii fruit-removing species (n = 8 species) were evaluated with a non-parametric ANOVA (Kruskal-Wallis test, P ≤ 0.05). Multiple comparison tests among treatments were carried out with the function posthoc.Kruskal.conover.test in a Kruskal-Wallis test, which includes a Bonferroni correction (Pohlert 2014).
Because the number of fruits per replicate was different, we performed a mixed model to include this variable as the random factor. One-way linear mixed model with binomial error was carried out to evaluate differences in the number of fruits removed among vegetation types (MCF, POF and OA) for each exclusion treatment (Small, Medium and Control). The fixed and random effects variables were the vegetation type and number of fruit, respectively. To assess the effect of the understory conditions (dense and sparse) in the two forest types (two sites in MCF and POF) on the number of fruits removed, for each exclusion treatment (access to animals, only small, small + medium, or without exclusion), a generalized two-way linear mixed-effects model (GLMM) with binomial error (binary response) was performed. The fixed effects variables were the understory condition*forest type + sites, and the random effect variable was the number of fruits by day. The GLMM was performed with the function glmer on the lme4 package (Bates et al. 2015). The normal distribution of the model’s residuals was revised, and a Type III Wald chi-square test was performed.
Linear regressions were performed to evaluate the relationship between the vegetation density (saplings + shrub + herbs) and fruits removal proportion and uniformity index of fruits removal. Compliance with the assumptions of normality of distribution and homogeneity of variance were verified prior to the conduction of the regression tests. The effect of vegetation type (forests type + understory, n = 5) on the time delay until the removal of first fruits (starting time) and uniformity index of fruit removal was evaluated using a Kruskal-Wallis test (P ≤ 0.05) for each exclusion treatment type. Multiple comparison tests among treatments were performed with the function posthoc.kruskal.conover.test in a Kruskal-Wallis test, which includes a Bonferroni correction (Pohlert 2014). All statistical analyses were conducted in R v. 3.2.1 software (R Core Team 2015).
Results
Fruits removal pattern. In general, the percentage of Persea hintonii fruits removed was higher in MCF, compared to POF and OA (70.10 ± 3.34 %, 11.98 ± 1.63 %, and 9.46 ± 6.56 %, average ± SE, respectively). Vegetation types significantly affected the P. hintonii fruits removal in the Small (χ2 = 51.42, d.f. = 2, P = 6.79-12), Medium (small + medium) (χ2 = 57.95, d.f. = 2, P = 2.60-13), and Control (small + medium + big vertebrates) (χ2 = 28.21, d.f. = 2, P = 7.45-07) treatments (Figure 1). For all three exclusion treatments, the percentage of P. hintonii fruit removal was influenced by the interaction between the understory presence and forest type. The highest percentage of fruits removal by small vertebrates was recorded in the MCF, either with a dense or sparse understory (79.06 ± 0.6 % and 67.08 ± 14.43 %, respectively), and in POF with dense understory (20.62 ± 7.51 %) (Figure 1A; Table 1A). Regardless of the understory condition, fruit removal by small and medium vertebrates was more than five times higher in MCF (MCFdu = 73.50 ± 1.06, MCFsu = 50.50 ± 8.03) compared to POF (POFdu = 15.05 ± 2.87 %, POFsu = 1.00 ± 0.70 %) (Figure 1B, Table 1B). Similar results in fruit removal were observed among forest types from areas without exclusion, where all of the vertebrates could access the fruits (Figure 1C, Table 1C). In general, fruit removal in the OA was low; however, the fruit removal recorded in the Control treatment was higher compared to fruit removal by small and medium vertebrates. As a result of this analyses, the variation associated with the number of fruits per experimental unit was ˂ 1 (random effect); therefore, the number of fruits removed not vary in relation to the number of fruits per exclusion.
