Taxonomía y florística
Description and distribution of Nemacystus howei
(W.R. Taylor) Kylin (Chordariaceae) from the northern coast of the Yucatán
peninsula
Descripción y distribución de Nemacystus howei
(W.R. Taylor) Kylin (Chordariaceae) en la costa norte de la península de
Yucatán
1Departamento de Biología Marina, UADY, Itzimna
Mérida, Yucatán. México
2Centro de Investigaciones Marinas, UH, La
Habana, Cuba.
3UMDI-Sisal, Facultad de Ciencias, UNAM/Escuela
Nacional de Estudios Superiores-Unidad Mérida, UNAM, Yucatán,
México
4Department of Ecology and Evolutionary Biology
and Herbarium, University of Michigan, Ann Arbor, MI, USA.
Abstract
Background:
There are few studies about Nemacystus on the Atlantic
coast, and N. howei has been reported only once for Mexico;
that record, however, did not provide a description of this species.
Question:
What are the morphological characteristics of Nemacystus
howei and its distribution on the Yucatan coast?
Species study:
Macroalgae, Phaeophyceae, Nemacystus howei.
Study site and year of study:
North coast of Yucatán Peninsula of Mexico. Dry and rainy seasons from 2017
to 2019.
Methods:
Sampling was carried out at five localities, at 5 and 10 m depths, with
scuba equipment. Specimens were preserved in diluted formalin in sea water;
silica gel and dried-pressed vouchers are housed at the UADY
Results:
Six samples were collected at the five localities. Each one contains a small
mass of filamentous of specimens of Nemacystus howei, that
were found only during dry season, at both depths sampled. This alga has a
thallus of an entangled filamentous mass of cylindrical, mucilaginous axes,
of a light brown color. Younger branches have a solid central axis.
Branching is abundant. The main, older axes are hollow. The assimilatory
filaments are branched a few times, producing uniseriate colorless hairs and
with plurilocular sporangia arising from the base. Nemacystus
howei is easily confused with Cladosiphon
occidentalis Kylin due to their similar morphology, branching
pattern, and size.
Conclusion:
The significant morphological similarities between these species as well as
the paucity of specific works on the Phaeophyceae of the Yucatan Peninsula
may explain why Nemacystus howei had not been previously
reported.
Keywords: Cladosiphon; Gulf of Mexico; morphology; Phaeophyceae
Resumen
Antecedentes:
Nemacystus es un género poco estudiado y N.
howei solo se ha reportado una vez para México, sin una
descripción de esta especie.
Pregunta:
¿Cuáles son las características morfológicas de Nemacystus
howei y su distribución en la costa de Yucatán?
Especie de estudio:
Macroalgas, Phaeophyceae, Nemacystus howei.
Sitio y años de estudio:
Costa norte de la península de Yucatán. Temporadas de secas y lluvias, de
2017 a 2019.
Métodos:
Se muestrearon 5 sitios a lo largo de la costa a 5 y 10 metros de
profundidad, con equipo de buceo autónomo. Los especímenes se conservaron en
formalina al 4 %, sílica gel y en cartulina y se depositaron en el herbario
de la UADY Mérida.
Resultados:
Se colectaron 6 muestras en cinco localidades cada una contenía unos
filamentos de especímenes de Nemacystus howei. Solo se
encontró durante la época de seca, a ambas profundidades. Se presenta como
una masa de filamentos cilíndricos, y mucilaginosos, de color café claro.
Los ejes principales son huecos y las ramas juveniles tienen eje central
sólido. La ramificación es abundante. Los filamentos asimiladores son poco
ramificados, producen pelos incoloros uniseriados y la esporangia
plurilocular surge de su base. Nemacystus howei, se
confunde fácilmente con Cladosiphon occidentalis Kylin,
debido a su morfología, patrón de ramificación y tamaño similares.
