











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Maize (Zea mays L.) is the most important crop cultivated in Mexico, central to the diets of urban and rural consumers. It occupies the largest planted area in the country devoted to any crop and involves cultivation by many small-scale farmers (Ureta et al. 2020).
Mexico is the primary center of origin and diversity for maize with 59 native landraces (Sánchez et al. 2000). Landraces are defined as a dynamic population(s) of a cultivated plant that has a historical origin, distinct identity, and lacks formal crop improvement, as well as often being genetically diverse, locally adapted, and associated with traditional farming systems (Camacho-Villa et al. 2005). Maize landraces in Mexico show remarkable diversity and the ability to thrive in a wide range of climates, ranging from arid to humid conditions and from temperate to tropical environments (Bellon et al. 2011). Landraces constitute an important aspect of global crop genetic resources, and their diversity is continually evolving-including in response to climate change (Pace et al. 2024).
In southeastern Mexico, the current Mayan milpa farming system has been shaped by weather variation, leading to crop diversification, an important Mayan livelihood strategy (Camacho-Villa et al. 2021). The maize varieties grown by peninsular Maya farmers exhibit characteristics of three principal maize landraces: Nal teel, Tuxpeño, and Dzit bacal which represent 75 % of the maize diversity collected in the whole Yucatan Peninsula (Fenzi et al. 2017, Ku-Pech et al 2023). Maize diversification with short-cycle and long-cycle varieties raises the possibility that farmers are deploying crop diversity to spread out temporal risk and increase harvest security (Tuxill et al. 2010). If water or heat stress occurs at the reproductive stage of maize it can disrupt the flowering process, reduce pollination, and result in poor seed formation, all of which contribute to a decrease in maize yield (El-Sappah et al. 2022, Rasheed et al. 2023).
Two contrasting landraces in terms of cycle length and plant morphology are commonly grown in the Yucatan Peninsula. The first is one of the oldest maize races grown in Mexico, Nal tel, which has a short cycle length, is short in stature, slender, with low grain yield production (Camacho-Villa & Chávez-Servia 2004). The second is the lowland tropical maize race Tuxpeño, one of the most productive and adaptable landraces that have been widely used in tropical maize improvement programs (Tuxill et al. 2010). Tuxpeño has a long cycle length, it is tall, and robust with large ears and high grain yield (Tuxill et al. 2010). However, to seize and define the potential use of maize genetic resources available in Mexico, it’s required to characterize them from a physiological perspective (Pace et al. 2024). Therefore, it is important to understand the physiological mechanisms used by different maize landraces to grow and yield in a particular agroecological region. Studies in this regard are useful for designing strategies that reduce climate's negative effects on maize yield in small-scale management systems (Pérez-Hernández et al. 2021). Vegetative traits such as crop cycle and plant height, are distinctive traits of maize landraces. Long-cycle varieties typically exhibit greater biomass production (Liu et al. 2023). In maize, plant height and leaf area positively correlate with yield due to the dependency of maize's photosynthetic capacity on leaf physiological characteristics. Enhanced leaf area enhances photosynthesis and biomass production, resulting in improved performance and higher productivity (Cong-Feng et al. 2015).
In addition, climate change affects crop productivity, as the increase in temperature and changes in rainfall distribution induce water stress in the plant due to increased evapotranspiration (Pérez-Hernández et al. 2021). This leads to dehydration and a decrease in photosynthesis, which alters the development, growth, and productivity of the plants (Rasheed et al. 2023, Mares-Guerrero et al. 2024). Physiological traits such as chlorophyll fluorescence and leaf gas exchange are frequently employed to assess the maize's response to both biotic and abiotic stresses (Singh et al. 2022). Nevertheless, few studies have explored the physiological response of maize landraces in Mexico to increase its yield potential in a region. Dos-Santos et al. (2019) explored the agronomic and physiological characteristics of 12 maize landrace populations from southern Mexico and underlined maize landraces with high photosynthesis, water use efficiency, and superior grain yield in Yucatan. Later, Mares-Guerrero et al. (2024) evaluated the physiological response of a maize landrace in monoculture and milpa systems in different soil types in Yucatan. The authors highlighted the importance of the management system as a mechanism that contributes to providing better microclimatic conditions for the physiological performance of maize (Mares-Guerrero et al. 2024).
Therefore, this work aimed to compare the growth, chlorophyll fluorescence, and gas exchange of three landrace maize populations from contrasting maize races in southeastern Mexico growing under the same field conditions.
Materials and methods
Site location and treatments. The field experiment was conducted at the facilities of Tizimin Institute of Technology, in Tizimin, Yucatan, Mexico (21° 09' 37.0" N; -88° 10' 21.7" W), between February and June of 2017. Climate conditions are warm and sub-humid with summer rains (García 2004). Data from local monthly weather conditions during the field experiment were Tm 27.0 ºC, Tmax 33.6 ºC Tmin 20.4 ºC, evapotranspiration 163 mm, and mean precipitation 77.9 mm (Figure S1) (CONAGUA 2017). The soil was categorized as Luvisol, featuring a pH of 7.7, a total nitrogen content of 0.26 %, and respective total values for phosphorus (P), potassium (K), calcium (Ca), and magnesium (Mg) at 21, 625, 5010, and 199 mg kg-1.
Three treatments, comprising three contrasting maize landrace populations collected from farmers in Yucatan, Mexico were evaluated. The landrace populations are locally named Elotillo (belong to Nal tel maize race), Xmejen-naal (belong to the complex Nal tel x Tuxpeño maize race), and Xnuc-naal (belong to Tuxpeño maize race) (Ku-Pech et al. 2023). The specific collected locations on these landraces can be found in Dos-Santos et al. 2017.
Experimental design. The experiment was set in a complete randomized block design, with six replicates assisted with a drip irrigation system. Water was supplied every third day at a flow rate of 1 L per hour for 4 hours. The sowing was carried out with a distance of 80 cm between rows and 20 cm between seeds (62,000 plants ha-1). Useful plots consisted of the center of two rows with 5 m length containing 60 plants (360 plants per treatment). The formula 120-80-00 (NPK) kg ha-1, was applied, and the fertilizer application was divided into two applications: half of the N and all the P were applied one week after the emergence of the seedlings, and the rest of the fertilizer was applied four weeks after the first fertilizer application (growth stage). Weed control was performed manually and Spinetoram was applied once for pest control.
