











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
Los Atherinopsidae son teleósteos marinos pelágico costeros y dulceacuícolas, de regiones templadas a tropicales del Nuevo Mundo, que se diferencian de los Atherinidae por tener el premaxilar retráctil, sin proceso maxilar y con el extremo posterior expandido, y por tener conectado el canal sensorial preopercular del sistema sensorial de la línea lateral con el canal mandibular. La familia Atherinopsidae está formada por 11 géneros (Nelson, 2006), entre los cuales se encuentra Chirostoma. C. humboldtianum (Valenciennes, 1835) es la primera especie íctica descrita para la ciencia en México, endémica del Altiplano Mexicano, habita de forma discontinua, en sistemas lénticos de la cuenca del Sistema Lerma- Santiago, desde el Estado de México hasta Nayarit, pasando por Michoacán y Jalisco Hernández-Rubio et al. (2006). Actualmente ha sido extirpado de la Cuenca de México. Esta especie, junto con las demás de su género, ha sido de gran trascendencia ecológica, económica y cultural, desde tiempos prehispánicos.
El desarrollo de estudios anatómicos es un aspecto fundamental en su biología y su cultivo, debido a que los procesos energéticos y las funciones básicas de los peces tienen lugar a expensas de la energía que entra al organismo en forma de alimento (Nikolsky, 1963). Se han desarrollado estudios anatómicos en diversas especies de aterinópsidos, Paulo-Maya y Cortés (1996) describieron la osteología de Atherinella balsana (Meek, 1902); otros autores han descrito aspectos parciales del esqueleto visceral y la dentición, como Rodríguez-Ruiz y Granado-Lorencio (1988), Lameiro-Rodrigues y Bembenuti (2001), da Silva et al. (2003), Soria-Barreto y González-Díaz (2004), Soria-Barreto y Paulo-Maya (2005) y Keivany (2014).
Entre el limitado número de trabajos sobre el tubo digestivo de aterinópsidos se encuentra el de Logothetis et al. (2001) sobre Atherinops affinis (Ayres, 1860) y el de Horn et al. (2006) sobre especies de los géneros Atherinops y Leuresthes. Respecto a la musculatura mandibular de Atherinidae, sólo se encuentra el trabajo de Werneburg (2015), quien ilustra parte de la musculatura mandibular, palatina y opercular de Atherina boyeri Risso, 1810. Los trabajos relacionados con el sistema digestivo del género Chirostoma se reducen a los de Martínez-Palacios et al. (2006) y Ross et al. (2006); sin embargo, carecen de la descripción de la totalidad del esqueleto visceral y de su musculatura. No obstante, el trabajo reciente de Kobelkowsky (2012) describe en Chirostoma estor estor Jordan, 1879 tanto el esqueleto visceral, como la musculatura visceral y el tubo digestivo.
De acuerdo con Wilson y Castro (2011) el tubo digestivo de los peces sigue el mismo patrón morfológico de los vertebrados y su estudio permite entender la nutrición de estos vertebrados en su ambiente natural y en sistemas de cultivo. Varios autores como Moyle y Cech (2000) indican que el sistema digestivo de los peces teleósteos está formado por un tubo o tracto digestivo y unas glándulas anexas al mismo. El tubo digestivo se inicia con la boca y termina con el ano, y está regionalizado en cavidad bucofaríngea, esófago, estómago e intestino. Las glándulas anexas al tubo digestivo son el páncreas y el hígado.
Tomando en cuenta que la boca, la cavidad bucal y la cavidad faríngea están estructuradas por huesos, los cuales pueden llevar dientes y que se mueven por acción de músculos, se manifiesta la necesidad de conocer su forma, sus relaciones articulares y por lo tanto, de entender su papel en el proceso de captación del alimento y su transporte al esófago. Particularmente para C. humboldtianum, es vital el conocimiento derivado de la organización del sistema digestivo para describir los mecanismos de alimentación, que permitan implementar su cultivo y conservación. El objetivo del presente estudio es la descripción de su sistema digestivo en la etapa adulta, incluyendo el esqueleto y la musculatura viscerales.
Materiales y métodos
Se dispuso de siete ejemplares adultos de C. humboldtianum de 140 a 168 mm de longitud patrón, fijados en formaldehído al 10% y conservados en alcohol etílico al 70% provenientes de la presa Tiacaque, Jocotitán, Edo de México, México (19º 38´ 29 ´´ N y 99 º 42´ 27´´ O). Dichos ejemplares se depositaron en la Colección Nacional de Peces Dulceacuícolas Mexicanos de la ENCB-IPN, con el número de catálogo ENCB-IPN-P6478.
Se aplicó la siguiente técnica de disección del sistema digestivo:
Se retiró la piel de la región cefálica con el objetivo de exponer los músculos viscerales más superficiales, los cuales se analizaron y describieron.
Se removieron tanto los músculos viscerales como el globo ocular con los correspondientes músculos extra oculares, con el propósito de disponer de mayor espacio para la observación de los músculos palatinos y de la suspensión mandibular.
Se analizaron y describieron las mandíbulas superior e inferior, la serie palatina, la suspensión mandibular y la serie opercular.
Se retiraron las series de huesos antes mencionadas con el fin de exponer el aparato hioideo, las branquias y parte de la cavidad bucofaríngea.
Se retiraron el músculo protractor hyoidei y el aparato hioideo.