Conclusión:
Se considera que, por la gran similitud morfológica con C.
occidentalis y la falta de trabajos especializados sobre el
grupo Phaeophyceae, es posible que por ello no se había reportado
anteriormente para la Península de Yucatán.
Palabras claves: Cladosiphon; Golfo de México; morfología; Phaeophyceae
Nemacystus is a genus in the family Chordariaceae (Order Ectocarpales,
Class Phaeophyceae) and contains 8 species (Guiry &
Guiry 2020). The type species of the genus is N. ramulosus
Derbès & Solier. There are relatively few studies on Nemacystus,
the most complete one being that of de Jong &
Prud'homme (1997), who did a review of the genus based on morphological
characters and presumed phylogenetic relationships. A molecular phylogeny, however, has
not yet been carried out for this genus. Species differ from each other on such subtle
morphological traits as the dimensions and shape of the assimilatory filaments,
plurilocular sporangia, color, ramification, and general habit (Jong & Prud'homme 1997).
In North America only two species have been reported, Nemacystus
brandegeei (Setchell & Gardner) Kylin on the Pacific coast (Pedroche et al. 2008, Norris 2010) and Nemacystus howei
(W.R. Taylor) Kylin on the Atlantic coast. On the Atlantic coast N.
howei has been reported and described from Florida, North Carolina, and
Bermuda (Taylor 1928, 1960, Schneider & Searles
1975, 1991, Schneider 2003). In Brazil, this species was recorded by Cassano & Pedrini (1992), which was the first
report for the Atlantic coast of South America. Menezes
& Campos de Paula (2010) later reported it from the southeastern coast of
Brazil. It was reported and described from Florida by Taylor (1928, 1960) and Dawes & Mathieson (2008); for North Carolina it
was reported and described by Schneider & Searles
(1991), and for the Caribbean islands of Aruba and Bonaire by Vroman & Stegenga (1988). For the Gulf of
Mexico, Fredericq et al. (2009)
reported N. howei as a synonym of “Eudesme howei”
Taylor nom. inval., citing the report of Phillips &
Springer (1960) for the Gulf coast of Florida. Finally, for the Atlantic
coast of Mexico, Mateo-Cid et al.
(2013) reported N. howei as a new record from the subtidal
benthic macroalgae from the Campeche Banks.
Almost all the previous works with descriptions showed only illustrations of the
species, especially of the assimilatory filaments and the plurilocular sporangia, except
that of Cassano & Pedrini (1992), which also
included photos of transverse sections of the thallus, depicting the central axis to be
solid in younger parts and hollow in older parts. For the Atlantic coast of Mexico,
however, despite being the first report, a description of the morphological
characteristics or figures of this species has not yet been given for Mexico. So, this
report is the first morphological descriptive account of this species in Mexico, and we
provide the distribution of Nemacystus howei for the northern coast of
the Yucatan Peninsula.
Materials and methods
Sampling was carried out on the northern coast of Yucatan during the dry season (with
a 26.6 °C temperature average, 38 UPS and 8.8 pH) and the rainy season (with a 29.4
°C temperature average, 36 UPS and 7.9 pH ) at five localities: Ría Lagartos, 21°
38' 31.5'' N; 88° 10' 19.8'' W, San Felipe 5m (21° 37.080' N; 88°13.818' W), San
Felipe 10 m (21° 39.344' N; 88° 16.344' W), Coloradas 10 m (21° 43' 17.2'' N; 87°
58' 187'' W), Chicxulub 10 m (21° 32.284' N; 89° 30.973' W), “Bajos 10” 10m (21°
21.210 N; 90° 09.125 W) and “Bajos 10” 5m (21° 20.870 N; 90° 08.940 W.) at Sisal
reefs (Figure 1) from 2017 to 2019 at two
depths (5 and 10 m) in the sublittoral, using SCUBA equipment. Specimens were
separated, labeled, and preserved in a 4 % formaldehyde solution in seawater, silica
gel and two dry vouchers were made from each sample, which are housed at the
Herbarium Alfredo Barrera Marin of the University Autonomous of Yucatan (UADY). The
determination was made using the keys and descriptions of Taylor (1960), Schneider &
Searles (1991), Cassano & Pedrini
(1992), and Dawes & Mathieson
(2008).