Vegetative traits. Response variables were days to 50 % anthesis (plants disseminated pollen from the tassel (male inflorescence)) and days to 50 % silking (DS) (plants with long stigma (female inflorescence) receptive to pollination) then, the anthesis-silking interval (ASI) was calculated. A plot was considered as having reached anthesis or silking if at least 50 % of plants in a plot attained visible anthers or silking. After that, vegetative characteristics of plant height (PH), measured from the base of the plant to the tip of the tassel, and total leaf area (LA) per plant were taken in five plants at random in each experimental plot (30 plants in total per landrace). Leaf area was measured with a leaf-area meter LI-3000C (LI-COR, Lincoln, Nebraska).
Plant dry matter and biomass allocation. At physiological maturity, determined by a change in grain texture to a pasty texture and the presence of a black layer in the basal part of the grain, twenty plants per treatment were carefully extracted from the soil. After rinsing roots, plants were cut and separated by tissues: roots, stems (including leaf sheath), leaves, and tassel. The ear was separated into husk cover, grain, and cob. Samples were dried in an oven at 70 ºC to a constant weight and plant dry matter (g plant-1) was calculated.
Grain yield and yield components. The grain yield (g plant-1) and yield components were measured in the twenty plants mentioned above. Ear length and diameter, ear diameter/length ratio, number of ear rows, kernels per row, cob diameter, and weight of 100 grains were measured. The grain characteristics thickness, length, and width were measured as a mean of 10 grains taken from the central part of the ear.
Chlorophyll fluorescence and gas exchange analyses. The chlorophyll fluorescence parameters were measured in vivo with a portable pulse amplitude modulation fluorometer (PAM Walz, Effeltrich, Germany). Measurements were performed during the tassel emergence (VT stage) between 07:00 h and 09:00 h, on the second upper leaf among five leaves per treatment, with three readings taken per leaf. Previously, leaves were covered with aluminum foil and paper bags to acclimate the leaf to darkness for at least one hour before the measurements. Steady-state fluorescence, the photochemical (qP) and non-photochemical (NPQ) quenching coefficients, the relative electron transport rate (ETRPSII), and the effective quantum yield of PSII (ΦPSII) were measured according to Samaniego-Gámez et al. (2016) using a saturating light pulse of 8,000 µmol m-2 s-1 actinic light.
Fifteen measurements were taken on the central part of each leaf (five leaves per treatment, with three readings taken per leaf) using an infrared gas analyzer LI-6400XT (LI-COR, Lincoln, Nebraska) set with a photon flux density of 2,500 µmol m-2 s-1 and CO2 concentration of 400 µmol mol-1. The physiological traits measured were CO2 assimilation rate (AN), intercellular CO2 concentration (Ci), stomatal conductance (gs), transpiration (E), and water use efficiency (WUE) calculated as AN/E (Garruña-Hernández et al. 2014).
Also, the infrared gas analyzer LI-6400XT was used to measure the response of photosynthesis (CO2 assimilation rate, AN) to the CO2 concentration inside leaf air spaces (intercellular CO2 concentration, Ci) and photosynthetic photon flux density (PPFD). Between 7:00 h and 10:00 h, fully expanded apical leaves were placed in the leaf chamber and the analyses were repeated on five different leaves for each treatment. At the time, the temperature was 30 ± 2 °C and the relative humidity to 55 ± 5 %. Thus, photosynthetic CO2 response curves (AN/Ci) and photosynthetic light response curves (AN/PPFD) were calculated. For the CO2 saturation curves (AN/Ci) the concentration was from 50 to 1,500 µmol CO2 mol-1 and the PPFD was set at 1,200 µmol photons m-2 s-1. The AN/Ci curves were fitted to an empirical rectangular hyperbola (Tezara et al. 2003). For the light curves (AN/PPFD) the CO2 concentration was set at 400 µmol-1 and the PPFD was from 0 to 2,500 µmol photons m-2 s-1 (Osorio-Zambrano et al. 2023).
In addition, the maximum photosynthetic rate (Amax) and light-saturated value of photosynthesis (Asat) were estimated at a PPFD of ca. 1,200 mmol m-2 s-1 using equations developed by Von Caemmerer & Farquhar (1981). The maximum catalytic activity of Rubisco (Vcmax) and the maximum electron transport rate contributing to the regeneration of RuBP (Jmax) were calculated following Ethier & Livingston (2004). Stomatal limitation (l), which is the proportionate decrease in light-saturated net CO2 assimilation attributable to stomata, was calculated according to Farquhar & Sharkey (1982) as l = (A0 - A1) / A0, where A0 is the A at Ci of 360 mmol mol-1 and A1 is A at close about of 360 mmol mol-1. All these parameters were calculated as indicated in the manufacturer manual (LI-COR 2012).
Data analysis. The data were analyzed using one-way ANOVA, where maize landraces were treated as a factor with three levels. Data analyses were performed with Infostat software version 2020 (Di Rienzo et al. 2020). Differences among means were evaluated using the Tukey multiple comparison test with significance levels set at P ≤ 0.05. The Shapiro-Wilks test and a graphical method (Q-Q plots) were used to test the normality of the data. The biomass allocation data was adjusted for normality using an arcsine square root transformation.
Results
Vegetative traits of three maize landraces in Yucatan. Differences between maize landraces were observed for days to silking (F = 224.9, P < 0.0001), anthesis-silking interval (F = 39.4, P < 0.0001), plant height (F = 152.9, P < 0.0001), and leaf area (F =6.7, P < 0.0114). Plants from Xnuc-naal statistically had a longer cycle (89.2 days to silking) and a greater anthesis-silking interval (10.3 days) than other genotypes (Figures 1A and 1B). In addition, were statistically taller (239.1 cm) and had greater leaf area (432 cm2 per plant) than Elotillo and Xmejen-naal (Figures 1C and 1D). Many similarities were found between maize landraces Xmejen-naal and Elotillo. Both exhibited a short duration cycle, with Xmejen-naal at 67.7 days and Elotillo at 68.0 days to silking, along with minimal anthesis-silking intervals, with Xmejen-naal at 0.43 days and Elotillo at 2.17 days. Additionally, Elotillo displayed a leaf area of 376.8 cm² compared to Xmejen-naal 349.9 cm² (Figures 1D). Nevertheless, plants from Xmejen-naal were 30 cm taller than Elotillo plants (Figure 1C).
