











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
The quantitative study of ecosystems provides essential information for their thorough understanding and proper management and conservation (Seppelt et al. 2011, Viani et al. 2017, Valtonen et al. 2021). Nonetheless, current knowledge about practically every ecosystem on Earth is far from satisfactory. An attractive alternative to advance in this regard is offered by remote sensing, a highly relevant science that studies the biophysical features of the terrain through the analysis of data acquired by remote sensors (Chinea 2002, Turner et al. 2003, Aplin 2004, Navulur 2007, Schowengerdt 2007, Mabwoga & Thukral 2014, Valderrama-Landeros et al. 2018, Chuvieco 2020). Remote sensing has fostered the study of ecosystems mainly by focusing on their plant cover, which is their most conspicuous component (Rasolofoharinoro et al. 1998, Hansen et al. 2013, Mezaal et al. 2017, Abdel-Hamid et al. 2018, Einzmann et al. 2021). Numerous studies on various aspects of the planet’s vegetation through remote sensing have demonstrated the great potential of this discipline to identify, map and monitor plant communities and their attributes without having direct contact with the terrain (Chinea 2002, Aplin 2004, Xie et al. 2008, Wang et al. 2010, Hansen et al. 2013, Mabwoga & Thukral 2014, Song et al. 2016, Putut Ash Shidiq et al. 2017, Islam & Ma 2018, Einzmann et al. 2021).
Wetlands are among the most important and, at the same time, most critically endangered ecosystems of the world (Valiela et al. 2001, Islam 2010, Islam et al. 2014, Mabwoga & Thukral 2014, Al-Naimi et al. 2016, Mao et al. 2021). Climate change seriously threatens their persistence, and multiple human activities strongly impact them (Kovacs et al. 2009, Lee et al. 2014, Islam et al. 2014, Thomas et al. 2017, Cho & Qi 2023). Given the physical characteristics of their habitats and the peculiarities of the vegetation typical of these ecosystems, access to them is difficult, mainly when they cover extensive areas (Kuenzer et al. 2011, Sharma 2018). This difficulty explains to some extent the relatively low pace at which crucial information on their structure, composition, and conservation status is gathered; this is worrisome given their high rates of transformation and fast disappearance (López-Portillo & Ezcurra 2002, Flores Mejía et al. 2010, Hogarth 2007, Landgrave & Moreno-Casasola 2012, Steinbach et al. 2023). An additional property of wetland ecosystems is the high variation of their vegetation; from a practical perspective, such variation can be reduced to the distinction between those communities dominated by herbs (herbaceous wetlands) and those in which dominance is shared by several woody species (wetland forests; Figure 1), among which mangrove forests have received the most attention from vegetation ecologists (Moreno-Casasola et al. 2009, Infante Mata et al. 2011).

Figure 1 Images of tropical wetland forests in Mexico. Wetland forests encompass mainly mangroves but also other forests which are subjected to permanent or periodic flooding. (A) Mangrove forest dominated by Rhizophora mangle L. in Pantanos de Centla Biosphere Reserve, Tabasco. (B) Tintal (seasonally flooded forest dominated by Haematoxylum campechianum L.) in Nahá-Metzabok Biosphere Reserve, Chiapas. (C) Pukteal (seasonally flooded forest dominated by Terminalia buceras (L.) C. Wright), Pantanos de Centla Biosphere Reserve, Tabasco. (D) Wetland forest with tasiste palm (Acoelorraphe wrightii (Griseb. & H.Wendl.) H.Wendl. ex Becc.), Pantanos de Centla Biosphere Reserve, Tabasco. (E) Apompal (wetland forest dominated by Pachira aquatica Aubl.), Sontecomapan Lagoon, Veracruz. (F) Mangrove forest in El Cometa Lagoon, Tabasco. Photographs: Daniel Chávez (A), Jorge A. Meave (B, D), Derio Jiménez-López (C), Guillermo Ibarra-Manríquez (E), Jorge López-Portillo (F).
In recent decades, remote sensing has been increasingly used in the study of mangroves and other tropical wetland forests (e.g., Aschbacher et al. 1995, Proisy et al. 2000, Couteron et al. 2005, Fatoyinbo & Armstrong 2010, Solórzano et al. 2018). Most studies have been based on the analysis of vegetation reflectance to delineate their spatial distribution and temporal dynamics (Huete 1988, Gao 1996, Foody et al. 2001, Foody 2003, Lu et al. 2004, Kuenzer et al. 2011, Valderrama-Landeros et al. 2018), while other aspects, for example, their internal structure, have received less attention. In this paper, we provide an overview of the different goals pursued and achievements in studying tropical wetland forests through remote sensing. The review compares conceptual and methodological approaches, whose variety has resulted in a broad gamut of information that would probably not exist without the available theoretical framework and analytical tools. The initial sections provide a brief synopsis of tropical wetland forests to clearly define the object of this review and a historical overview of the development of remote sensing-based wetland forest studies.