Results
Six samples were collected at the five localities and each one contains a small mass
of filamentous specimens of the species Nemacystus howei (W.R.
Taylor) Kylin 1940, that belongs to: Kingdom
Chromista, Phylum Ochrophyta, Class Phaeophyceae, Order Ectocarpales, Family
Chordariaceae, Genus Nemacystus Derbès & Solier, 1850.
Basionym. Castagnea howei W.R. Taylor 1928: p. 111, Reported by Taylor
1960: p. 249, Earle 1969: p. 184,
Dawes 1974: p. 107, Schneider & Searles 1991: p.138, Dawes & Mathieson 2008: p.145. Type locality: Dry Tortugas,
Florida. Distribution in the Western Atlantic: The Carolinas, Florida, northern Gulf
of Mexico, Gulf of Mexico, Brazil, Bermuda. Distribution on the northern coast of
Yucatán Ría Lagartos, San Felipe, Coloradas, Chicxulub, and “Bajos 10” at Sisal
reefs and Telchac (Figure 1). We observed
N. howei to occur only in the dry season
between March-June. It can be epiphytic or saxicolous, from 5 to 10 m depths (Table 1).
Table 1 Material examined.
| Date |
Localities |
Genus |
species |
I.D Number |
Depth (m) |
| 06/05/2017 |
Coloradas |
Nemacystus |
howei |
UADY 1594 |
10 |
| 06/05/2017 |
Ría Lagartos |
Nemacystus |
howei |
UADY 1595 |
5 |
| 06/05/2017 |
San Felipe |
Nemacystus |
howei |
UADY 1596 |
5 |
| 06/05/2017 |
San Felipe |
Nemacystus |
howei |
UADY 1597 |
10 |
| 07/06/2019 |
Chicxulub |
Nemacystus |
howei |
UADY 1598 |
10 |
| 15/06/2019 |
Bajos de 10 |
Nemacystus |
howei |
UADY 1599 |
5 |
| 28/04/2017 |
Bajos de 10 |
Cladosiphon |
occidentalis |
UADY 1593 |
10 |
| 17/10/2010 |
Telchac |
Nemacystus |
howei (reported as
brandegeei) |
UADY 1136 |
- (Epiphyte of Sargassum) |
Examined Material. We collected six samples of specimens of
Nemacystus howei (Table
1): UADY 1594 at Coloradas, UADY 1595 at Ría Lagartos, UADY 1596 at San
Felipe, UADY 1597 at San Felipe, UADY 1598 at Chicxulub, UADY 1599 at “Bajos de 10”
(Sisal Reefs).