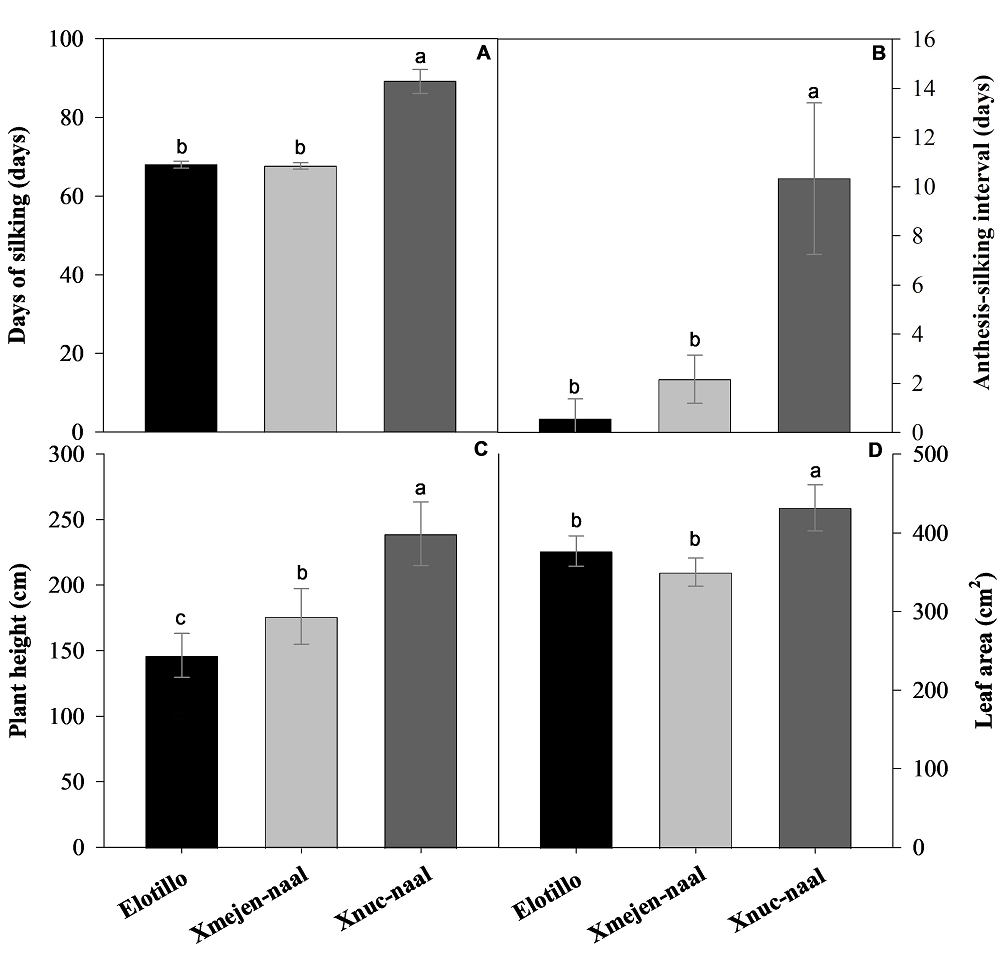
Figure 1 Days of silking A), anthesis-silking interval B), plant height C), and leaf area D) of three maize landraces in Yucatan, Mexico. Data are means ± SE. Different letters represent significant differences between treatments according to the Tukey test (P ≤ 0.05).
Plant dry matter and biomass allocation of three maize landraces. Maize plants from Xnuc-naal accumulated significantly more dry matter (273 g) than Xmejen-naal (226 g) and Elotillo (178 g) (Figure 2A).
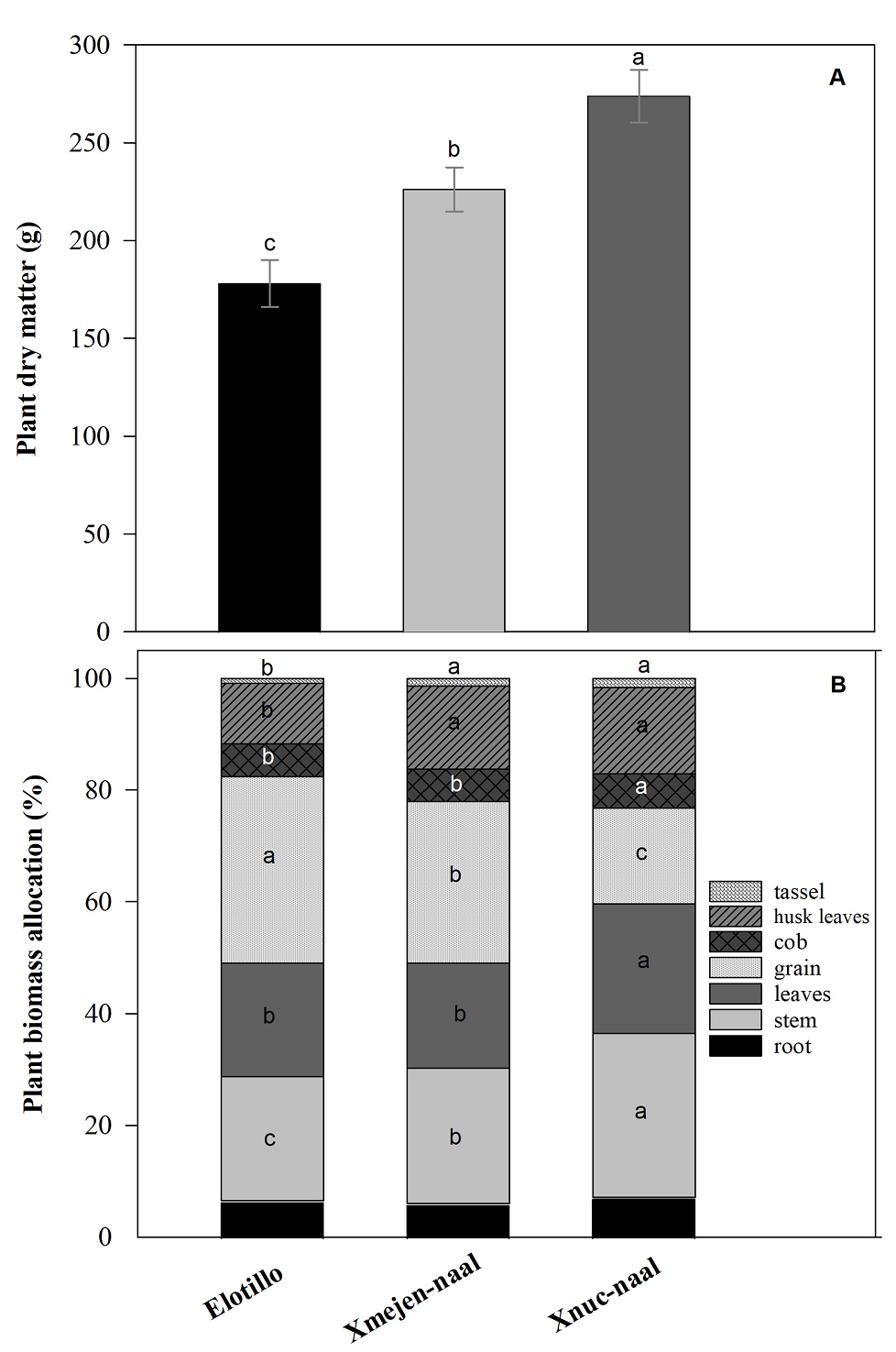
Figure 2 Plant dry matter A), and biomass allocation B) in three maize landraces in Yucatan, Mexico. Data are means ± SE. Different letters represent significant differences between treatments and within each trait, according to the Tukey test (P ≤ 0.05).