Mangrove and other tropical wetland forests in a nutshell
Forested wetland ecosystems usually occur in intertidal zones of tropical and subtropical regions of the World (Saenger et al. 1983, Tomlinson 1986, Kathiresan & Bingham 2001, Lin & Dushoff 2004, Hogarth 2007, Sharma 2018; Figure 2A). The most common forest type characterizing these ecosystems is called mangrove or mangal (sensuMacnae 1969). Mangroves are typically dominated by a handful of tree species to which the term mangrove is also commonly applied. Mangrove forests are strongly influenced by tidal dynamics and occur in environments with variable salinity, from nearly fresh to hypersaline, with their optimal development in brackish water (Blasco et al. 2001, Agráz-Hernández et al. 2006). Due to their morphological and physiological attributes, mangrove trees can tolerate high temperatures, salinity, and anaerobiosis in frequently flooded substrates (Tomlinson 1986, Hogarth 2007, Sharma 2018). These attributes include aerial roots that anchor plants in unstable substrates, respiratory roots (pneumatophores and lenticels) to increase access to atmospheric oxygen, regulation of water potential through variation of xylem salt concentration, stomatal regulation, and atmospheric water uptake, and salt secretion through specialized foliar glands (López-Portillo et al. 2014, Coopman et al. 2021), vivipary (germination on the branches of the parental tree), and hydrochorous dispersal of their propagules (Blasco et al. 2001, Kathiresan & Bingham 2001, Zhou et al. 2016). Although many of these physiological features also apply to non-mangrove wetland forests, these tend to develop in very low to no salinity conditions; their canopies are composed of a less limited number of tree and palm species, among which members of Terminalia, Acoelorrhaphe and Pachira are frequent (Figure 1C, D, E).
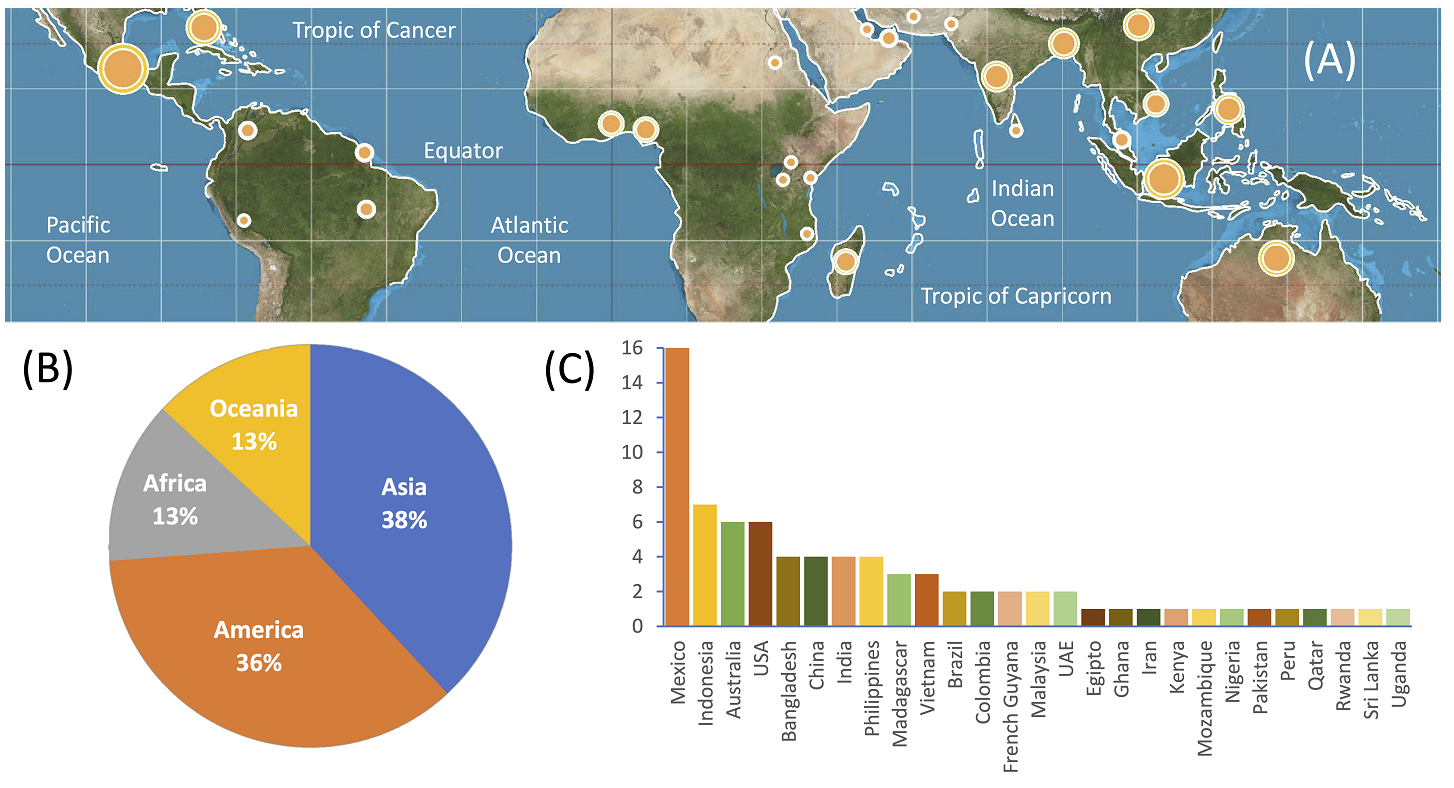
Figure 2 (A) Potential distribution of tropical wetland forests in the tropical regions of the world (30º N to 30º S). The white lines depict the coastlines of all continents where wetland forests may be found; due to scale, inland wetland forests are not shown (modified from Polidoro et al. 2010). Note the occurrence of wetland forests in arid regions of the world. The orange circles represent the locations (countries) of remote sensing-based wetland forest studies reviewed in this work; the circle size indicates the number of studies reviewed by country, according to the information shown in panel (C). (B) Continental distribution of the remote sensing-based wetland forest studies included in this review. (C) Distribution of these studies by country; USA, United States of America; UAE, United Arab Emirates. The fact of having recorded more studies from Mexico than from other countries is partly due to a higher ability of the authors to find more studies that were not easily accessible from this country. Map in (A) taken from Wikipedia (Strebe, CC BY-SA 3.0 <https://creativecommons.org/licenses/by-sa/3.0>, via Wikimedia Commons).