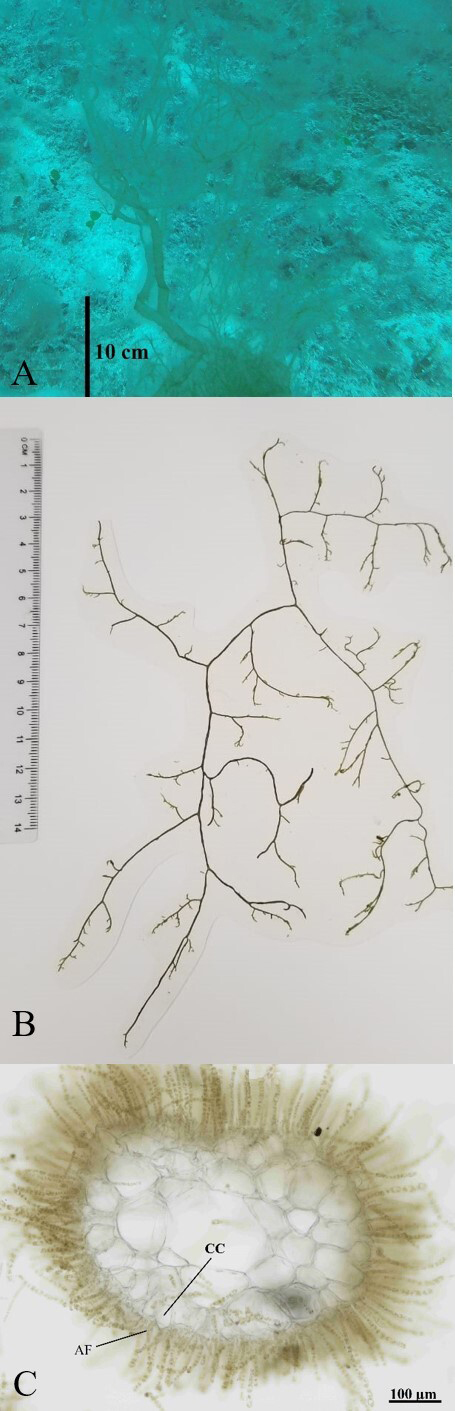
Description. Thallus with a filamentous form, soft and mucilaginous
entangled masses, up to 45 cm long, color light brown and somewhat greenish (Figure 2A-B). The axis is cylindrical, and the
uniaxial structure is visible proximal to the apical cells. Branching is abundant,
up to three orders, alternate to irregular. The internal structure is differentiated
between a medulla (with 2-4 rows of colorless cells) and cortical cells. The main
and older axes become hollow with a diameter between 0.3 - 0.6 mm (Figures 2C, 3A); the younger branches have a
solid central axis, thin, cylindrical and often entangled, 0.1 - 0.3 mm in diameter
(Figure 3C). The medullary cells are from
55-120 by 30-60 μm in diameter. The cortical cells are 12-25 by 12-45 μm in
diameter.
The assimilatory filaments are not branched, 6-8(-13) cells long, with rounded
moniliform or reniform cells, 10-15 μm near the apices and to 4-6 μm below. There
are several discoid plastids per cell. These cells produce colorless hairs, 8-10 μm
in diameter (Figure 3C). Unilocular sporangia
were not observed. Plurilocular sporangia are uniseriate, arising from the base of
the assimilatory filaments or directly from the cortical cells; 40-80 μm long with
8-23 loculi, which are 4-6 μm in diameter (Figure
3D).
Discussion
The review of the morphological descriptions of the specimens of Nemacystus
howei by previous authors (Taylor
1960, Schneider & Searles
1991, Cassano & Pedrini 1992, de
Jong & Prud’homme 1997, Dawes & Mathieson 2008), with respect to
what was found in Yucatan, are very similar in size.
The reports of N. howei from the northern Gulf of Mexico, Florida,
and Bermuda, showed a thallus length of approximately 45 cm, which is like our
specimens, and with a diameter similar to those values for North Carolina specimens,
as reported by Schneider & Searles
(1991), who found specimens to be slender with main axes not exceeding a
diameter of 0.5 mm. Also, our specimens were found at lower depths than other
reports (Table 2 in Appendix 1).