Se desprendieron los filamentos branquiales para evidenciar los arcos branquiales y los músculos que los relacionan dorsalmente con el neurocráneo (levatores externos, levatores internos y levator posterior), los que los relacionan con el urohial (rectus communis) y que lo hacen con el cleitrum (pharyngo clavicularis externus y pharyngoclavicularis internus).
Se retiraron los músculos antes mencionados, exponiendo el urohial, los arcos branquiales y el músculo retractor dorsalis.
Se analizaron y describieron los músculos anteriormente mencionados.
Se separó por completo el aparato branquial para su análisis anatómico.
Se expuso la cavidad visceral retirando la piel de un costado del cuerpo, y a continuación los miotomos correspondientes, con el objeto de evidenciar y desarticular las costillas.
Se analizó y describió la organografía de la cavidad visceral y se determinaron el enrollamiento del intestino y su longitud. Se relacionó cualitativamente la longitud del intestino con la longitud patrón del pez.
Se extrajeron el tubo digestivo y el hígado, seccionando transversalmente el esófago, el recto, las venas hepáticas y removiendo el mesenterio dorsal.
Se realizó un corte longitudinal del tubo digestivo observando las características de su superficie interna.
Se describió la forma del hígado y de la vesícula biliar, y se determinó el punto de conexión del colédoco o conducto biliar común con el intestino.
Las disecciones de los ejemplares se realizaron principalmente bajo un microscopio estereoscópico Olympus, con el propósito de observar, describir e ilustrar el tubo digestivo, el hígado, el esqueleto visceral y la musculatura visceral.
Las descripciones se realizaron organizando el sistema digestivo de la especie en estudio en los siguientes componentes: tracto digestivo e hígado, esqueleto visceral y musculatura visceral.
Se realizó una comparación de la morfología del sistema digestivo de C. humboldtianum con la de Chirostoma estor estor (Kobelkowsky, 2012) debido a sus semejanzas en los principales caracteres osteológicos, miológicos y del tubo digestivo e hígado. Para tal efecto se utilizaron siete ejemplares adultos de cada especie.
Las ilustraciones se realizaron mediante una camera lucida o tubo de dibujo acoplada a un microscopio estereoscópico Wild M3Z.
La terminología osteológica siguió los criterios de Gregory (1959) y Dyer (1997); la terminología de la musculatura se realizó de acuerdo al criterio de Winterbottom (1974); y la correspondiente a los ligamentos acorde a Dyer (1997).
Resultados
Tracto digestivo. El tracto digestivo de C. humboldtianum está formado por la boca, la cavidad bucofaríngea y el tubo digestivo, que a su vez está constituido por el esófago, el intestino y el ano. La especie en estudio es agástrica, por carecer de estómago diferenciado. Anexo al intestino se localiza el hígado.
La boca es terminal (Figura 1a), ligeramente prognata, con los labios relativamente gruesos y con dientes pequeños en los premaxilares y los dentarios. Posterior a la boca se aprecia el reborde cóncavo e inmóvil formado por los bordes anteriores del lacrimal y el hueso suborbital 2 (Figura 7a); anterior a dicho reborde se perciben el bulto del ligamento labial y el extremo inferior del hueso maxilar.

Figuras 1a-c Cavidad bucofaríngea de Chirostoma humboldtianum (Valenciennes, 1835). a) Vista lateral izquierda de la región cefálica. b) Corte sagital de parte de la cavidad bucofaríngea, mostrándose, además, las branquias y el aparato hioideo. c) Corte sagital de la totalidad de la cavidad bucofaríngea. La flecha grande señala la placa dentaria del ceratobranquial 4 y las flechas pequeñas las placas faríngeas superiores y ventral.

Figuras 2a-d Tubo digestivo de Chirostoma humboldtianum (Valenciennes, 1835). a) Vista lateral izquierda. b) Interpretación del enrollamiento del tubo digestivo. c) Corte sagital del tubo digestivo y el hígado en vista lateral izquierda; la flecha señala el punto de contacto del conducto biliar común con el intestino. d) Vista lateral derecha.

Figuras 3a-d Tubo digestivo e hígado de Chirostoma humboldtianum (Valenciennes, 1835). a) Vista ventral. b) Vista dorsal. c) Vista dorsal del hígado y la vesícula biliar. d) Región perianal.

Figuras 4a-g Esqueleto visceral de Chirostoma humboldtianum (Valenciennes, 1835). a) Vista lateral izquierda de las mandíbulas, serie palatina, suspensión mandibular y serie opercular; la flecha señala la posición del ectopterigoides. b) Vista dorsal de las mandíbulas superiores. c) Vista lateral izquierda de la mandíbula inferior. d) Vista dorsal de parte de la porción dentada del dentario. e) Vista lateral izquierda de la mandíbula superior. f) Vista dorsal de parte de la porción dentada del premaxilar. g) Vista interna de la mandíbula inferior.