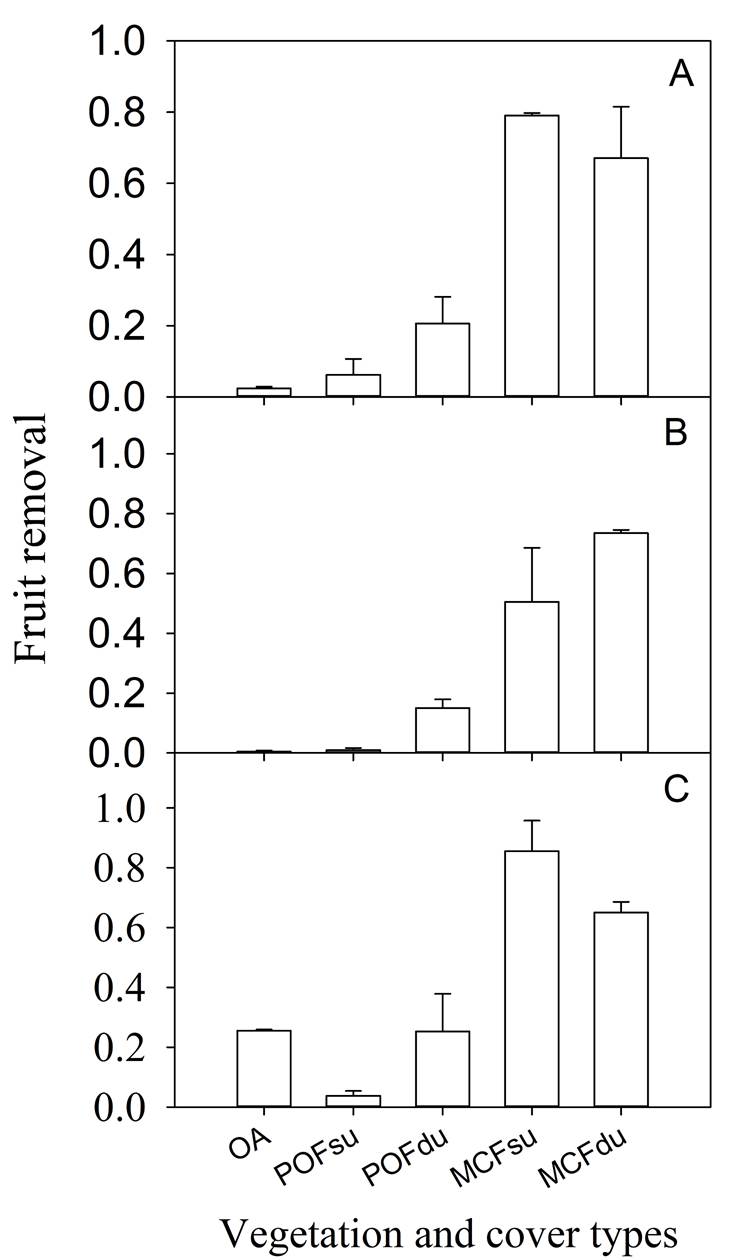
Figure. 1 Proportion of Persea hintonii fruits removal by (A) small, (B) small + medium and (C) all vertebrates without exclusion in five vegetation and understory cover types. MCFdu = montane cloud forest with dense understory, MCFsu = montane cloud forest with a sparse understory, POFdu = pine-oak forest with a dense understory, POFsu = pine-oak forest with a sparse understory, and OA = open area with dense shrub cover (four replicates by exclusion treatment and vegetation and cover type). The whiskers represent the SE in each treatment.