A notable feature of wetland forests compared with upland tropical forest types is their low species richness, particularly in the case of mangrove forests. Although species richness varies among biogeographical regions (Tomlinson 1986), the total known species richness for mangrove forests ranges from 55 to 73 taxa (Polidoro et al. 2010, Spalding et al. 2010), with Avicenniaceae and Rhizophoraceae being the most species-rich and abundant families (Hogarth 2007). Despite the relatively low tree diversity, wetland forests are recognized for their high productivity and the provision of essential ecosystem services (Whittaker & Likens 1973, Odum & Heald 1975, Tomlinson 1986, Kathiresan & Bingham 2001, Sharma 2018), including coastal protection, carbon sequestration, provision of habitat for migratory species, reproductive grounds for a specialized fauna including economically important species for fisheries, and provision of wood (Saenger et al. 1983, Lin & Dushoff 2004, Charcape-Ravelo & Moutarde 2005, FAO 2005a, Kuenzer et al. 2011).
Despite the unique character of wetland forests, their ecological fragility and high societal value, the present situation for most of them is alarming (Alongi 2008, Islam et al. 2014, Himes-Cornell et al. 2018). Factors responsible for their deterioration include both human activities (agriculture, cattle ranching, aquaculture, tourism) and natural phenomena, mainly tropical storms and climate change (Charcape-Ravelo & Moutarde 2005, Agráz-Hernández et al. 2006, Gilman et al. 2008, Kovacs et al. 2009, Cho & Qi 2023). These factors have significantly contributed to the loss of their cover worldwide (Polidoro et al. 2010), which in mangrove forests alone was between 20 and 35 % in the 1980-2005 period (Kathiresan & Bingham 2001, FAO 2005b, Hogarth 2007, Flores Mejía et al. 2010, Giri et al. 2011, Barbier et al. 2011), and have brought about a growing need to conserve and restore them. In this context, remote sensing is an option with great potential for efficiently studying mangroves and other wetland forests (Mahdavi et al. 2018).
Historical overview of remote sensing-based wetland forests studies
Early development of remote sensing took place in a military context (van der Meer et al. 2001). Nonetheless, remote sensing found its way since early times in ecosystem studies (Kumar et al. 2001). At first, aerial photographs were essential to assess and identify natural resources through forest inventories (West 1956, Colwell 1964, Aschbacher et al. 1995, Wang et al. 2019). Some inventories included mangrove cover of a region or country in thematic maps (e.g., FAO 1963, Flores Mata et al. 1971, Vázquez-Yanes 1971, Blasco et al. 1998, Sulong et al. 2002, FAO 2005b, Valderrama et al. 2014). MacDonald et al. (1971) published one of the earliest examples of the use of radar imagery to study coastal landscapes.
With the launching of the first artificial satellites to explore the territory in the 1970s, remote sensing studies underwent an explosive diversification, and their coverage expanded to the entire planet (Wulder et al. 2019). Interestingly, the first formal publications in remote sensing were seemingly related to the study of wetlands, some of which included mangrove forests (Eitel 1974, Butera 1983, Hardisky et al. 1986, Guo et al. 2017). To our knowledge, Biña et al. (1978) and Lorenzo et al. (1979) published the first analyses of spatial changes in mangroves and the monitoring of their deterioration in Southeast Asia based on Landsat images. The use of satellite imagery grew considerably during the following decade; the diversification of platforms (Landsat, SPOT, ERS-1, RADAR) increased the options to study and demarcate wetland forests, particularly mangrove communities (Herz & Jaskow 1985, Dutrieux et al. 1990, FAO 2005b, Lucas et al. 2007, Kovacs et al. 2008, Abdel-Hamid et al. 2018, Mahdavi et al. 2018). Over time, the remote sensing approach was applied to the study of mangroves in many tropical regions of the world (e.g.,Pasqualini et al. 1999, Saito et al. 2003, Kovacs et al. 2005, Satyanarayana et al. 2011: Figure 2B, C), with theoretical and technological tools providing higher-quality information on the status of vegetation at each locality (Zhang et al. 2023). More recently, community structural attributes of wetland forests (e.g., aboveground biomass, canopy height) have been explored using mathematical modeling (Couteron et al. 2005, Kayitakire et al. 2006, Proisy et al. 2007), and LiDAR (Light Detection and Ranging; Owers et al. 2018, Pereira et al. 2018, Koma et al. 2021), although wetland forest mapping and monitoring continue to prevail (e.g., Sulong et al. 2002, Saito et al. 2003, Adam et al. 2010, Barbier et al. 2011, Alsaaideh et al. 2013, Jones et al. 2016, Chatziantoniou et al. 2017, Guo et al. 2017, Yevugah et al. 2017, Roy et al. 2019, Li et al. 2023, Steenvoorden et al. 2023; Figure 3A).