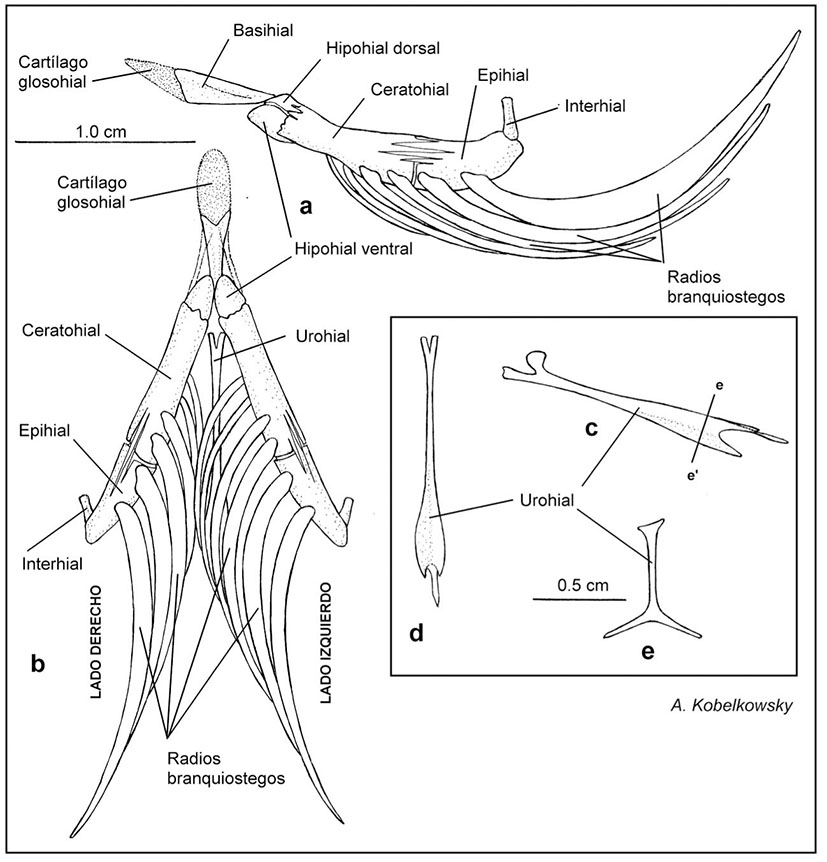
Figuras 5a-e Aparato hioideo de Chirostoma humboldtianum (Valenciennes, 1835). a) Vista lateral izquierda. b) Vista ventral. c) Vista lateral izquierda del urohial. d) Vista dorsal del urohial. e) Sección transversal del urohial.
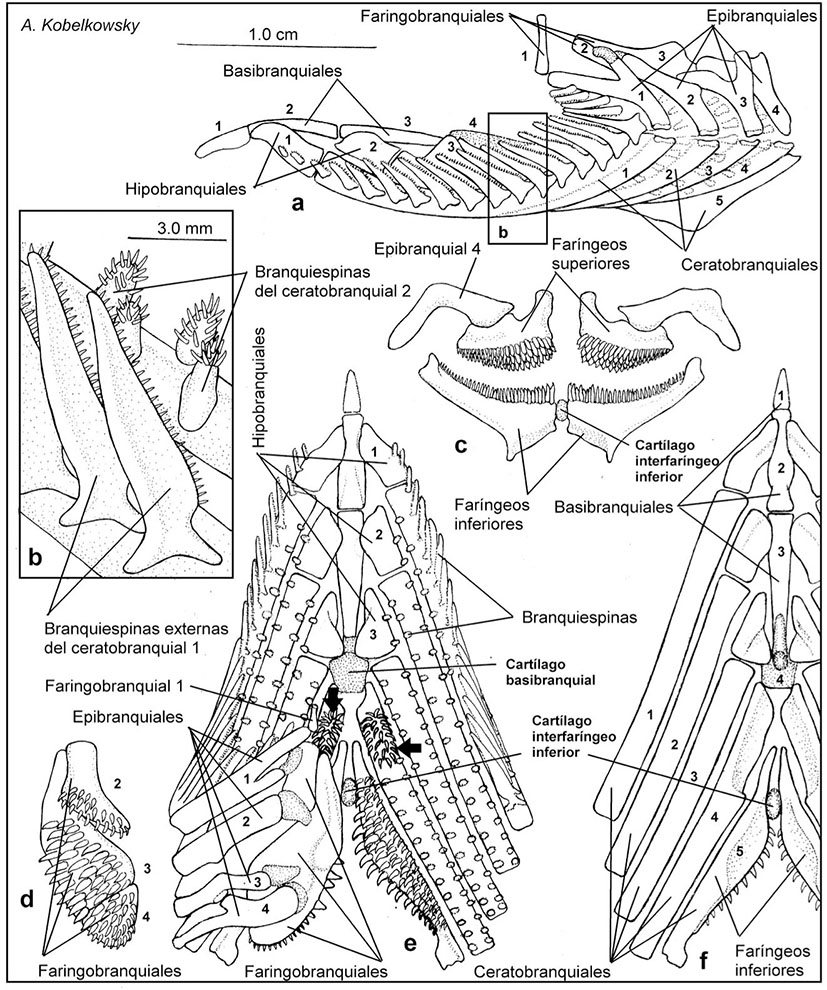
Figuras 6a-f Aparato branquial de Chirostoma humboldtianum (Valenciennes, 1835). a) Vista lateral izquierda. b) Vista lateral de una porción de los arcos branquiales 1 y 2, mostrando las branquiespinas. c) Vista posterior de los huesos faríngeos superiores e inferiores. d) Vista ventral de los huesos faríngeos superiores del lado izquierdo. e) Vista dorsal del aparato branquial, con las ramas superiores derechas de los arcos branquiales removidas; las flechas señalan las placas dentarias del ceratobranquial 4. f) Vista ventral del aparato branquial.