Plant biomass allocation showed differences between maize landraces in the plant components stem, leaves, grain, cob, husk cover, and tassel percentages (Figure 2B). Maize components that have accumulated more dry matter were grain, stem, and leaves with 26.5, 25.2, and 20.8 % plant biomass, respectively. The Xnuc-naal had 29.3 % dry mass on the stem, significantly greater than Xmejen-naal with 24.2 %, and Elotillo with 22.2 %. Similarly, the leaves percentage was significantly greater in Xnuc-naal with 23.2 % leaves biomass than in Xmejen-naal and Elotillo with 18.8 and 20.3 % respectively. Grain percentage was significantly higher on Elotillo with 33.4 % compared to Xmejen-naal with 28.9 % and Xnuc-naal with 17.1 % grain biomass. The cob percentage ranged between 5.8 to 6.2 % biomass. The Xnuc-naal cob percentage was significantly greater than both, the Elotillo and Xmejen-naal maize landraces. Nevertheless, Xnuc-naal and Xmejen-naal have superior husk cover with 15.5 and 14.9 %, respectively than Elotillo with 10.8 % husk cover. The tassel percentage ranged between 0.9 % on Elotillo to 1.6 % on Xnuc-naal similar to Xmejen-naal with 1.4 % tassel biomass (Figure 2B).
Grain yield and yield components of three maize landraces. Grain yield and yield components were also measured on maize landraces (Table 1). Grain yield was statistically highest on early landraces Elotillo (69.2 g plant-1) and Xmejen-naal (65.3 g plant-1) compared to Xnuc-naal with 46.5 g plant-1. Nevertheless, ear length was greater (13.7 cm) on Xnuc-naal plants than on Elotillo and Xmejen-naal (11.4 and 12.3 cm respectively). In contrast, ear diameter was larger on early landraces Xmejen-naal with 3.9 cm and Elotillo with 3.8 cm ear diameter. These variables directly affected ear diameter/length ratio equally superior on landraces Xmejen-naal and Elotillo with 0.32 and 0.34 units, respectively, that exceeded Xnuc-naal with 0.25 units ratio, as well for, the variable cob diameter statistically superior on both early landraces (Xmejen-naal and Elotillo) with 2.3 cm mean cob diameter. The variable number of ear rows showed differences between treatments (Table 1). The landrace Elotillo resulted in a greater number of ear rows (18 rows) than Xmejen-naal (13 rows) and Xnuc-naal (10 rows). Maize grain thickness was greater on Elotillo with 3.8 mm, followed by Xmejen-naal (3.7 mm) and Xnuc-naal with 3.5 mm. Nevertheless, grain length and width were higher on Xnuc-naal (9.6 mm and 8.7 mm, respectively) than both, Elotillo and Xmejen-naal. Similarly, the weight of 100 grains was higher on Xnuc-naal with 23.4 g than on Xmejen-naal and Elotillo, with 22.5 and 18.3 g, respectively. The variable grain row number was greater on Xnuc-naal with 27 grains, followed by Xmejen-naal with 24 grains and Elotillo with 22 grains (Table 1).
Table 1 Grain yield and yield components measured on three maize landraces in Yucatan, Mexico.
| Yield components | Elotillo | Xmejen-naal | Xnuc-naal |
|---|---|---|---|
| Grain yield (g plant-1) | 69.2 ± 4.1a | 65.3 ± 3.4a | 46.5 ± 3.3b |
| Ear length (cm) | 11.4 ± 0.3b | 12.3 ± 0.3b | 13.7 ± 0.4a |
| Ear diameter (cm) | 3.8 ± 0.1a | 3.9 ± 0.1a | 3.4 ± 0.1b |
| Diameter/Length | 0.34 ± 0.1a | 0.32 ± 0.1a | 0.25 ± 0.1b |
| Cob diameter (cm) | 2.3 ± 0.1a | 2.4 ± 0.0a | 1.9 ± 0.0b |
| Number of ear rows | 18.3 ± 0.4a | 13.2 ± 0.3b | 9.7 ± 0.2c |
| Grain thickness (mm) | 3.8 ± 0.1a | 3.7 ± 0.1ab | 3.5 ± 0.1b |
| Grain length (mm) | 8.8 ± 0.2b | 9.4 ± 0.1a | 9.6 ± 0.2a |
| Grain width (mm) | 6.6 ± 0.1c | 7.9 ± 0.1b | 8.7 ± 0.1a |
| Weight of 100 grains (g) | 18.3 ± 0.2c | 22.5 ± 0.1b | 23.4 ± 0.2a |
| Grain row number | 22.3 ± 0.9b | 24.0 ± 0.6ab | 27.0 ± 1.5a |
Data are means ± ES. Different letters in the same line represent significant differences between treatments according to the Tukey test (P ≤ 0.05)
Chlorophyll fluorescence in three maize landraces. Quantum yield of photosystem (ΦPSII) shows a similar trend in the three maize landraces from lowest to highest photosynthetic photon flux density and the plants of the three landraces showed high values of fluorescence (Figure 3A). The photochemical quenching (qP) showed that plants from landraces Xmejen-naal and Elotillo were superior to Xnuc-naal plants (Figure 3B). Likewise, Elotillo had the highest values of non-photochemical quenching (NPQ) concerning Xmejen-naal and Xnuc-naal plants respectively (Figure 3C). The electron transport rate (ETR) of PSII was higher on landraces Xmejen-naal and Elotillo and greater than Xnuc-naal (Figure 3D).