Spatial delimitation and mapping of mangroves and other wetland forests
A fundamental aim in the study of mangroves and other wetland forests through remote sensing has been the definition of their spatial limits (Dale et al. 1996, Hirano et al. 2003, Yang et al. 2009, Margono et al. 2014, Wang et al. 2019, Steinbach et al. 2023), which is the basis for their cartographic expression (Figure 3A). This is a necessary step to understand the persistence of wetland forests or the changes in their spatial coverage over time (Kovacs et al. 2001, Guerra Martínez & Ochoa Gaona 2006, Ren et al. 2011, Mansaray et al. 2016, Maryantika & Lin 2017, Roy et al. 2019, Pan et al. 2020, Zhang et al. 2023). Aerial photography, Landsat, and SPOT imagery are among the most widely used inputs in the study of wetland forests due to their availability and continuity (Figure 3B; Tables 1, 2). Most studies using satellite images are based on the analysis of vegetation reflectance (Fei et al. 2011, Kuenzer et al. 2011, Abdel-Hamid et al. 2018, Apostolopoulos et al. 2023) under the tenet of an existing relationship between reflectance values recorded in the image and terrain features (Colwell 1974). However, the rapid development and growth of other remote sensing technologies, in particular LiDAR, have unlocked new ways to explore how the various elements of ecosystem structure impact functional diversity and ecosystem services (Mascaro et al. 2011, Asner et al. 2012, Davies & Asner 2014, Owers et al. 2018). LiDAR is rapidly transitioning from a demonstrative concept to a pivotal tool for estimating carbon stocks in tropical forests (Asner et al. 2012) and is thus effective at generating accurate above-ground biomass maps for mangroves, enabling carbon stock assessment and facilitating coastal management (Pereira et al. 2018).
Table 1 Classification of remote sensing-based wetland forests studies according to their main goals, resources, and platforms. Explanation of goals: Mapping, spatial delimitation of wetland forests on cartography; Classification, recognition of the different wetland types, including wetland forests, differing in structure, species composition, or health condition; Quantification, assessment of quantitative characteristics of wetland forests, particularly structural attributes; Monitoring, multi-temporal assessment of wetland forests properties to detect trends of change; Prediction, construction of models relating characteristics of the remote sensing resources and vegetation properties to predict forest attributes in sites lacking field data. See Table 2 for explanations of remote sensing input acronyms.
| Remote sensing inputs | Mapping | Classification | Quantification | Monitoring | Prediction |
|---|---|---|---|---|---|
| Passive sensors | |||||
| Aerial photography | 8, 20, 63, 85 | 7 | 12, 25, 51, 55, 85 | ||
| Satellite imagery | |||||
| Landsat | 10, 15, 18, 19, 23, 48, 54, 58, 61, 70, 77. | 11, 47, 73, 86 | 2, 35, 83 | 3, 13, 27, 29, 31, 43, 44, 53, 55, 60, 64, 68, 80, 81, 87, 91, 92, 93, 94 | 82 |
| Sentinel-2 | 67, 70, 79, 84, 88, 89, 90 | 72 | 92 | 82 | |
| SPOT | 5, 8, 9, 10, 14, 24, 70 | 47 | 4, 6, 12 | ||
| IKONOS | 26, 28 | 34, 46 | |||
| QuickBird | 26, 41 | 38 | 80 | ||
| Google Earth | 58, 66 | ||||
| RapidEye | 72 | ||||
| Hyperion | 57 | 76 | |||
| Planetscope | 72 | ||||
| WorldView | 65, 70 | 51 | |||
| Kompsat | 75 | ||||
| ICESAT | 37 | ||||
| Radiometer | 24 | ||||
| Spectrometer | |||||
| CASI | 10, 21, 45 | ||||
| AVIRIS | 22 | ||||
| ASD FieldSpec | 52 | ||||
| Field Spec Pro | 39 | ||||
| Active sensors | |||||
| Radar | |||||
| Sentinel-1 | 84, 88, 90 | 86 | |||
| Polarimetric Radar | 1, 5, 16, 17, 71 | 11, 47 | 69 | ||
| AIRSAR | 21 | ||||
| ENVISAT ASAR | 36 | ||||
| SRTM | 30, 37 | 35 | 91 | ||
| ERS-1 SAR | 16 | 69 | |||
| PALSAR | 54, 58 | 50 | 33 | 69 | |
| LIDAR | 42, 56, 59, 63, 78 | 49 | 62, 87 | ||
| GLAS | 37, 78 | ||||