Figuras 7a-e Musculatura mandibular de Chirostoma humboldtianum (Valenciennes, 1835). a) Vista lateral izquierda de la región cefálica después de retirar la piel. b) Vista lateral izquierda del músculo adductor mandibulae, con la sección A1 proyectada. c) Corte transversal del hueso opercular y el músculo levator operculi. d) Vista lateral izquierda del aductor mandibullae, con las secciones A1 y A2 proyectadas. e) Vista lateral izquierda del complejo tendinoso de la sección A1.
Cavidad bucofaríngea. La cavidad bucofaríngea es alargada (Figs. 1b-c). Inmediatamente detrás de la dentición del premaxilar y el dentario se forma la válvula oral con sus componentes superior e inferior. No se encuentran dientes en el paladar y la lengua.
En la porción faríngea (Figura 1b) son notables las branquiespinas del borde externo del primer arco branquial, las cuales son triangulares y de longitud mediana (Figs. 6a, b, e); las branquiespinas del borde interno de este arco, así como también las de ambos bordes de los arcos branquiales 2 a 4 son mas cortas que las del primer arco branquial (Figs. 6b, e) y se intercalan las de un borde con las del borde del arco vecino.
Todas las branquiespinas llevan dientes, así como también los huesos faríngeos superiores e inferiores. Adicionalmente, se localiza una placa dentaria, en la porción anterior de los ceratobranquiales 4 (Figura 6e).
Cavidad visceral. La cavidad visceral es alargada y se extiende más atrás del nivel de los primeros pterigióforos de la aleta anal. Se encuentra delimitada por las vértebras precaudales, las costillas, las cinturas escapular y pélvica, los primeros pterigióforos de la aleta anal y por la musculatura hipaxial. Dentro de esta cavidad se encuentran la vejiga gaseosa, el bazo, las gónadas, el hígado, el esófago y el intestino. La vejiga gaseosa se prolonga hacia atrás hasta ocupar la porción extendida de la cavidad visceral. El riñón mesonéfrico es de carácter extraperitoneal.
Tubo digestivo. El esófago es de mediana longitud (Figs. 1c, 2-3) y de sección ovalada; su superficie interna está formada por pliegues longitudinales finos. El estómago no se encuentra diferenciado, por lo que el intestino se inicia a continuación del esófago; lo cual implica que el conducto biliar común se conecte al intestino, cerca del término del esófago (Figura 2c).
El intestino es relativamente grueso (Figs. 2-3); la primera porción del mismo es ligeramente más amplia que el resto del órgano y forma un ángulo recto con relación al esófago. El resto del intestino desarrolla dos asas (Figs. 2b-c), orientándose la anterior hacia la parte dorsal y la posterior hacia atrás. La superficie interna del intestino tiene un aspecto finamente reticulado y está tapizada por papilas esbeltas; su longitud corresponde a 0.5 veces la longitud patrón del pez. No se forman ciegos pilóricos ni válvula intestinal. El recto tiene surcos finos en su superficie interna y termina en el ano. El ano se abre en posición anterior a la papila urogenital (Figura 3d).
El hígado (Figs. 2-3) muestra el lóbulo hepático derecho más amplio que el izquierdo. La vesícula biliar se ubica entre ambos lóbulos hepáticos (Figs. 3a, c). El conducto del colédoco o conducto biliar común se conecta al intestino inmediatamente después del esófago.
Esqueleto visceral. El esqueleto visceral o branquiocráneo está formado en cada lado por las siguientes series o conjuntos de huesos: las mandíbulas superior e inferior, la serie palatina, la suspensión mandibular, el aparato hioideo, la serie opercular y el aparato branquial. Algunos de los elementos esqueléticos se interconectan mediante cartílagos y ligamentos.
La mandíbula superior está formada en cada lado por el premaxilar y el maxilar, mientras que la mandíbula inferior está constituida por el dentario, el ánguloarticular, el retroarticular y el coronomeckeliano (Figura 4).
El premaxilar posee un agudo proceso ascendente, que se intercala entre los procesos articulares del maxilar (Figura 4b), mientras que su porción posterior es notablemente amplia y redondeada (Figs. 4a, e). Entre ambos premaxilares se localiza el cartílago rostral (Figura 4b). El maxilar es delgado y marcadamente curvado, su porción anterior o cabeza desarrolla el proceso articular dorsal y el ventral, los cuales son largos y agudos. Entre la superficie interna del maxilar y la externa del dentario se encuentra el ligamento coronomaxilar. Entre el maxilar y el etmoides se extiende el ligamento etmo-maxilar, mientras que entre los procesos anteriores dorsales de ambos maxilares se forma el ligamento intermaxilar.
El dentario es robusto y lleva en su superficie ventrolateral una rama del sistema sensorial de la línea lateral con cerca de seis orificios (Figs. 4a, c); su proceso coronoides es amplio y redondeado; en su borde labial lleva una banda de dientes; en su escotadura posterior se intercala el ánguloarticular. El ánguloarticular es considerablemente angosto en su mayor longitud (Figs. 4c, g); lleva cerca de su extremo posterior la faceta articular que recibe al cuadrado, posterior a la cual un pequeño proceso recibe desde el preopercular al ligamento preopérculo-retroarticular; en su superficie interna se encuentra el cartílago de Meckel, que es largo y cilíndrico. El coronomeckeliano es lanceolado y es el elemento más pequeño de la mandíbula inferior; se localiza en la superficie interna del ánguloarticular, cerca del cartílago de Meckel. El retroarticular es un hueso pequeño, unido firmemente a la porción posteroventral del ánguloarticular; recibe desde el interopercular al ligamento interoperculo-retroarticular.