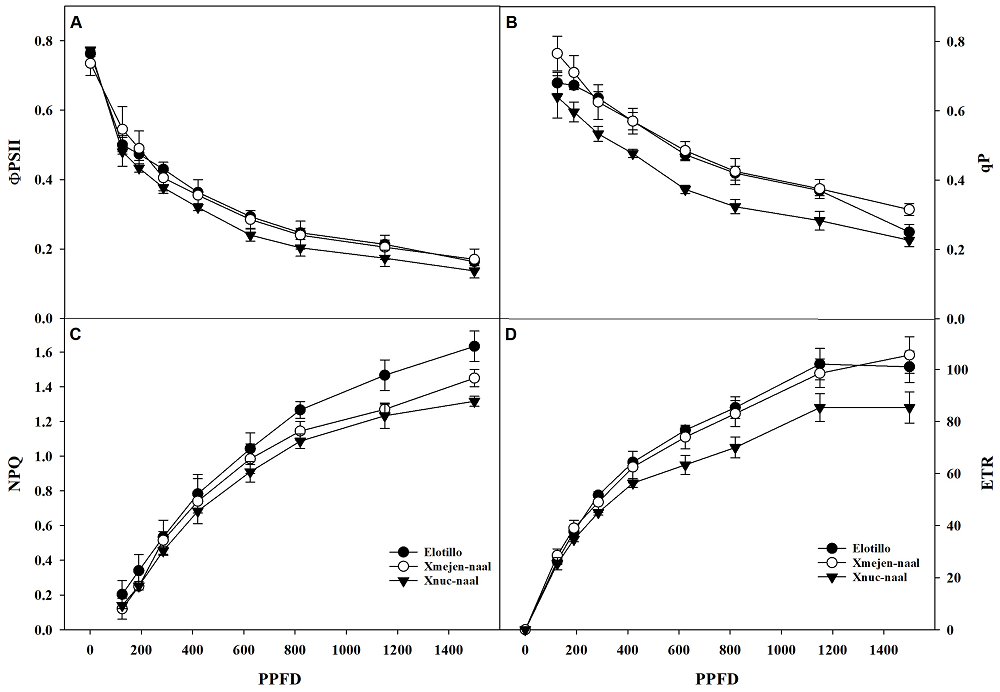
Figure 3 Quantum yield of photosystem II (ΦPSII) A), photochemical quenching (qP) B), non-photochemical quenching (NPQ) C), and electron transport rate (ETR) D) in response to the photon flux density for the photosynthesis (PPFD) of three maize landraces in Yucatan. Data are means ± SE.
Gas-exchange analyses of three maize landraces. The net CO2 assimilation rate (AN) was higher in the landrace Xnuc-naal with 39.3 µmol m-2 s-1 and statistically similar to the Elotillo (Figure 4A). The stomatal conductance (gs) and transpiration (E) were on average 13 and 28 % greater in the Xnuc-naal and Elotillo than in Xmejen-naal (Figures 4B and 4C). The water use-efficiency (WUE) was statistically superior on Xmejen-naal with 8.1 µmol CO2 mmol H2O than on Elotillo (7.2 µmol CO2 mmol H2O) and Xnuc-naal (6.4 µmol CO2 mmol H2O) (Figure 4D).
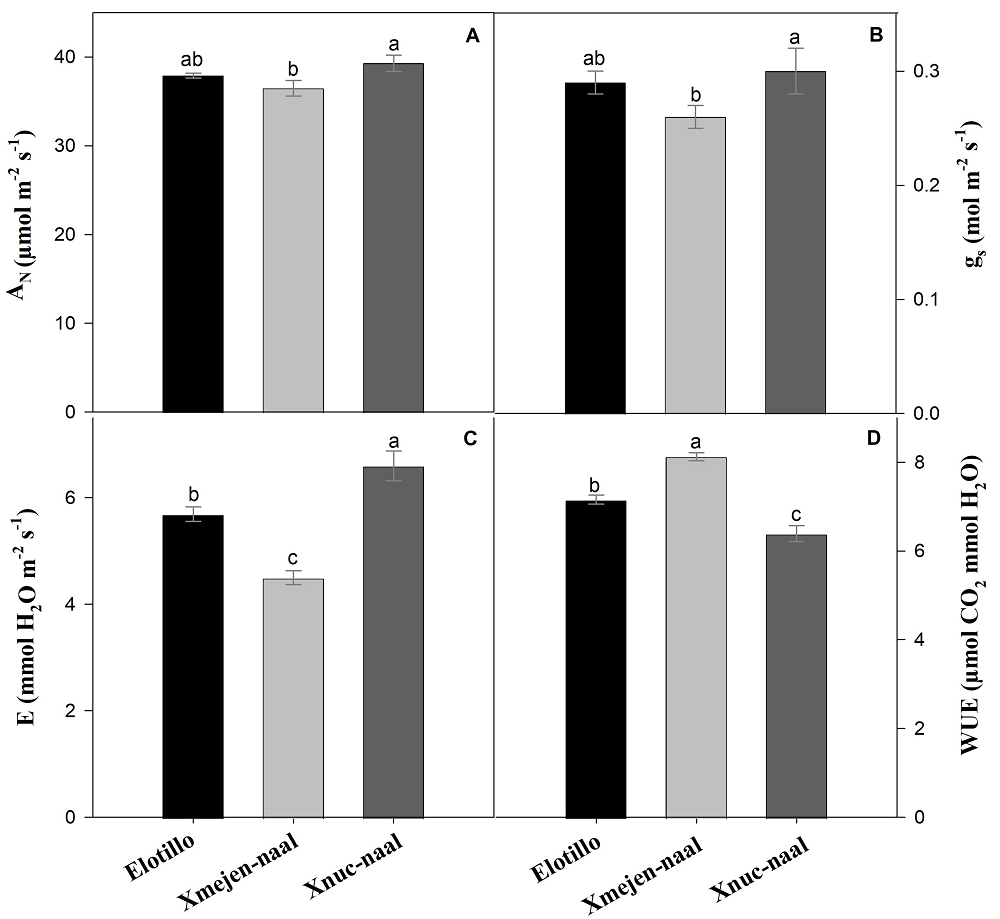
Figure 4 Net CO2 assimilation rate (AN) A), stomatal conductance (gs) B), transpiration (E) C), and water use efficiency (WUE) D) of three maize landraces in Yucatan, Mexico. Data are means ± SE. Different letters represent significant differences between treatments according to the Tukey test (P ≤ 0.05).