Table 1 Analysis of deviance (Type III Wald chi-square tests) for two-way linear mixed models with binomial error (binary response) to test the effect of understory condition and forest type (MCF and POF) on the amount of Persea hintonii fruit removal in Small (A), Medium (small+medium) (B) and Control (small + medium + big) (C) treatments.
| Treatment | χ2 | df | Pr(>χ2) |
|---|---|---|---|
| A | |||
| Vegetation Type (VT) | 27.47 | 1 | < 0.0001 |
| Understory (U) | 0.64 | 1 | 0.42 |
| U*VT | 6.44 | 1 | 0.01 |
| B | |||
| Vegetation Type (VT) | 73.57 | 1 | < 0.0001 |
| Understory (U) | 10.60 | 1 | 0.001 |
| U*VT | 6.37 | 1 | 0.01 |
| C | |||
| Vegetation Type (VT) | 76.58 | 1 | < 0.0001 |
| Understory (U) | 18.79 | 1 | < 0.0001 |
| U*VT | 6.03 | 1 | 0.01 |
In our study, the vegetation density of understory + canopy and P. hintonii fruit removal was found to be positively related. We showed that fruit removal increased in areas with a higher vegetation density (N = 10, F = 8.84, P = 0.018; Figure 2A). A similar pattern was observed in the uniformity index of fruit removal and vegetation density (N = 10, F = 30.92, P = 0.0005; Figure 2B); the uniformity of removal was higher in areas with greater vegetation cover. The starting time of P. hintonii fruit removal differed among vegetation types (χ 2 = 39.08, d.f. = 4, P = 6.68-08); it was shorter in MCFdu, MCFsu, and POFdu, where fruits removal began two and seven days earlier than it began in POFsu and OA (13.75 ± 0.49 and 11.70 ± 0.91 days, respectively; Figure 3).
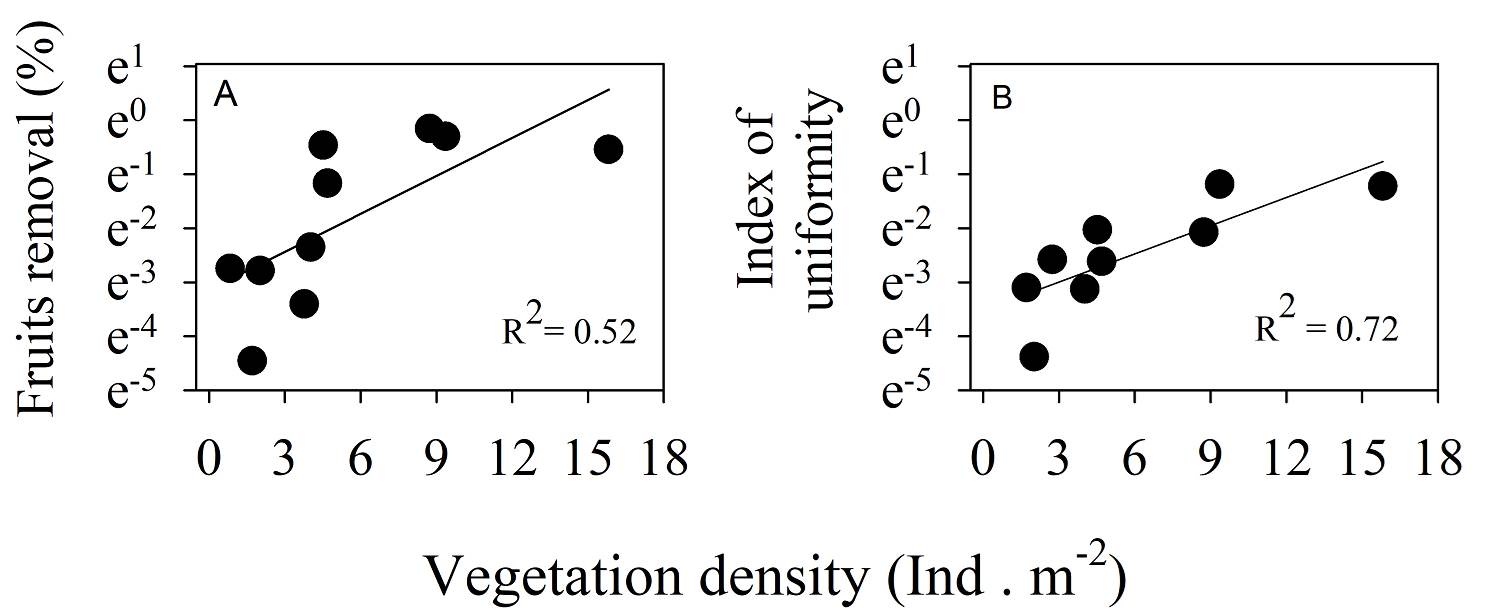
Figure. 2 Linear regression among the densities of vegetal cover and the percentage of Persea hintonii fruits removed A) and the index of uniformity (B) in LJSS, Jalisco, Mexico.