1 MacDonald et al. (1971), 2Bina et al. (1978), 3Lorenzo et al. (1979), 4Dutrieux et al. (1990), 5Aschbacher et al. (1995), 6Ramsey III & Jensen (1996), 7Dale et al. (1996); 8Blasco et al. (1998), 9Gao (1998), 10Green et al. (1998b), 11Ramsey III et al. (1998), 12Rasolofoharinoro et al. (1998), 14Pasqualini et al. (1999), 15Koutsias et al. (2000), 16Kushwaha et al. (2000), 17Proisy et al. (2000), 18Kovacs et al. (2001), 19Foody et al. (2001), 20Sulong et al. (2002), 21Held et al. (2003), 22Hirano et al. (2003), 23Hossain et al. (2003), 24Saito et al. (2003), 25Dahdouh-Guebas et al. (2004), 26Wang et al. (2004); 27Hernández Cornejo et al. (2005), 28Kovacs et al. (2005); 29Berlanga-Robles & Ruiz-Luna (2006), 30Simard et al. (2006), 31Berlanga-Robles & Ruiz-Luna (2007), 32Jensen et al. (2007), 33Lucas et al. (2007), 34Proisy et al. (2007), 35Fatoyinbo et al. (2008), 36Kovacs et al. (2008), 37Simard et al. (2008), 38Myint et al. (2011), 39Wang & Sousa (2009), 40Yang et al. (2009), 41Kovacs et al. (2009), 42Fatoyinbo & Armstrong (2010), 43Berlanga-Robles & Ruiz-Luna (2011), 44Giri et al. (2011), 45Kamal & Phinn (2011), 46Satyanarayana et al. (2011), 47Fei et al. (2011), 48Alsaaideh et al. (2013); 49Wannasiri et al. (2013); 50Darmawan et al. (2014); 51Heenkenda et al. (2014); 52Zhang et al. (2014); 53Mabwoga Thukral (2014); 54Margono et al. (2014); 55Valderrama et al. (2014), 56David & Ballardo (2015), 57Demuro & Chisholm (2015), 58Aslan et al. (2016), 59David & Ballardo (2016), 60Jia et al. (2016), 61Jones et al. (2016), 62Pada et al. (2016), 63David & Ballardo (2016), 64Mansaray et al. (2016), 65Shahzad et al. (2017), 66Yevugah et al. (2017), 67Chatziantoniou et al. (2017), 68Maryantika & Lin (2017), 69Thomas et al. (2017), 70Valderrama-Landeros et al. (2018), 71Abdel-Hamid et al. (2018), 72Baloloy et al. (2018), 73Gupta et al. (2018), 74Islam & Ma (2018), 75Solórzano et al. (2018), 76Pandey et al. (2019), 77Roy et al. (2019), 78Hu et al. (2020), 79Mahdianpari et al. (2020), 80Pan et al. (2020), 81Mao et al. (2021), 82Nguyen & Nguyen (2021), 83Nguyen et al. (2021), 84De Luca et al. (2022), 85Apostolopoulos et al. 2023, 86Cho & Qi (2023), 87Flores-de-Santiago et al. (2023), 88Hemati et al. (2023), 89Li et al. (2023), 90Pham et al. (2023), 91Shafi et al. (2023), 92Steinbach et al. (2023), 93Waleed et al. (2023), 94Zhang et al. (2023).
Table 2 Explanation of acronyms of remote sensing inputs listed in Table 3.
| Acronym | Explanation |
|---|---|
| SPOT | Satellite Pour l'Observation de la Terre |
| ICESAT | Ice, Cloud and Land Elevation Satellite |
| CASI | Compact Airborne Spectrographic Imager |
| AVIRIS | Airborne visible/infrared imaging spectrometer |
| AIRSAR | Airborne Synthetic Aperture Radar |
| ENVISTAT ASAR | ENVISAT Advanced Synthetic Aperture Radar |
| SRTM | Shuttle Radar Topography Mission |
| ERS-1 SAR | European Remote-Sensing Satellite-1 Synthetic Aperture Radar |
| PALSAR | Phased Array type L-band Synthetic Aperture Radar |
| LIDAR | Light Detection and Ranging |
| GLAS | Geoscience Laser Altimeter System |
Efforts to map mangroves and other wetland forests have faced methodological challenges due to their highly variable spectral characteristics and similar reflectance spectra with the underlying soil (Adam et al. 2010, Shahzad et al. 2017). Early studies pursuing this goal based on Landsat imagery (Biña et al. 1978, Lorenzo et al. 1979) produced results fraught with significant uncertainty in setting the spatial limits of these communities. This situation improved with the development of new sensors with higher spatial and spectral resolution (Table 3), allowing greater precision to discriminate mangroves from other types of cover (Aschbacher et al. 1995, Gao 1998, Blasco et al. 1998, Pasqualini et al. 1999). At the turn of the century, the availability of very high-resolution imagery (i.e., pixels < 4 m; QuickBird, GeoEye-1, Worldview-2) opened the possibility to map wetland forests areas in greater detail; however, aerial photography is still being used (Sulong et al. 2002, Hirano et al. 2003, Wang et al. 2004, Guo et al. 2017), mainly because of their potential to discriminate the different species that occur in these communities (Kamal & Phinn 2011, Valderrama-Landeros et al. 2018).
Table 3 Synthesis of the temporal, spatial and radiometric resolutions of the different remote sensing platforms used for the study of tropical wetland forests. Please note that for Landsat, Sentinel and Spot, the information for all the sensors used in their different missions is condensed and the resolutions are given as ranges. NA, not applicable.