The AN/Ci curve forms a hyperbola rectangular for all three maize landraces and photosynthetic activity (AN) increases with intercellular CO2 concentration until a saturation point different between maize landraces (Figure 5A). In the landraces, Elotillo and Xmejen-naal, the CO 2 saturation point was 250 and 350 µmol CO2 mol-1, respectively; in the Xnuc-naal the saturation point of the photosynthetic activity was higher than others and close to 700 µmol CO-2 mol-1. Thus, the photosynthetic acclimation (when plants reach their photosynthetic saturation point) was shown in all genotypes but, both Elotillo and Xmejen-naal reached the photosynthetic acclimation before Xnuc-naal. In addition, dark respiration only was observed in the first point of Xnuc-naal. In this way, both Elotillo and Xmejen-naal reached the compensation point (30.7 and 40.6 µmol CO-2 mol-1, respectively) before Xnuc-naal (81.4 µmol CO-2 mol-1) that is, the assimilation of carbon by photosynthesis exceeded the CO2 emitted by the effect of cellular respiration (Figure 5A).
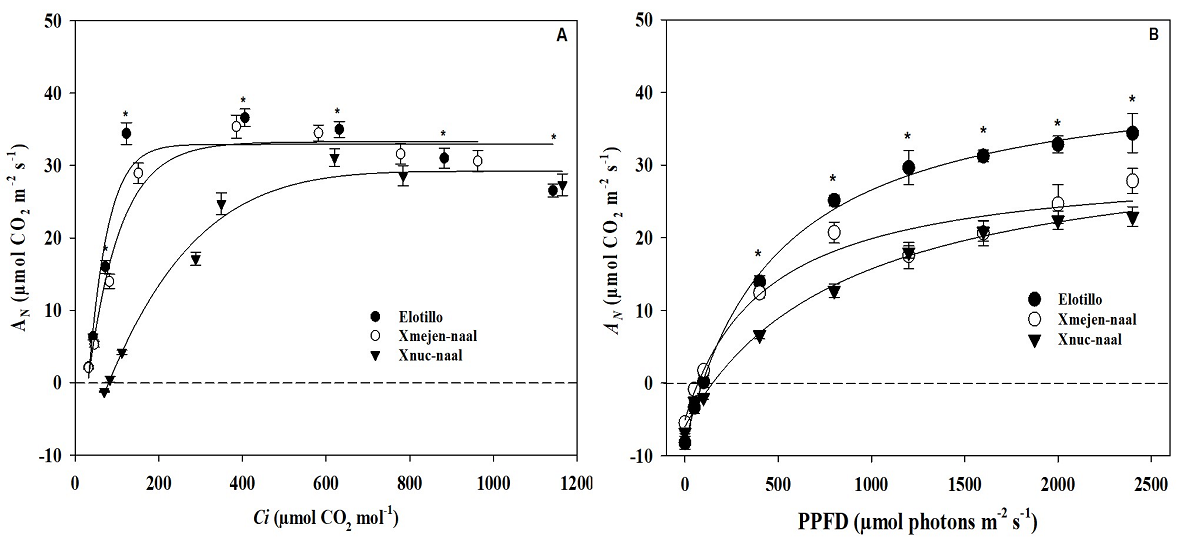
Figure 5 Photosynthetic CO2 response curves (AN/Ci) A), and photosynthetic light response curves (AN/PPFD) B) in three maize landraces in Yucatan. Data are means ± SE. * ANOVA (P ≤ 0.05).
In the AN/PPFD curve response, dark respiration was observed in all three maize landraces however, the compensation point was, 99.7, 69.5.0, and 145.8 µmol photons m-2 s-1 for Elotillo, Xmejen-naal, and Xnuc-naal, respectively (Figure 5B). In none of the maize landraces, photosynthetic acclimation was observed. However, the highest assimilation values were observed in Elotillo when PPFD increased (Figure 5B).
The CO2-saturated value of Amax ranged from 32.1 µmol m-2 s-1 on Xnuc-naal at a maximum of 36.5 µmol m-2 s-1 on Elotillo followed by Xmejen-naal with 35.9 µmol m-2 s-1. The light-saturated value of photosynthesis (Asat) was significantly higher on Elotillo with 34.5 µmol m-2 s-1 than on landraces Xmejen-naal (25.6 µmol m-2 s-1) and Xnuc-naal (24.8 µmol m-2 s-1). The apparent maximum carboxylation activity of Rubisco (Vcmax) was higher on both Xmejen-naal and Elotillo (96.3 and 95.1 µmol m-2 s-1 respectively) compared to Xnuc-naal with 77.9 µmol m-2 s-1. The maximum regeneration rate of RuBP was higher on Xmejen-naal (117.8 µmol m-2 s-1) than on Elotillo (87.9 µmol m-2 s-1) and Xnuc-naal (80.4 µmol m-2 s-1). Also, the stomatal limitation percentage (l) was higher on Xnuc-naal at 31.4 %, compared to Xmejen-naal (20.2 %) and Elotillo at 12.1 % (Table 2).