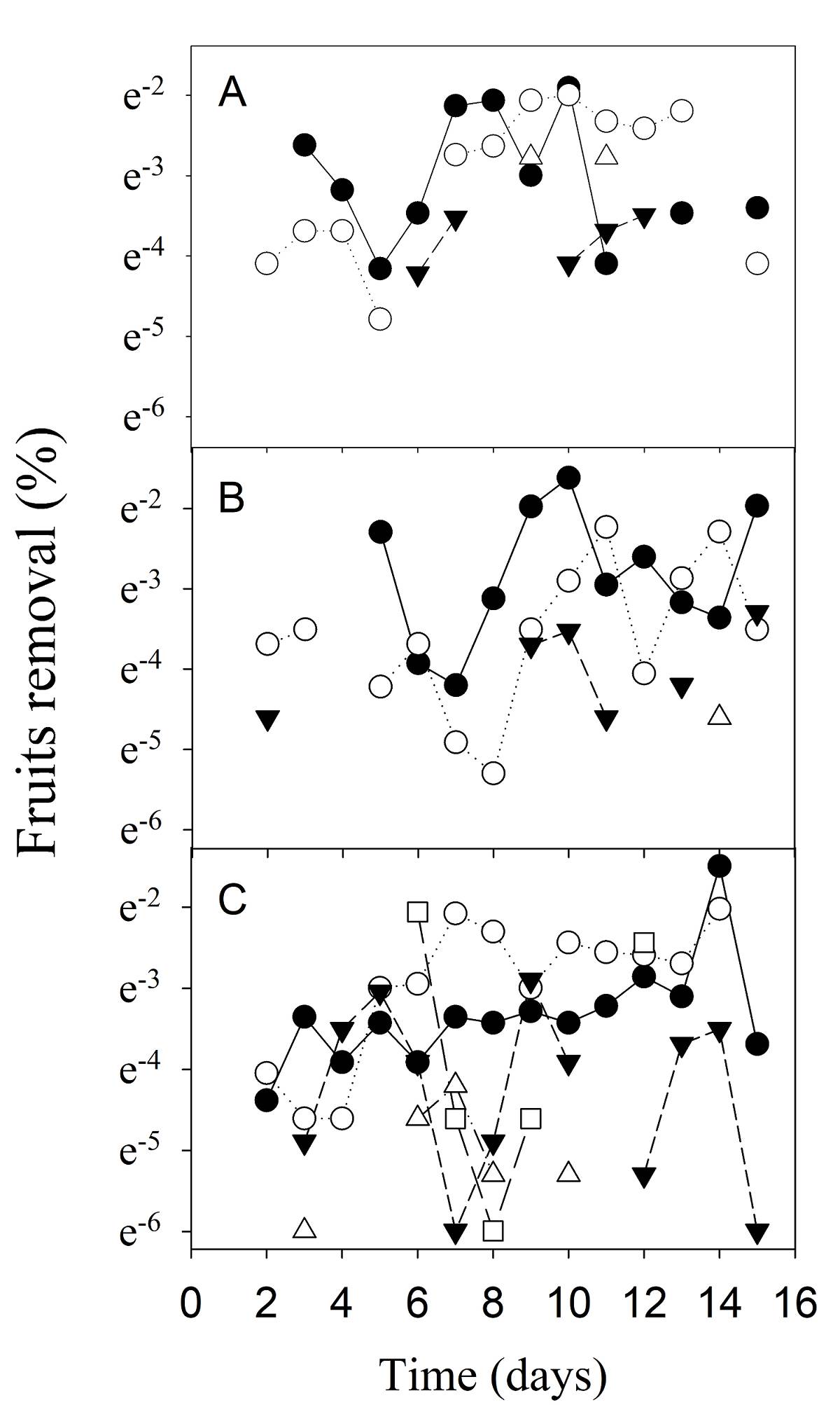
Figure. 3 Proportion of Persea hintonii fruits removed over the course of time in five vegetation plus understory cover types in LJSS, Jalisco, Mexico. Seed removal was performed through (A) Small, (B) Medium and (C) Control exclusion treatments (see text). MCF with dense understory = filled circle; MCF with a sparse understory = empty circle; POF with dense understory = filled triangle; POF with a sparse understory = empty triangle, and OAs with dense shrub cover = square.