La serie palatina (Figura 4a) está formada en cada lado por los huesos palatino, endopterigoides, metapterigoides y ectopterigoides. El palatino es robusto, tiene un proceso anterior y otro dorsal, y se prolonga hacia atrás mediante la espina posteroventral; lleva en su parte superior el ligamento etmo-palatino y el ligamento maxilo-palatino. El hueso más amplio es el endopterigoides, el cual es laminar y de forma oval; recibe en parte de su superficie dorsal al músculo adductor arcus palatini. El hueso más pequeño de la serie palatina es el ectopterigoides, el cual se sobrepone por la superficie interna de la serie palatina a los huesos palatino, endopterigoides y cuadrado.
La suspensión mandibular (Figura 4) está constituida por los huesos hiomandibular, metapterigoides, simpléctico y cuadrado. El hiomandibular tiene en su porción superior dos cóndilos, que se articulan respectivamente a los huesos esfenótico y pterótico, y otro cóndilo que se articula con el hueso opercular; en su superficie lateral externa se inserta el músculo levator arcus palatini, el cual está limitado posteriormente por un reborde vertical del mismo hiomandibular. Asimismo, el hiomandibular constituye la suspensión del aparato hioideo, mediante el hueso interhial y un corto ligamento (interhial-hiomandibular). El metapterigoides es laminar, irregular y se sobrepone externamente a la parte posterior del endopterigoides, manteniendo un espacio para una porción del músculo adductor arcus palatini. El simpléctico es laminar, más amplio que el metapterigoides y se une por sutura al cuadrado. El cuadrado tiene su porción anterior triangular y la posterior rectangular, la cual se adosa ampliamente al preopercular; en su ángulo anteroventral se forma el cóndilo que se articula al ángulo articular. El opérculo o serie opercular (Figura 4a) está formado por los huesos preopercular, opercular, interopercular y subopercular. La superficie interna de los huesos interopercular y subopercular recibe al conjunto de radios branquiostegos. El preopercular es amplio y de forma triangular, lleva una amplia rama del sistema sensorial de la línea lateral, anterior a la cual se origina parte del músculo adductor mandibulae; dicha rama es continua con la de la mandíbula inferior. El opercular tiene su borde superior ligeramente ondulado y forma un agudo proceso dorsal en el que se inserta el músculo dilatator operculi (Figura 7a). El interopercular es alargado y forma anteriormente un delgado proceso que mediante el ligamento interopercular-retroarticular se une al retroarticular (Figura 4a). El subopecular es curvo y forma un pequeño proceso que se intercala entre el preopercular y el opercular; su extremo posterosuperior es truncado.
El aparato hioideo (Figura 5) está suspendido desde el extremo de los hiomandibulares y está constituido por las dos barras hioideas, y por dos huesos impares. Cada barra hioidea está integrada por el hipohial dorsal, hipohial ventral, ceratohial, epihial, interhial y los radios branquiostegos 1-6; los huesos impares son el basihial y el urohial.
El basihial está precedido por el cartílago glosohial, formando ambos elementos la lengua (Figs. 1b-c, 5a-b). Cada hipohial ventral recibe ventralmente a un músculo hyohyoideus abductor (Figura 8c). Los ceratohiales son largos y se unen con los huesos vecinos por suturas; cada ceratohial sostiene cuatro radios branquiostegos. Los epihiales son medianos, ligeramente curvados y sostienen a los dos últimos branquiostegos. Los interhiales son pequeños y cilíndricos y se unen al extremo inferior de los hiomandibulares.
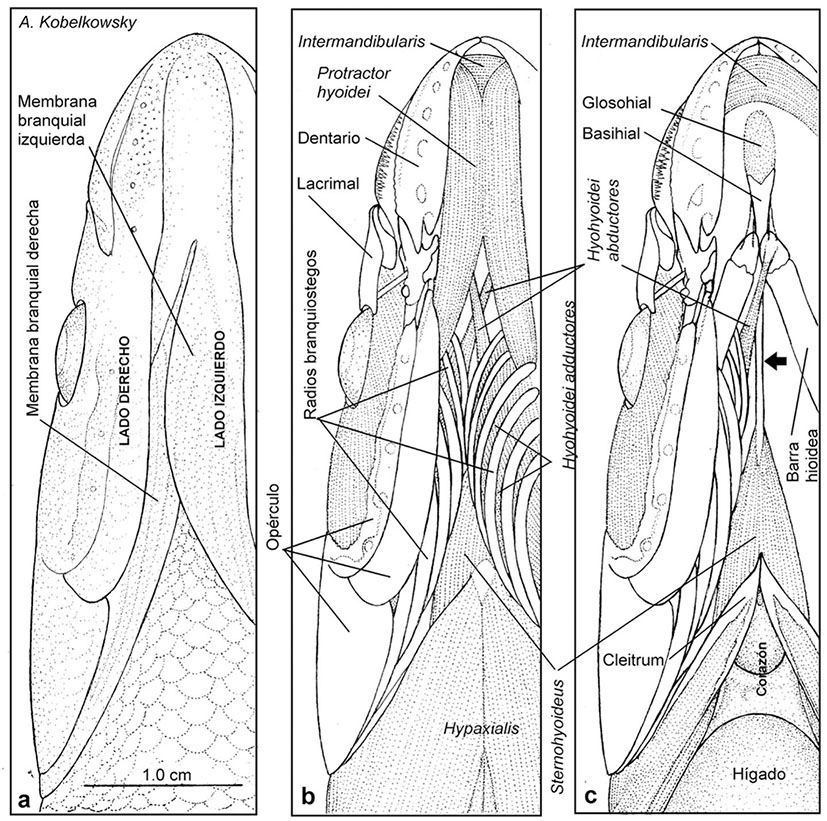
Figuras 8a-c Musculatura hioidea de Chirostoma humboldtianum (Valenciennes, 1835). a) Vista ventral de la región gular, mostrando la asimetría de las membranas branquiales. b) Vista ventral de la musculatura de la región gular después de retirar la piel. c) Vista ventral de la musculatura hioidea después de retirar los músculos protractores hyoidei y los radios branquiostegos izquierdos; la flecha señala el urohial.