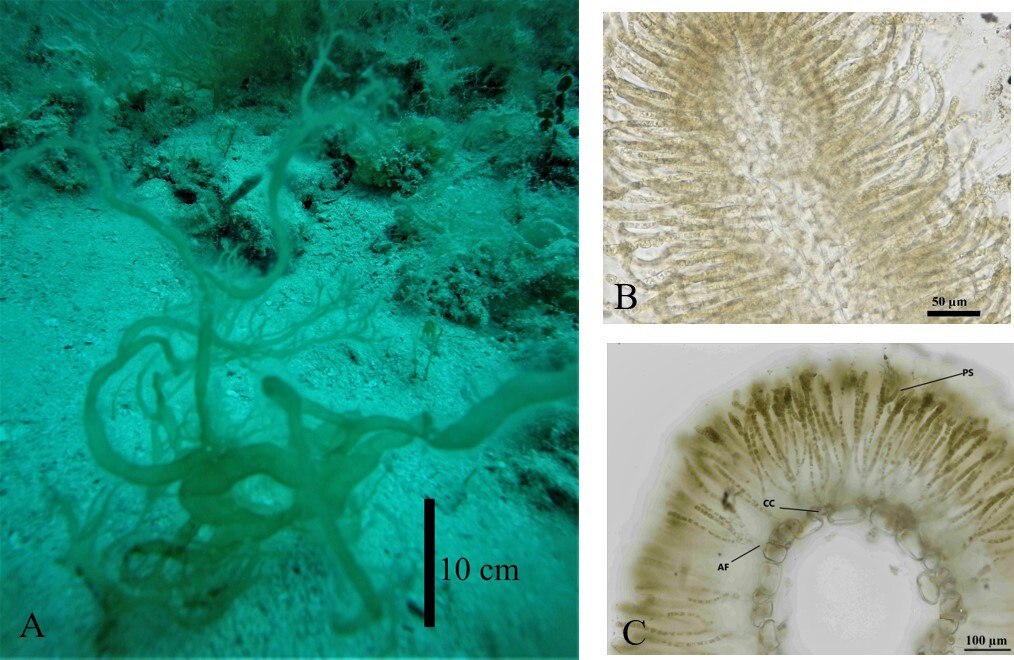
Nemacystus howei (Figure 2A,
3C) has often been misidentified and easily confused with Cladosiphon
occidentalis Kylin (Kylin 1940,
Schneider & Searles 1975, de Jong & Prud’homme 1997) because of their
very similar external morphology, ramification, and size (Figure 4A-B).
We found C. occidentalis (UADY 1593) at one locality (“Bajos de 10”
at Sisal Reef) (Table 1), and it was
necessary to make transverse sections to verify the identity. The main differences
are the solid medulla of the younger branches and more medullary cells in the older
branches of N. howei, whereas the cortex in
C. occidentalis is formed of only one row of cells. The
assimilatory filaments in C. occidentalis have a pedicel and then
bifurcate two or three times; also, the sporangia are formed at the end of the
filaments (Figure 4C), whereas in N.
howei the filaments are uniseriate and the sporangia arise at the base
of the filaments or directly on the cortical cells (Figure 3A).
At the UADY herbarium we found a single collection that had been identified as
N. brandegeei (Setchell & Gardner) Kylinin Telchac, Yucatan
(Figure 1), but it was misidentified and
turned out to be N. howei (Table
1). There were no vouchers of Cladosiphon besides the
ones that we collected.
Even though the external morphologies of these two species are very similar, it is
relatively easy to distinguish them when their internal characteristics are checked.
Their external resemblances have likely resulted in past confusion in their accurate
determinations. Also, the paucity of specific works on the Phaeophyceae for the
Yucatan Peninsula is another probable explanation why this is the first report of
N. howei for the Yucatan Peninsula.
Acknowledgments
We express our gratitude to the Universidad Nacional Autónoma de México which
supported the Program for Projects for the Innovation and Improvement of Education
PAPIME PE207317, which enabled us to carry out this project. Also, our thanks to Dr.
Xavier Chiappa Carrara, Chairman of the scientific network RECORECOS (Red para el
Conocimiento de los Recursos Costeros del Sureste) for supporting logistics.
Finally, we thank Dr. Armin Tuz, M.Sc. Johnny Valdez, and the students Mariana
Álvarez, Fernando Ayuso, Valery Ávila, and Karla Durán for their support in the
field.
Literature cited
Cassano V, Pedrini AG. 1993. Nemacystus howei
(Taylor) Kylin (Phaeophyta, Spermatochnaceae): nova referéncia para a flora
marinha Atlántica Sul-Americana. Hoehnea
20: 93-100.