In general, regarding the way fruit removal occurs over time, no uniform pattern was seen (Figure 3). Our result showed that in MCFdu, MCFsu, and POFdu, fruit removal by small consumers occurs continuously for a few days (0.36-0.20, uniformity index) as compared to the scattered removal in the POFsu and OA (< 0.07) (χ 2 = 10.9, d.f. = 4, P = 0.02; Figure 3A). The small+medium consumers removed more fruits in a shorter timespan in the MCFdu (0.41), compared to the other vegetation types (<0.12) (χ 2 = 27.4, d.f. = 4, P <0.0001; Figure 3B). The remotion of fruits without exclusion occurs continuously over time in MCFdu, MCFsu (0.33-0.21, respectively); however, more than 20 % of the fruits were removed in one day in MCFdu. Fruits removal was discontinous in the OA without exclusion, where 20 % of the P. hintonii fruits were removed only in two different dates (Figure 3C).
Fruits consumers on MCF. We identified eight vertebrate species as the removers of P. hintonii fruits on MCF (Figure 4). Our results showed that visit frequencies of the species differed depending on the photo-record interval analyzed (30 min, 12 h, and 24 h) (χ2 = 300.82, d.f. = 11, P = < 0.0001). With the exception of Neotoma mexicana, the visit frequency of each species decreased with an increase in the photo-record interval (χ2= 59.09, d.f. = 2, P = < 0.0001). Regardless the photo record interval (30 min, 12 h, and 24 h), the visit frequency differed among species (χ2= 379.9, d.f. = 7, P = < 0.0001). The 75 % of the fruits were removed by mammal species, out of which Pecari tajacu and Odocoileus virginianus are large size mammal, Didelphis virginiana and Sciurus colliaei are medium size, while Neotoma mexicana and Oligorizomys fulvescens are small. The collared peccary (P. tajacu) was the most frequent fruit remover (RAI30 min= 134.44 ± 25.59), followed by Virginia opossum (D. virginiana; RAI30 min= 77.8 ± 8.67). Mexican woodrat (N. mexicana and pygmy rice rat (O. fulvescens) were the least frequent (both with RAI30 min= 4.44 ± 2). Regarding birds, nightingale-thrushes (Catharus spp.) and green-striped brush finch (Arremon virenticeps) were also identified as important fruit removers (RAI30 min= 37.8 ± 7.7 and RAI30 min= 10 ± 1.9, respectively). During the experiment, only one excreta was recorded, belonging to crested guan (Penelope pupurascens), which contained 5 seeds of P. hintonii, but no photo-records of this bird were recorded.