| Platform | Number of bands | Temporal resolution (days) |
Spatial resolution (m) |
Radiometric resolution (µm)* |
|---|---|---|---|---|
| Aerial photography | NA | Variable (on demand) | 1 | NA |
| Landsat | 2 to 9 | 16 to 18 | 15 to 100 | 0.475 to 12.51 |
| Sentinel-2 | 12 | 6 to 10 | 5 to 60 | 0.43 to 2.28 |
| SPOT | 4 to 5 | 26 | 1.5 to 20 | 0.45 to 1.75 |
| IKONOS | 4 | 14 | 1 to 4 | 0.45 to 0.90 |
| QuickBird | 5 | 2.8 | 0.65 to 2.4 | 0.45 to 0.90 |
| RapidEye | 5 | 1 | 5 | 0.4 to 0.85 |
| Hyperion | 220 | 1 to 2 | 10 to 30 | 0.4 to 2.5 |
| PlanetScope | 8 | 1 | 3 | 0.43 to 0.88 |
| WorldView | 9 | 1.1 | 0.5 to 2.0 | 0.45 to 0.9 |
| Kompsat-2 | 4 | 28 | 1 to 4 | 0.45 to 0.9 |
| ICESAT | NA | 91 | NA | 0.532 |
| CASI | 228 | 85 frames/sec | 1 | 1.9 |
| AVIRIS | 224 | Variable (on demand) | 20 | 0.4 to 2.5 |
| FieldSpec Pro | 2,151 | Variable (on demand) | 30 | 0.35 to 2.5 |
| Polarimetric Radar | NA | NA | 3 to 30 | 9.47 GHz |
| SAR | NA | NA | 5 | 20 to 80 MHz |
| GLAS | NA | 91 | NA | 0.532 |
* Except in the case of Polarimetric Radar and SAR, whose resolution is GHz and MHz, respectively.
The platforms for acquiring satellite imagery are classified as passive sensors because they use solar radiation to capture terrain details. Other types of sensors are known as active because they emit energy and record its reflection, rendering them independent from physical and weather conditions at image acquisition, such as illumination, cloudiness, and water bodies (Aschbacher et al. 1995, Green et al. 1998a, Chinea 2002, Kovacs et al. 2008, Kuenzer et al. 2011, Li et al. 2023). Interestingly, recognizing the advantages of contrasting sensor types led to their combined use (Held et al. 2003, Asner et al. 2008, Simard et al. 2008, Aslan et al. 2016), increasing the capacity and efficacy to detect, differentiate and estimate wetland forest structure (Ramsey III et al. 1998, Rasolofoharinoro et al. 1998, Kayitakire et al. 2006, Aslan et al. 2016, Wang et al. 2019, De Luca et al. 2022, Steenvoorden et al. 2023). Likewise, precision in mangrove and other wetland forests mapping has improved by combining hyperspectral with multispectral, high, and very high-resolution images (Apostolopoulos et al. 2023). This procedure allows the detection of the spectral signature (Shaw & Burke 2003, Navulur 2007, Pandey et al. 2019) and a more accurate location of precise wetland forest limits (Blasco et al. 1998, Hossain et al. 2003, Adam et al. 2010).
Monitoring wetland forest dynamics
The possibility of accurately delimiting the territorial extent of wetland forests is also relevant when aiming to assess their losses or gains, as well as their horizontal displacements (Dahdouh-Guebas et al. 2004, Hernández Cornejo et al. 2005, Shafi et al. 2023). In remote sensing, one of the most popular methods for these goals is identifying the presence of a wetland forest through terrain reflectance values (Ingram et al. 1981, Ramsey III & Jensen 1996, Mahdavi et al. 2018, Wang et al. 2019). Although temporal differences in reflectance may represent variations in illumination, atmospheric conditions, and soil moisture, among other factors, there is no doubt that they also reflect changes in vegetation cover (Singh 1989).
Interestingly, despite the large availability of satellite-based resources, aerial photography remains essential for monitoring wetland forest dynamics due to an extensive series of aerial photographs of coastal regions predating the satellite-dominated age of remote sensing. The combined use of aerial photographs with satellite images and Geographic Information Systems (GIS) has yielded accurate assessments of wetland areas reductions (Dahdouh-Guebas et al. 2004, Heenkenda et al. 2014, Apostolopoulos et al. 2023). However, when the period delimited for the analysis of wetland forest dynamics does not include the pre-satellite image era, the studies are based primarily on satellite imagery (Figure 3B). For example, in a multi-temporal analysis of landscape changes associated with coastal wetlands, Berlanga-Robles & Ruiz-Luna (2006, 2007, 2011) successfully assessed losses in wetland cover over a 20-yr period based on the use of the panchromatic bands of Landsat imagery. Using a similar approach, Giri et al. (2011) made a more precise assessment of the extent of wetlands worldwide, including both herbaceous and forested wetlands, which was used to update previous reports on these ecosystems by FAO (2005b) and correct previous overestimation. Through the amalgamation of LiDAR and other remote sensing techniques, Asner et al. (2008) not only identified and tracked invasive plant species but also evaluated their ecological impact and provided accurate geographical data for conservation and management initiatives. This latter approach holds a huge potential for identifying species in low-diversity ecosystems, such as wetland forests, where the possibility of confounding the radiometric signals from a multitude of species is much more limited than in species-rich upland tropical forests. New and more precise assessments of the extent and limits of wetland forests are needed to provide a more robust basis to evaluate coverage changes, particularly the losses, of these ecosystems worldwide (Shafi et al. 2023).
The increasingly accurate assessment of temporal changes in vegetation structure and other aspects of wetland forest communities has benefited from the use of vegetation indices, which consist of numerical relationships between different spectral bands in the images. Among the many existing indices, the most used in wetland forests studies are the Normalized Difference Vegetation Index (NDVI), the Enhanced Vegetation Index (EVI), and the Normalized Difference Water Index (NDWI) (McFeeters 1996, Gao 1998, Kovacs et al. 2005, Alsaaideh et al. 2013, Kafy et al. 2023, Shafi et al. 2023). By minimizing the effects of factors linked to biophysical terrain parameters, vegetation indices provide reliable multi-temporal information on the conservation status and the functionality of these systems (Gao 1996, Huete et al. 1997, Gao 1998, Saito et al. 2003, Wang et al. 2004, Nasiri et al. 2022, Li et al. 2023).