El urohial es notablemente delgado y forma en su parte anterior un proceso ascendente, que recibe al músculo rectus communis. La sección transversal de su porción posterior tiene forma de T invertida y recibe al músculo sternohyoideus.
Los radios branquiostegos aumentan gradualmente su amplitud de adelante hacia atrás; mientras que los dos primeros branquiostegos son delgados y se unen a la cara interna del ceratohial, los restantes son más amplios y se unen a la superficie externa del ceratohial y el epihial.
El aparato branquial (Figura 6) está suspendido desde la región ótica del neurocráneo por los faringobranquiales 1 y está constituido por cinco arcos branquiales, formados a su vez por los huesos impares basibranquiales 1-4, y los huesos pareados hipobranquiales 1-3, ceratobranquiales 1-5, epibranquiales 1-4 y faringobranquiales 1-4.
Los basibranquiales son elementos esbeltos, de los cuales el más largo es el tercero y el más corto es el primero. El basibranquial 4 se curva hacia abajo y sirve de punto de inserción de los músculos obliquui ventrales III. Posterior al basibranquial 4 se localiza el cartílago basibranquial.
Los hipobranquiales son cortos y muestran su superficie ventral cóncava, de la cual se originan los músculos obliquui ventrales I, II y III.
Los ceratobranquiales son los elementos más largos del aparato branquial; los ceratobranquiales 1 a 4 son acanalados ventralmente para el paso de las arterias branquiales aferentes; los ceratobranquiales 4 muestran una amplia concavidad ventral para la inserción del músculo transversus ventralis IV, los ceratobranquiales 5 están modificados en los huesos “faríngeos inferiores”, los cuales en la superficie dorsal llevan dientes, y en la ventral muestran una concavidad donde se inserta el músculo transversus ventralis V (Figura 9e). Entre los faríngeos inferiores se localiza el cartílago interfaríngeo inferior.

Figuras 9a-e Musculatura branquial de Chirostoma humboldtianum (Valenciennes, 1835). a) Vista lateral izquierda de los músculos branquiales y hioideos. b) Vista lateral izquierda del músculo retractor dorsalis. c) Vista posterior de los músculos posteriores del aparato branquial. d) Vista dorsal de los músculos intrínsecos dorsales de los arcos branquiales. e) Vista ventral de los músculos intrínsecos ventrales del aparato branquial.
Los epibranquiales son de mediana longitud y forman cada uno un proceso dorsal que recibe a un músculo levator externi; sus extremos superiores son de cartílago. Los cartílagos correspondientes a los epibranquiales 2 a 4 se unen a la superficie dorsal de los huesos faringobranquiales, mientras que el del proceso dorsal del epibranquial 1 se une al faringobranquial 2, y se denomina cartílago interarcual.
Los faringobranquiales son de formas irregulares, siendo el primero corto, cilíndrico y constituye la suspensión del aparato branquial, desde la región ótica del neurocráneo.
Dentición. La dentición de C. humboldtianum se localiza en los arcos mandibular y branquiales, estando ausente en el paladar y la lengua. Los dientes mandibulares están implantados en el premaxilar y el dentario configurando bandas relativamente angostas (Figs. 4d, f); son pequeños, puntiagudos y curvados hacia atrás y hacia adentro.
Los dientes de las branquiespinas externas (Figura 6b) son notablemente pequeños y se localizan en el borde interno. Los dientes de las restantes branquiespinas (Figura 6b) son de mediano tamaño y se localizan en su extremo libre.
Es notable la formación de una placa dentaria con dientes más grandes en la porción anterior de cada ceratobranquial 4 (Figura 6e).
Los dientes del borde interno de cada hueso faríngeo inferior (ceratobranquial 5) son notablemente grandes y gruesos, mientras que los restantes disminuyen de tamaño del borde interno al externo (Figura 6e). Los dientes de los faringobranquiales 2, 3 y 4 (faríngeos superiores) son de mediano tamaño, aunque disminuyen gradualmente de talla hacia atrás (Figura 6d).