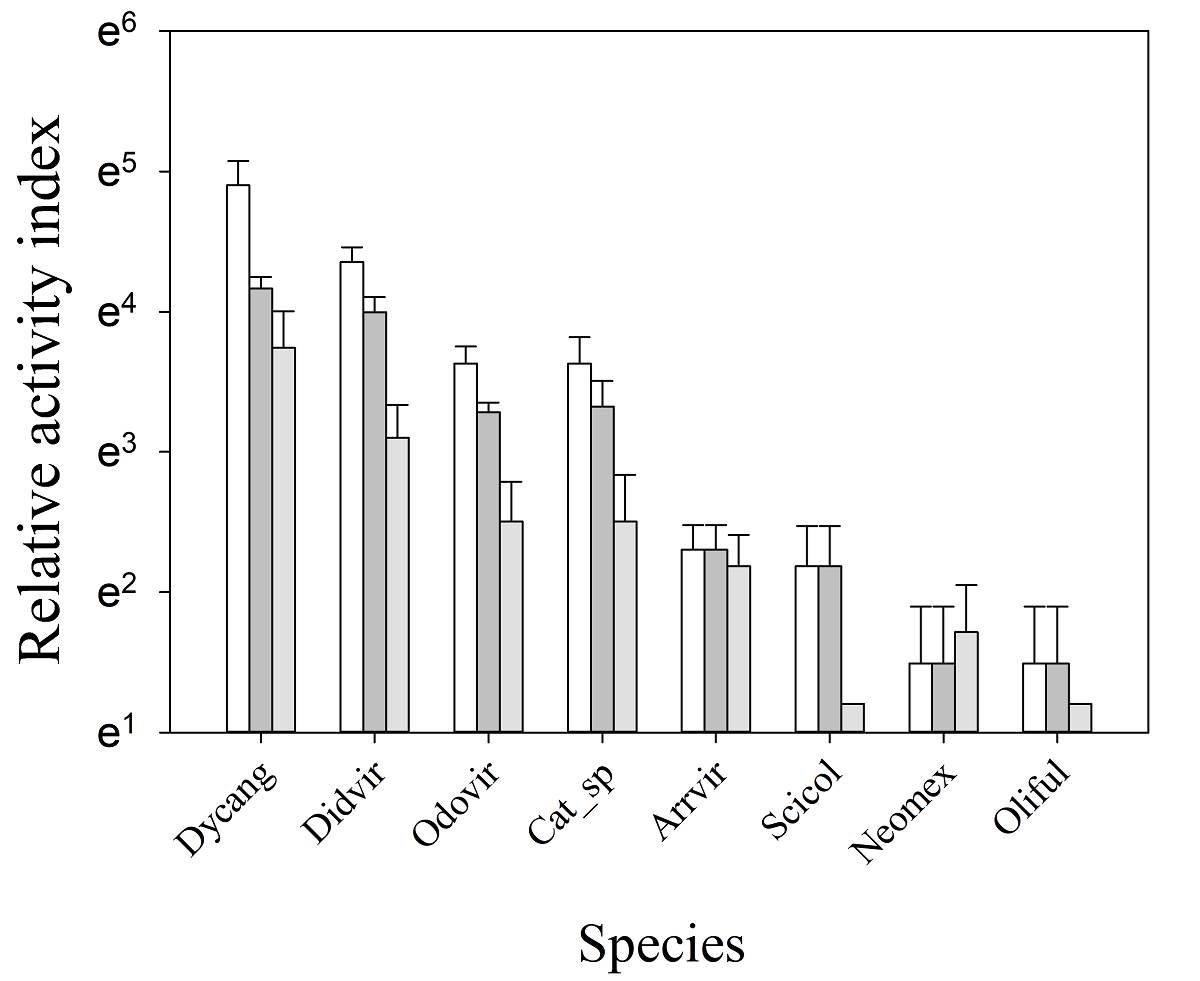
Figure. 4 Average relative activity index to different independent photographic record intervals of Persea hintonii fruit-removing species in the subtropical MCF of LJSS, Jalisco, Mexico. The whiskers represent the standard error (SE) in each treatment. Bars white = 30 min, bars gray dark = 12 h and bars gray = 24 h. Dicang = Dicotyles angulatus, Didvir = Didelphis virginiana, Odovir = Odocoileus virginianus, Scicol =Sciurus coliaei, Neomex = Neotoma mexicana, Oliful = Oligorizomys fulvescens, Arrvir = Arremon virenticeps, Cat_sp = Catharus spp.