The launching of new technologies has enabled the use of more detailed sources of information (Pada et al. 2016, Tassi & Vizzari 2020, Pham et al. 2023). Along with the progress in image processing and classification techniques with new analytical algorithms, we have increased our capacity to assess temporal changes in wetland forests accurately (Heenkenda et al. 2014, De Luca et al. 2022, Pan et al. 2022, Shafi et al. 2023). Within this framework, one of the most promising developments for mapping and monitoring wetland forests is Google Earth Engine (GEE), a free cloud-based platform to process vast geospatial datasets that allows users to access, observe and analyze geospatial data across the entire planet (Gorelick et al. 2017, Tassi & Vizzari 2020, Kafy et al. 2023). The GEE platform contains petabytes of fully accessible images from multiple remote sensing platforms; it offers an enormous volume of Earth observation data and high-performance parallel computing equipment to analyze them (Rahaman & Shermin 2022, Hemati et al. 2023). Thus, one major advantage of operating on the GEE platform is a significant reduction in satellite image processing time, which in a short period has contributed to improving the assessment of land use and land cover change, on top of traditional mapping and vegetation monitoring efforts (Mahdianpari et al. 2020, Nasiri et al. 2022). In addition, the integration of machine learning algorithms into the GEE platform has considerably accelerated the gathering of valuable information about the state of wetland forests, and thus are handy new tools for the classification, mapping, and monitoring of these ecosystems compared to traditional methods (Tassi & Vizzari 2020, Waleed et al. 2023, Flores-de Santiago et al. 2023).
Modeling and predicting wetland forest community attributes through remote sensing
In the advancement of remote sensing, particularly concerning the study of vegetation, one of the most prominent developments refers to the use of the characteristics of digital images to model community diversity and structural attributes (e.g., basal area, biomass, crown cover, stem density, species richness, canopy height) with various degrees of precision and certitude (Strahler et al. 1986, Woodcock & Strahler 1987, Chinea 2002, Chuvieco 2020). The basic tenet of vegetation attribute modeling from images acquired through remote sensors establishes the possibility of constructing algorithms relating image spectral features (i.e., reflectance) with the physical attributes of the terrain (e.g.,Strahler et al. 1986, Woodcock & Strahler 1987, Nagendra & Rocchini 2008).
Despite substantial progress made with this approach in the last decade, modeling and predicting wetland forest attributes remain a relatively little explored field of research (Proisy et al. 2007, Giri 2016). As early attempts in this direction, Lucas et al. (2007) and Simard et al. (2006) produced examples of mangrove attribute estimations based on remote sensing data acquired through active sensors: Advanced Land Observing Satellite/L-band polarimetric SAR (ALOS-PALSAR) and Ice, Cloud, and Land Elevation Satellite/Geoscience Laser Altimeter System (ICESat/GLAS). This early work delivered the foundations for a study that, to this date, continues to stand out as the most successful effort in the realm of wetland forest attribute modeling. This is the work of Proisy et al. (2007), who predicted biomass in mangrove forests of French Guyana. Their work relied on two novel theoretical and methodological contributions in modern remote sensing: the concept of image surface metrics and the FOTO (Fourier-based Textural Ordination) method to predict mangrove biomass based on very high-resolution (1 m) IKONOS imagery (Table 3).
Surface metrics quantify pixel spatial distribution and reflectance in an image and are closely related to landscape heterogeneity (McGarigal et al. 2009). Every pixel in a scene contains reflectance data for various wavelengths with valuable information on the structural composition of the surfaces (Haralik et al. 1973, Strahler et al. 1986, Woodcock & Strahler 1987, Shaw & Burke 2003). There are two types of surface metrics: (1) first-order or tonal metrics, calculated from the analysis of the reflectance recorded in individual pixels, and (2) second-order surface metrics, also known as textural metrics, calculated with the spectral values of at least two (but usually more) contiguous pixels (Haralik 1979).
Proisy et al. (2007)’s study represents the first successful attempt to predict mangrove biomass using high-resolution satellite imagery. From their work, relevant conclusions emerged, for example, regarding the optimal pixel window size for biomass modeling. For this reason, this work is an essential reference to study mangrove forests from a perspective based on the analysis of the spatial relationships of the components both in the images and on the terrain. The findings of this study triggered much research in many temperate (Block et al. 2016) and tropical (Couteron et al. 2005, Ploton et al. 2012, Solórzano et al. 2017) plant communities, including successional forests (Gallardo-Cruz et al. 2012). However, its impact on wetland forest studies is relatively modest. A notable exception is a study conducted by Solórzano et al. (2018), also based on high-resolution imagery and in the contrast of FOTO and GLCM (Gray Level Co-occurrence Matrix), two commonly used methods to analyze image texture. Despite the clear advantages of using these methodological tools for the accurate prediction of wetland forest attributes, recent studies continue to use tonal metrics and multiple regression as the basis for the prediction of forest attributes such as aboveground biomass and carbon stocks (Baloloy et al. 2018, Hu et al. 2020, Nguyen & Nguyen 2021). Therefore, the two approaches will likely continue to co-exist for some time.