Table 2 Light-saturated value of photosynthesis (Asat), maximum photosynthetic rate (Amax), maximum catalytic activity of Rubisco (Vcmax), maximum regeneration rate of RuBP (Jmax), and stomatal limitation (l) of three maize landraces in Yucatan, Mexico.
| Elotillo | Xmejen-naal | Xnuc-naal | |
|---|---|---|---|
| Asat (µmol m-2 s-1) | 34.5 ± 2.8a | 25.6 ± 1.9b | 24.8 ± 2.0b |
| Amax (µmol m-2 s-1) | 36.5 ± 1.5a | 35.9 ± 1.8ab | 32.1 ± 2.3b |
| Vcmax (µmol m-2 s-1) | 95.1 ± 4.1a | 96.3 ± 5.8a | 77.9 ± 3.5b |
| Jmax (µmol m-2 s-1) | 87.9 ± 6.7b | 117.7 ± 8.4a | 80.4 ± 5.2b |
| l (%) | 12.1 ± 1.1c | 20.2 ± 1.8b | 31.4 ± 2.2a |
Data are means ± ES. Different letters in the same line represent significant differences between treatments according to the Tukey test (P ≤ 0.05).
Discussion
Vegetative traits of three maize landraces in Yucatan. The vegetative performance was evaluated in terms of days to silking, anthesis-silking interval, plant height, and leaf area. Maize landraces showed a differentiated response concerning cycle length and vegetative morphology. The short cycle maize landraces, Elotillo and Xmejen-naal, were characterized by short plants with lesser leaf area and accurate flower synchrony. Contrasting, the long cycle landrace Xnuc-naal has tall plants, great leaf area, and an extended anthesis-silking interval (Figure 1). Plant height and leaf area in maize are highly positively associated with yield because the photosynthetic capacity of maize largely depends on the physiological characteristics of leaves (Cong-Feng et al. 2015). In the case of Xnuc-naal, although it stood out in plant height and leaf area, the extended anthesis-silking interval could have affected its grain yield performance. Data from biomass allocation above also corroborate this hypothesis. Also, if high temperatures or water deficits occur during the reproductive stage of maize it can disrupt the flowering process, decrease fecundation rates, and lead to inadequate seed formation. Even with an irrigation system in place, elevated temperatures alone could have a substantial negative impact on maize yields (Castro-Nava et al. 2014, El-Sappah et al. 2022). Then, short-cycle maize landraces with short anthesis-silking intervals are better adapted to the current short rainy season and drought conditions in the region. Revilla et al. (2016) suggested that dwarfism may be a strategy for improving plant tolerance to drought stress conditions resulting in reduced water requirements and increased photosynthetic activity. Additionally, note that maize landraces Elotillo and Xmejen-naal had low plant height (below 200 cm), which gives them tolerance to stem lodging.
Plant dry matter and biomass allocation of three maize landraces. The landrace Xnuc-naal demonstrated high dry matter production and exhibited a low harvest index, accumulating high dry mass proportion on stem, leaves, and husk cover. Our results suggested that the landrace Xnuc-naal has a great aptitude for forage maize production. Maize landraces usually are tall plants that encourage higher production of dry matter when compared to hybrids (Robles-Jimenez et al. 2021). Similar findings were reported by Robles-Jimenez et al. (2021), who assessed the forage yield of maize landraces in Mexico. The authors reported an average yield of 279.1 g plant-1 in maize landraces, which is close to the observed yield of 273.8 g plant-1 in the maize landrace Xnuc-naal in Yucatan. In a previous study conducted by the authors, we assessed the growth and yield of tropical maize landraces and commercial cultivars in Yucatan (Dos-Santos et al. 2024). We observed that maize dry matter ranged from 141 g plant-1 in the landrace Elotillo to 253 g plant-1 in an Xnuc-naal landrace population, which is lower than the dry matter weight observed in this study, which could potentially be attributed to differences in soil fertility. It has been reported that plant dry matter varies according to the cultivar, the fertility of the soil, the cutting age, and the plant density, among other factors (Li et al. 2015).
Also, plant biomass allocation showed differences in the percentage composition of the parts that make up the maize plant with stem, leaves, and ear maize components that accumulated more dry matter (Figure 2B). In addition, results showed differences between treatments in most plant components. These differences especially stand out on stem percentage biomass that was superior on Xnuc-naal, grain percentage, especially higher on Elotillo, and husk cover percentage greater on Xnuc-naal and Xmejen-naal (Figure 2B). In the study of Amador & Boschini (2000) with a maize forage landrace, the authors showed that stem had accumulated the highest percentage of plant dry matter with 56.6 %, and leaves and ear, 14.9 and 20.0 % DM, respectively. Additionally, in the aforementioned study conducted by the authors in Yucatan, it was observed that a greater proportion of plant components consisted of stems, accounting for 35.5 %, followed by grain at 22.4 % and leaves at 16.8 % (Dos-Santos et al. 2024).
Grain yield and yield components of three maize landraces. All agronomic traits evaluated showed statistical differences between maize landraces. Several similarities were identified for grain yield and ear traits between Elotillo and Xmejen-naal differently from Xnuc-naal. However, statistical differences between all three maize landraces were observed in the number of ear rows and grain traits. The grain yield was 31 % higher on the short cycle landraces Elotillo and Xmejen-naal than on the long cycle landrace Xnuc-naal. Maize landraces Elotillo and Xmejen-naal also stood out on yield traits such as ear diameter, cob diameter, number of ear rows, and grain thickness. These maize landraces are characterized by short ears and grains. Similar results on grain yield and yield components were observed by Dos-Santos et al. (2019) and Villalobos-González et al. (2019) with maize landraces from southeastern Mexico. In contrast, Xnuc-naal stood out on traits such as ear length, grain length, grain width, weight of 100 grains, and grain row number. It is characterized by large ears and grains nevertheless, in our study grain yield was inferior to Elotillo and Xmejen-naal. Dos-Santos et al. (2019) evaluated the agronomic performance of 12 maize landraces populations from southeastern Mexico and reported a grain yield between 45 to 118 g plant-1. Similarly, Villalobos-González et al. (2019) evaluated 13 maize landrace populations from the Yucatan peninsula and reported a grain yield between 37 to 118 g plant-1. Based on those studies, the maize landraces Elotillo and Xmejen-naal are positioned in the intermediate range of grain yield within maize landraces from the Yucatan peninsula. In addition, in the study of Villalobos-González et al. (2019) the landrace populations Xmejen-naal and Xnuc-naal were statistically similar for grain yield with 96.1 and 99.1 g plant-1.