[ Links ]
Dawes CJ, 1974. Algae of the West Coast of Florida.
USA: University of Miami Press. ISBN-10 :9780870242588
[ Links ]
Dawes CJ, Mathieson AC. 2008. The seaweeds of
Florida. Florida: University Press of Florida. ISBN:
978-0-8130-3148-4
[ Links ]
Earle SA. 1969. Phaeophyta of the eastern Gulf of Mexico.
Phycologia
7: 71-254. DOI: https://doi.org/10.2216/i0031-8884-7-2-71.1
[ Links ]
Fredericq S, Cho TO, Earle SA, Gurgel CF Krayesky DM Mateo-Cid LE,
Mendoza-González AC, Norris JN, Suárez AM. 2009. Seaweeds of the Gulf of Mexico,
In: Felder D.L, D.K. Camp, eds. Gulf of
Mexico-Origins, Waters, and Biota. I. Biodiversity. Texas: Texas
A&M Press, pp. 187-259. ISBN-13: 978-1603440943
[ Links ]
Guiry MD, Guiry, GM. 2020. AlgaeBase. World-wide
electronic publication, National University of Ireland, Galway. http://www.algaebase.org (accessed April 14,
2020).
[ Links ]
Jong YSDM de, Prud'homme van Reine WF. 1997. A review of the genus
Nemacystus (Spermatochnaceae, Chordariales, Phaeophyceae),
including phylogenetic and biogeographical hypothesis. Nova
Hedwigia
64: 1-40.
[ Links ]
Kylin H. 1940. Die Phaeophyceenordnung Chordariales. Acta
Universitatis Lundensis
36: 1-67.
[ Links ]
Mateo-Cid LE, Mendoza-González C, Fredericq S. 2013. Lista de algas
marinas submareales de los bancos de Campeche, México. Acta Botanica
Venezuelica
36: 95-108. DOI: https://doi.org/10.21829/abm104.2013.57
[ Links ]
Menezes Széchy MT, Campos de Paula J. 2010. Phaeophyceae.
In: Forzza RC, ed. Catálogo de Plantas e Fungos do
Brasil. Vol. 1. Rio de Janeiro: Andrea Jakobsson Estúdio; Instituto
de Pesquisas Jardim Botânico do Rio de Janeiro, pp. 404-408. ISBN:
978-85-88742-42-0.
[ Links ]
Norris JN. 2010. Marine algae of the Northern Gulf of California:
Chlorophyta and Phaeophyceae. Smithsonian Contributions to
Botany
94: 1-276. DOI: https://doi.org/10.5479/si.0081024X.94.276
[ Links ]
Pedroche PF, Silva PC, Aguilar Rosas LE, Dreckmann KM, Aguilar Rosas
R. 2008. Catálogo de las algas benthónicas del Pacífico de México II.
Phaeophycota. México, D.F: Universidad Autónoma Metropolitana.
ISBN: 978-970-31-0909-8
[ Links ]
Phillips RC, Springer VG. 1960. Observations on the offshore benthic
flora in the Gulf of Mexico off Pinellas County, Florida. American
Midland Naturalist
64: 362-381. DOI: https://doi.org/10.2307/2422669
[ Links ]
Schneider CW. 2003. An annotated checklist and bibliography of the
marine macroalgae of the Bermuda Islands. Nova Hedwigia
76: 275-361. DOI: https://doi.org/10.1127/0029-5035/2003/0076-0275
[ Links ]
Schneider CW, Searles RB. 1975. North Carolina marine algae, IV.
Further contribution from the continental shelf, including two new species of
Rhodophyta. Nova Hedwigia
26:83-103.
[ Links ]
Schneider CW, Searles RB. 1991. Seaweeds of the southeastern
United States. Cape Hatteras to Cape Canaveral. Durham &
London: Duke University Press. ISBN: 0-8223-1101-1.