Musculatura visceral. La musculatura visceral de C. humboldtianum está organizada en los siguientes grupos de músculos: a) los que relacionan las suspensiones mandibulares con las mandíbulas superiores e inferiores (Figs. 7a-b, d), b) los que relacionan el neurocráneo con el paladar, la suspensiones mandibulares y los opérculos (Figura 7a, c), c) los que relacionan el neurocráneo con las ramas superiores de los arcos branquiales (Figura 9a, d) los que relacionan el aparato hioideo con las mandíbulas inferiores y con los cleitra (Figura 9a), e) los que relacionan los cleitra con los huesos faríngeos inferiores y con el urohial (Figs. 9a, f) los que relacionan los huesos faringobranquiales con las primeras vértebras (Figura 9b).
El principal músculo que permite los movimientos de cada mandíbula superior e inferior es el adductor mandibulae, el cual está formado por las secciones A1, A2, A3 (Figura 7) y Aw. Cerca del extremo anterior de cada sección A1 se forma un complejo tendinoso que se ramifica en un tendón largo que se inserta en la superficie interna del lacrimal, otro de mediana longitud que se inserta en el borde posterior del maxilar y otro más corto que se inserta en la superficie externa del ánguloarticular (Figura 7e).
Los músculos que permiten el movimiento del paladar y la suspensión mandibular se originan en el neurocráneo y son en cada lado el adductor arcus palatini que se inserta en el endopterigoides y el metapterigoides, el levator arcus palatini que se inserta en la superficie externa del hiomandibular y el adductor hyomandibulae que se inserta en la superficie interna del hiomandibular.
La depresión mandibular se realiza por el músculo protractor hyoidei, el cual resulta de la fusión del componente izquierdo con el derecho, con el origen en cada ceratohial y la inserción cerca de la sínfisis de los dentarios; sus extremos anteriores están divididos por el paso del músculo intermandibularis (Figs. 8b-c).
Entre los músculos que se originan en el neurocráneo y se insertan en los epibranquiales se encuentran de cada lado los cuatro levatores dorsales externi y el levator posterior (Figura 9a).
El músculo que de cada lado participa en el descenso del piso del aparato branquial es el rectus communis (Figura 9a); se origina mediante un largo tendón en el ceratobranquial 5 y se inserta en el urohial. Los dos músculos que se originan en los ceratobranquiales 5 y que se insertan en los cleitra son los pharyngoclavicularis internus y pharingoclavicularis externus; este último se encuentra dividido en las secciones A y B (Figura 9a).
El músculo sternohyoideus se origina en la porción inferior de los cleitra y se inserta en la porción posterior del urohial (Figura 9a).
Los músculos retractores dorsales se originan en las vértebras 2 a 4 y se insertan en los huesos faringobranquiales 4 (Figs. 1c, 9b).
Aspectos comparativos de la morfología del sistema digestivo de C. humboldtianum con relación al de C. e. estor. Las principales diferencias entre ambas especies son las siguientes: a) La disposición del intestino tanto en el plano vertical como en el longitudinal en C. humboldtianum, mientras que en C. e. estor es solo en el sentido longitudinal (Figura 10a). b) La mayor longitud de ambas asas intestinales en C. humboldtianum (Figura 10a). c) La mayor altura y la menor longitud de los lóbulos hepáticos en C. humboldtianum, e inversamente la menor altura y la mayor longitud en C. e. estor (Figura 10b). d) La amplitud de cada hueso premaxilar principalmente en su extremo posterior en C. humboldtianum (Figura 10c). e) La mayor curvatura del hueso maxilar en C. humboldtianum (Figura 10d). f) La mayor amplitud del dentario (Figura 10e) en C. humboldtianum, debido principalmente a una mayor altura de los procesos coronoides.

Figuras 10a-e Principales diferencias del sistema digestivo entre Chirostoma estor estor Jordan, 1879 y Chirostoma humboldtianum (Valenciennes, 1835). a) Enrollamiento intestinal; vista lateral izquierda. b) Hígado; vista lateral izquierda. c) Premaxilar; vista lateral izquierda. d) Maxilar; vista lateral izquierda. e) Mandíbula inferior; vista lateral izquierda.
Discusión
El conocimiento del sistema digestivo de los teleósteos implica el análisis anatómico de los huesos, cartílagos y ligamentos que conforman la cavidad bucofaríngea, así como también de los músculos que los mueven.
En este trabajo, el análisis anatómico de la organización del sistema digestivo del pescado blanco, C. humboldtianum, permite reconocer un patrón morfológico correspondiente al de los Atherinomorpha, y en particular al de los Atherinopsidae. La organización anatómica del esqueleto, dentición y musculatura viscerales de esta especie corresponde a la principal estrategia trófica de los acantopterigios o teleósteos superiores, que de acuerdo con Vandewalle et al. (2000) consiste en la captura de la presa, que generalmente es por succión y su procesamiento en la cavidad faríngea. En cada una de estas fases se utilizan diferentes sistemas funcionales o unidades mecánicas.
En el presente trabajo a diferencia de otros estudios se consideran las estructuras de los teleósteos superiores o acantopterigios que constituyen las llamadas por Lauder (1982) “unidades mecánicas”. Dicho autor resalta las acciones de apertura de la boca, retractilidad de la mandíbula superior y succión alimenticia.
La organización del esqueleto visceral de C. humboldtianum corresponde al patrón morfológico de los Atherinopsidae, observándose una gran similitud con las descripciones osteológicas de especies sudamericanas (de La Hoz & Aldunate, 1994; Dyer, 1997; Bemvenuti, 2005).