Chlorophyll fluorescence in three maize landraces. The quantum yield of the photosystem (ΦPSII) measures the proportion of light absorbed by chlorophyll associated with PSII that is used in photochemistry (Cong-feng et al. 2015). In our study, the three landraces evaluated showed a high photosynthetic capacity in vivo which denotes its adaptability to the conditions of southeastern Mexico. It has been shown that the activity PSII potential decreases when the plants are subjected to abiotic stress (Rasheed et al. 2023). Nevertheless, the electron transport rate (ETR) of PSII was higher in the leaves of Xmejen-naal and Elotillo. These maize landraces exhibit an enhanced ability to tolerate the stress induced by excessive light energy, suggesting the potential for increased photosynthesis rates and enhanced plant growth.
The photochemical quenching (qP) denotes the proportion of excitons captured by open traps and being converted to chemical energy in the PSII reaction center and NPQ that dissipates excess excitation energy as heat (Samaniego-Gámez et al. 2016). Therefore, plants from landraces Xmejen-naal and Elotillo were more efficient at operating the light absorbed, than Xnuc-naal plants. Our data implicate that Elotillo and Xmejen-naal landraces allocated more energy to photochemical processes (qP) and lower to NPQ. In this manner, Elotillo and Xmejen-naal provide more electrons (ETR) for photosynthetic carbon metabolism, and subsequently higher photosynthesis rate than Xnuc-naal. Cong-Feng et al. (2015) also analyzed eco-physiological photosynthetic traits in maize lines released in recent decades. The authors indicate that the increase in grain yield in modern lines is primarily due to a higher photosynthetic capacity of the maize lines. Thus, eco-physiological traits such as increased light energy capture capacity and the performance indices of the photochemical process could be efficient secondary improvement characteristics, as they appear to be genetically correlated with grain yield.
Gas-exchange analyses of three maize landraces. Gas-exchange analyses support in part data from leaf photochemical. The three maize landraces evaluated showed a higher photosynthetic capacity (AN) nevertheless, the stomatal conductance (gs) and transpiration (E) were greater in the Xnuc-naal and Elotillo than in Xmejen-naal. The gs is an indicator of the degree of openness of the stomata, while E indicates the amounts of molecules of water lost per CO2 molecules entering in leaves (Samaniego-Gámez et al. 2016). Our results on WUE showed that the maize landrace Xmejen-naal maintains a photosynthetic mechanism efficiently and reduces water release by leaves. In a study conducted by Dos-Santos et al. (2019) on maize landraces from the Yucatan Peninsula, the authors reported WUE values ranging from 6.4 to 8.9 µmol CO2 mmol H2O and AN value within the range of 34.5 to 39.9 µmol m-2 s-1. Oliveira et al. (2020) compared the physiological response of maize landraces and one maize hybrid at different phenological stages, in Brazil. Although the hybrid showed better physiological characteristics than maize landraces. The results from both studies closely correspond with the observations noted in the current study regarding AN and WUE. Conversely, Revilla et al. (2016) evaluated the physiological characteristics of maize populations from the Sahara and Spain. Authors reported a net photosynthetic rate between 20.3 to 27.6 µmol m-2 s-1 lesser values than on subtropical maize landraces from Yucatan. In addition, Janzen et al. (2022) observed that lowland maize landrace populations, such as Mayan maize landraces, show consistently high δ13C, indicating low WUE compared with the Mexican highland populations, detailing drought-adapted maize landraces in the Mexican highlands. Our findings regarding WUE indicate that certain maize landraces in southern Mexico can efficiently maintain photosynthetic mechanisms while releasing minimal water through their leaves and have promising characteristics for future selection studies.
The AN/Ci curve indicated that maize landraces Elotillo and Xmejen-naal maintain a photosynthetic mechanism more efficiently with a high AN rate and low Ci concentration. Also, maize landraces Elotillo and Xmejen-naal showed superior photosynthetic performance in the Amax as well as in the light-saturated value of photosynthesis (Asat). The high photosynthetic efficiency observed in these landraces (Elotillo and Xmejen-naal) might have contributed to the enhanced grain yield and yield components observed in these landraces, as indicated in Table 1. Castro-Nava et al. (2014) evaluated two contrasting maize landraces affected by high temperatures and drought in the north of Mexico. In that study, the authors showed that photosynthesis was higher in the stress-tolerant genotype than in the stress-susceptible genotype. The variations observed between tolerant and susceptible plants were primarily attributed to greater leaf area and enhanced photosynthetic capacity.
Also, both Vcmax (the apparent maximum carboxylation activity of Rubisco) and Jmax (the maximum regeneration rate of RuBP) are critical parameters used to understand the limitations and potential of photosynthesis in plants (Li et al. 2015). In this study, Vcmax and Jmax were also superior in Xmejen-naal and Elotillo than that on Xnuc-naal. According to Meneses-Lazo et al. (2018), an increase in electron transport efficiency enhances the production of ATP and NADPH, leading to an efficient regeneration of RuBP. The process of photosynthesis relies on Rubisco's ability to utilize RuBP, along with the Calvin cycle's capacity and the electron transport system, to effectively regenerate RuBP. The electron transport rate in Figure 3D above supports this asseveration. In contrast, Xnuc-naal exhibited a higher stomatal limitation percentage (l), suggesting that stomatal closure played a significant role as a limiting factor for its photosynthetic potential performance.
In conclusion, maize landraces evaluated were defined by cycle length, plant morphology, and yield performance. The short-cycle maize landraces Elotillo and Xmejen-naal exhibit superior physiological traits, characterized by an efficient photosynthetic mechanism, featuring high rates of assimilation (AN) and low intercellular CO2 concentration (Ci). These traits enable them to optimize water utilization effectively, leading to higher grain yields compared to the Xnuc-naal. These landraces also can contribute to the development of improved maize varieties with enhanced physiology efficiency in southeastern Mexico. With great dry matter production and efficient allocation across the stem, leaves, and husk cover, the landrace Xnuc-naal has a great aptitude for forage production.
Supplementary material
Supplemental material for this article can be accessed here https://doi.org/10.17129/botsci.3459