Future challenges in the study of wetland forests through remote sensing
Notwithstanding the key conceptual and methodological developments achieved in the field of remote sensing focused on the study of mangroves and other wetland forests, the best-known aspect of these communities across the tropics continues to be their spatial distribution (Table 1). Indeed, wetland forests delimitation and mapping continue to be essential tasks that will allow us to evaluate as precisely as possible its territorial extent, loss, or expansion (Green et al. 1998b, Blasco et al. 1998, Sulong et al. 2002, Wang et al. 2019, Nguyen et al. 2021). Nonetheless, it is also true that remote sensing approaches have opened many possibilities of analysis previously unforeseen, and many of them are only now beginning to be fully appreciated.
Research on wetland forests through remote sensing has faced challenges that have sometimes caused biases in the type of studies conducted, mainly favoring the mapping of these ecosystems, leaving the quantification and modeling of their structural attributes on a second plane. The main challenge is the modeling of some wetland forests characteristics, such as the complex salinity-flooding relationship (Thom 1967, Semeniuk 1980, Tomlinson 1986), that distinguish them from terrestrial ecosystems (Chatziantoniou et al. 2017), along with the difficulty to discriminate wetland forests, particularly mangroves, from other adjacent forest types using satellite imagery and other remote sensing inputs (Gupta et al. 2018). Due to the complex and highly dynamic flooding regimes that characterize wetland forests, acquiring the reflectance information of the site from remote sensing inputs is challenging. This difficulty points to the need for new methodological approaches. A promising alternative is to combine active and passive sensors as an efficient way to provide a more accurate depiction of the physical characteristics of these communities (Aschbacher et al. 1995, Green et al. 1998a, Kushwaha et al. 2000, Fatoyinbo et al. 2008, Kovacs et al. 2008, Aslan et al. 2016), even though to this date these studies continue to rely more heavily on passive than on active sensors (Figure 3B). Additional alternatives for the study of wetland forests imply working at different scales because different sensors are likely to capture different aspects of their variation (Steenvoorden et al. 2023). Finally, it would be valuable to use existing indices or propose new and more exact methods to extract specific biophysical parameters (Wannasiri et al. 2013), such as reducing the exposed water surfaces for more accurate estimates of community attributes.
While numerous LiDAR-based studies have utilized aerial vehicles for terrain data acquisition, there has been a recent emergence of the use of Terrestrial Laser Scanning (TLS), involving sensors positioned on the ground (Fröhlich & Mettenleiter 2004, Liang et al. 2016). Recent studies have showcased the effectiveness of TLS technology in precisely quantifying biomass within intricate coastal wetland vegetation. For example, Owers et al. (2018) demonstrated a good match between TLS-derived estimates of mangrove biomass and conventional allometric techniques. Furthermore, the dependability of TLS is reinforced by 3D surface reconstruction models, resulting in comparable above-ground biomass estimates for mangroves.
The concept of nature-based coastal protection is gaining increasing traction as a promising, sustainable, and cost-effective strategy to mitigate the risks of coastal flooding (van Hespen et al. 2023). Wetland forests, with their remarkable wave-attenuating capabilities, play a significant role in this natural approach to flood defense. Moreover, the restoration of mangroves and other wetland forests not only enhances flood resilience but also catalyzes a substantial economic growth (Debrot et al. 2022).
Wetlands play a pivotal role in disaster prevention, water quality improvement, and carbon storage. Given that remote sensing technology can guide restoration efforts, as well as surveillance and monitoring, especially through high-resolution platforms, such as IKONOS, Hyperion, QuickBird and PlanetScope (Table 3), we urge governments and international organizations to make greater investments in remote sensing-based research and to use it as a law enforcement tool for the protection of these threatened ecosystems.
Final remarks
The world of remote sensing is constantly growing. With the rapid development of remote sensing imagery, new approaches have emerged; undoubtedly, there are still some unresolved difficulties (Nasiri et al. 2022, Pan et al. 2022). Remote sensing-based studies of mangrove and other wetland forest communities have accomplished half a century of development. Despite such a long period of constant advancement, it is noteworthy that most studies in this field have focused on the delimitation and mapping of this ecosystem. Notably, current remote sensing studies of wetland forests also address new research questions with novel goals and aims. A limited but rapidly growing number of studies have aimed at modeling and predicting various wetland forest attributes, especially their biomass and three-dimensional structure. However, this type of research still represents a small fraction of all remote sensing-based studies focused on these ecosystems. The difficulty to accurately describe and predict community attributes in forests that possess such a complex physical structure will likely be overcome by using LiDAR and radar-based techniques (Darmawan et al. 2014, David & Ballardo 2015, 2016) and by increasing the use of machine learning and other automated information analysis processes (Mahdianpari et al. 2020, Yang et al. 2022, Pham et al. 2023).
Considering the critical conservation issues of wetland ecosystems (Kathiresan & Bingham 2001, Lee et al. 2014, Islam et al. 2014) in the face of global warming (Soares 2009, Sandilyan & Kathiresan 2012), the need to gather accurate and reliable information on these systems has become a matter of urgency. Modeling wetland forest attributes from remote sensing inputs promises to achieve this goal. However, the study of wetland forests through remote sensing is still in an early stage of development. In the future, we will undoubtedly witness conceptual and methodological progress that will enable us to surmount the challenges remaining to this date.