Discussion
The percentage of removal of Persea hintonii fruits by vertebrates is high in subtropical MCF (50-85 %), and like reported values for other species in tropical forests (Holl & Lullow 1997, Blendinger & Díaz-Vélez 2010, García-Hernández et al. 2016). Based on the photographic records, large- and medium-sized mammals, such as Pecari tajacu, Odocoileus virginianus and Didelphis virginiana, are the more frequent consumers of P. hintonii fruits in the MCF. However, the results of the fruits removal exclusion treatments show that small-sized vertebrates can also be important removers of P. hintonii fruits (65-70 %). Even though we fixed trap cameras at 30 cm off the ground, it is possible that the camera-trap method underestimated the frequency of the appearance of small-sized vertebrates (Chávez et al. 2013). Therefore, small- and medium-sized mammals are also important removers of P. hintonii fruits, such as Oligorizomys fulvescens, Neotoma mexicana and Sciurus colliaei. The birds as secondary removers of P. hintonii fruits were less frequent than mammals; we only record two bird species in our sites. In other Lauraceae trees (such as Ocotea andresiana Mez), mice were also more frequent than birds as the main fruit/seed secondary removers (Wenny 2000). Our results do not allow us to determine whether P. hintonii fruits are predated or dispersed a second time. However, we can suppose that due to P. hintonii seeds doesn't have a hard testa (or endocarp), they are unlikely to survive consumption by mammals such as Pecari tajacu, Odocoileus virginianus, Didelphis virginiana and rodents. Therefore, most of the post-dispersal removal of P. hintonii fruits by the above-mentioned species, will probably result in predation. On the other hand, if during their movements, the birds Catharus spp., Arremon virenticeps and Penelope purpurascens regurgitate or defecate the removed fruits, they can facilitate the secondary dispersal of P. hintonii fruits. However, Catharus spp and Arremon virenticeps are omnivores, and probably they consume only occasionally the fruits of P. hiintoni in search of a specific nutrient. Therefore, these bird species could do not contributed significantly to post-dispersal remotion of P. hiintoni fruits. As an additional observation, the crested guan (Penelope pupurascens), although was not in the photo-records, can be considered a consumer of P. hintonii fruits, since seeds were found in their excreta. It is important to note that this species can act both as a pre-dispersal (foraging in the canopy) and post-dispersal (foraging on the ground) consumer. Fruit removal by ants has not been reported for P. hintonii, and during fruit collection or visits to the exclusions, no ant activity was observed at the sites. The seeds of this species do not have aril, so it is unlikely that they are removed by ants. Seed removal by ants has been reported for Persea indica (L.) Spreng (Naranjo 2010), however, in this work no insect repellent was used, so it is possible that some of the fruit removal from P. hintonii could be due to ant activity. The specific impact of insects as seed movers of P. hintonii should be investigated to design better management strategies for future propagation projects. Our third hypothesis is partially fulfilled, since in addition to medium fruit/seed eaters, large fruit eaters could be the main fruit post-dispersal removers of P. hintonii.