La dentición del adulto de C. humboldtianum corresponde a la de un teleósteo, que incluye en su dieta a otros teleósteos. El conjunto de numerosos dientes pequeños en las branquiespinas de los arcos 1 a 4 y en los huesos faríngeos superiores e inferiores es característica de teleósteos ictiófagos (Moyle & Cech, 2000).
Aunque Soria-Barreto y González-Díaz (2004) registran una placa dentaria en los endopterigoides de Chirostoma attenuatum, y Bemvenuti (2005) en el de varias especies de Odontotesthes, en el presente trabajo no se observaron. Sin embargo, se registra por primera vez la presencia de una placa dentaria en los ceratobranquiales 4. Considerando que como en muchos teleósteos existen dientes en los huesos ceratobranquiales 5 o “faríngeos inferiores”, que proporcionan un área dentada del piso de la faringe, la cual se opone al área dentada del techo de la misma, formada por los huesos faríngeos superiores, el área inferior en C. humboldtianum es incrementada por las placas dentadas de los ceratobranquiales 4. Dicha condición morfológica probablemente permita un mejor manejo de la presa.
La presencia del cartílago “interfaríngeo inferior”, no descrito en otras especies en la literatura, probablemente funcione como una diartrosis que permite movimientos diferenciales de los dos huesos faríngeos inferiores.
En el presente estudio se confirman como principales aspectos miológicos los siguientes: a) el gran desarrollo del músculo levator operculi y el carácter externo de su inserción sobre el hueso opercular registrados previamente por Kobelkowsky (2012) en C. e. estor ; b) la división del músculo adductor mandibulae en las secciones A1, A2, A3 y Aw, c) la presencia ya descrita por autores como de la Hoz (1994) y Dyer (1997) del complejo tendinoso de la sección A1 del músculo adductor mandibulae que lo une a los huesos lacrimal, maxilar y ánguloarticular, d) la división en dos secciones del músculo pharyngoclavicularis externus, que en este estudio se nombran como “sección A“ y sección B”, e) el origen de cada músculo retractor dorsalis en las vértebras 2 a 4 y su inserción en el faringobranquial 4; f) se identifican el músculo transversus dorsalis anterior y el músculo transversus dorsalis posterior, g) el carácter asimétrico de los músculos hyohyoidei abductores, obliquui ventralis III, y transversus dorsalis anterior.
Entre los aspectos morfológicos sobresalientes del tubo digestivo de C. humboldtianum están la ausencia del estómago, de los ciegos pilóricos y de la válvula intestinal. La ausencia del estómago de C. humboldtianum corresponde a la diagnosis del superorden Atherinomorpha (Nelson, 2006), y a la afirmación de Wilson & Castro (2011) sobre la ausencia de esta porción del tubo digestivo en el 7% de las familias de teleósteos y del 20 al 27% de las especies. En el presente trabajo se reconoce que el carácter que determina el inicio del intestino es el nivel de la conexión del conducto biliar común con el tubo digestivo, de la misma forma que Logothetis et al. (2001) lo hace en A. affinis. Estos últimos autores comentan, además la ausencia de secreción del ácido clorhídrico y de pepsinógeno, lo que comprueba la ausencia de estómago.
Buddington y Diamond (1987) asignaron a los ciegos pilóricos la función de aumento de la superficie de absorción; y señalan que es frecuente su ausencia en los teleósteos.
La principal diferencia con relación a los Atherinidae, es la gran amplitud en C. humboldtianum y C. e. estor de la porción posterior del premaxilar y la mayor retractilidad de la mandíbula superior.
La comparación anatómica del sistema digestivo de C. humboldtianum con el de C. e. estor permite identificar algunas diferencias en las proporciones de los huesos premaxilar, maxilar, dentario y del hígado, así como también en el enrollamiento del intestino, las cuales se correlacionan con diferencias de altura del cuerpo entre ambas especies. Así, se reconoce que mientras que el cuerpo de C. humboldtianum es más alto, sus huesos premaxilar y maxilar son más amplios en el sentido vertical, el hígado es más alto y corto y la disposición de su intestino en el plano vertical es mayor que en C. e. estor.
El presente análisis morfológico del sistema digestivo de C. humboldtianum y su comparación con la descripción del mismo sistema en C. e. estor por Kobelkowsky (2012) permite definir las siguientes características de ambas especies y que probablemente correspondan a las del género Chirostoma:
La presencia de una placa dentaria en cada ceratobranquial 4.
La presencia del cartílago “interfaríngeo”.
La amplia inserción del músculo levator operculi (elevador del opérculo) sobre la parte externa del hueso opercular.
La definición de las secciones A1, A2, A3 y Aw del músculo adductor mandibulae (aductor mandibular).
La presencia del complejo tendinoso de la sección A1 del músculo adductor mandibulae.
La presencia de un nuevo músculo o sección muscular del pharyngoclavicularis externus (faringoclavicular externo), no registrado en el resto de los teleósteos, denominado en el presente trabajo como “sección B” del mismo músculo.
La definición del origen de cada músculo retractor dorsalis (retractor dorsal) en los centros vertebrales 2 a 4.
El mayor desarrollo del lóbulo hepático derecho.
Las características anatómicas descritas permiten definir aspectos ecomorfológicos de estas especies, ya que su sistema digestivo está relacionado con el tipo de presas que consume en el período adulto, como son los peces forrajeros y proponer tipos de alimento formulados para peces que carecen de estómago, útiles en sistemas de cultivo con fines de conservación o producción.