Regardless of whether fruit removers are small, medium, or big-sized mammal species, fruit removal of P. hintonii is higher in the MCF than in the POF. Our results show that the fruit removal pattern of P. hintonii is consistent with the Janzen-Connell hypothesis (Janzen 1970, Connell 1971), which predicted that fruits predation will be higher in areas with a higher fruits density, near conspecifics. The reproductive individuals of P. hintonii were mainly in the MCF. Our first hypothesis is fulfilled, therefore, the high fruits removal in this forest could be the result of predictability as a density-dependent response. High fruit densities near conspecific trees are an attractant for fruit predators. Our study showed that vegetation type and density interact related to the percentages of fruits removed and removal uniformity. In general, we found that increased vegetation density is related to higher uniformity in the fruits’ removal of P. hintonii by vertebrates. This pattern was clear in the POF, where areas with a sparse or no understory presented lower fruit removal than in a forest with a dense understory; these results confirm our second hypothesis. Similar results are reported for different species; the loss of seeds by broadcasting was higher in areas with higher vegetation cover (Doust 2011). On the other hand, for some fruit/seed consumer vertebrates, a complex microhabitat structure is necessary to avoid predation (Pons & Pausas 2007, Stevens & Tello 2009, Perea 2012). In areas with a more open vegetation structure, there can be greater predation risk and a consequent decrease in foraging time (Hixon 2003, Doherty et al. 2015). This behavior has been well documented for small rodents and bird species in areas where the main risk comes from aerial predators (Lima 1985, Brown 1988). Therefore, the low removal of P. hintonii fruits in the POF (particularly POFsu) and OA could be a result of the high risk of predation to which vertebrates are exposed. On the other hand, the lower removal of P. hintonii fruit in POF and OA, compared to MCF, may be due to the feeding preferences of the fruit/seed eaters. During the course of the experiment, the opening and dispersal of P. douglasiana seeds occurred, a food resource that is first preferred over P. hitonii fruits by some species of mice (Rojo-Cruz 2016).
It is important to note that in the OA, P. hintonii fruit removal was higher in the Control treatment, compared to the exclusion treatments. Fruits removal in OA may be associated with the biggest herbivorous mammals in the study area, such as the collared peccary, white-tailed deer or opossum. Some studies report that ungulates prefer to forage mainly in gaps and less into the forest, provided the gap allows long-range visibility to detect large predators (Reyna-Hurtado & Tanner 2005, Stears & Shrader 2015, Davies et al. 2016, Esparza-Carlos et al. 2018). Additionally, the environmental conditions negatively affect the composition and amount of seed reserves (Garcias-Morales et al. 2021). The P. hintonii seeds are of the recalcitrant type (RBGK 2020). It is highly probable that the environmental condition in OA, characterized by high temperatures and low relative humidity, could have altered the composition of seed reserves. Therefore, low seed removal in the OA may be associated with a decrease or loss of seed quality with respect to an attribute appreciated by consumers. Therefore, additional studies should be conducted to evaluate how the seed quality of P. hintonii can be another factor affecting seed consumers’ behavior.
Our study showed that P. hintonii fruits removal in pine-oak forest is lower compared to that in the montane cloud forest. In our study area, the seedling establishment of broadleaf species such as P. hintonii in the pine-oak forest is common under certain conditions (sites with wetter and fertile soils, and greater canopy openness in comparison of MCF). If the fruit removers do not damage the seed, reduced fruits removal in the pine-oak forest can, therefore, be a factor that facilitate the establishment of P. hintonii and other broadleaf species in this vegetation type. This behavior is probably indirectly mediated by the herbivores’ fear of predation. Based on the relative activity index, large fruits eaters are the main Persea seed removers in MCF. Our hypothesis was confirmed for P. hintonii: fruits removal by small- and medium-sized mammals is lower in areas with a sparse vegetation cover and a higher predation risk. Foraging patterns of fruits removers can, thus, have a direct implication on the potential success of natural regeneration and the successional process in the subtropical mountain plant communities. In the context of forests’ conservation and management, the use of exclusions is a technique that allows the evaluation on susceptibility of fruits to remotion by different types of vertebrates, and the selection of potential areas for their successful establishment in projects of assisted migration.
Supplementary material
Supplemental data for this article can be accessed here: https://doi.org/10.17129/botsci.3106