[ Links ]
Taylor WR. 1928. The marine algae of Florida with special
reference to the Dry Tortugas. USA: Publications of the Carnegie
Institution of Washington.
[ Links ]
Taylor WR. 1960. Marine algae of the eastern tropical and
subtropical coasts of the Americas. Ann Arbor: The University of
Michigan Press.
[ Links ]
Vroman M, Stegenga H. 1988. An annotated checklist of the marine
algae of the Caribbean islands of Aruba and Bonaire. Nova
Hedwigia
46: 433-480.
[ Links ]
Appendix 1. Morphological characters of Nemacystus
howei by different authors.
|
Taylor 1960
|
Schneider & Searles
1991
|
Cassano & Pedrini
1992
|
Jong & Prud’homme
1997
|
Dawes & Mathieson
2008
|
Ortegon-Aznar et al.
this work |
| Habit |
Entangled with other algae |
In tangled masses |
Filamentous, twisted and matted with other
algae. |
Much entangled. dark brown |
Attached by a rhizoidal holdfast, epiphytic,
entangled or on limestone |
Filamentous, twisted and matted with other
algae. |
| Depth (m) |
7-55 |
15-25 |
- |
Bermuda: 54-64, Florida: 7.2-34.2, North
Carolina: 15-25. |
13- 55 |
5-10 |
| Distribution months |
- |
May-August, November. |
October- December |
- |
- |
March-June |
| Size (cm) |
20-40 |
40 |
35 |
20-50 |
40 |
20-45 |
| Branches ramification |
Alternate abundant |
Alternate abundant, to 3 orders |
Irregular abundant rarely alternate |
Abundant irregular, occasionally dichotomous,
Alternate to 3 orders |
Alternate to irregular abundant, to 3
orders |
Alternate to irregular abundant, to 3
orders |
| Cortex |
- |
- |
Solid at apical zones and hollow in
intermediate part |
Central axis solid, hollow in older
portions |
- |
Solid at apical zones and hollow in older
parts |
| Cortex diameter (mm) |
0.3-1 |
to 1 |
- |
0.3- 1 |
to 1 |
0.3-.06 |
| Medullar cells (μm) wide by long |
58-166 x 132-1410 |
30-166 by 132-1410 |
48-91 by 26.5-60 |
30-166 by132-1410 |
60-170 by 130-1400 |
55-120 by 30-60 |
| Cortical cells (μm) wide by long |
- |
- |
19-39 by 12-27 |
22 x 14 |
- |
12-25 by 12-45 |
| Assimilatory filaments Shape |
Cylindrical below moniliform or reniform
above unbranched |
Cylindrical below moniliform or reniform
above - |
|
Cylindrical below moniliform or reniform
above. Simple, in small bundles at the base (Branched
at the base)
|
rounded cells unbranched |
|
|
Assimilatory filaments Number of cells long
|
8 |
8-13 |
(6-) 8-12 (-16) |
(6-)7-9 (-12) |
8-13 |
6-8 (-13) |
| Assimilatory filaments Size diam (μm) |
10-16.5 toward their summits 3.2-8 toward the
bases |
10-17 near apices |
12-26 apical cell |
16-19 apical cell |
10-15 apical cells |
10-15 apical cells |
| Sporangia Plurilocular Shape |
- |
Uniseriate-bi seriate |
Single, rarely branched, straight or curved,
with pedicel |
Single or branching pedicel or without
pedicel |
Uniseriate-bi seriate, Single or branching
pedicel |
Single, rarely branched, straight or
curved, |
| Sporangia size long (μm) |
|
37-68 |
29-77 |
30-80 |
40-80 |
40-80 |
| No of sporangia per fascicle |
to 10 |
to 10 |
to 15 |
to 10 |
to 6 |
To 6 |
| Sporangia Size diam (μm) |
6.6-7.5 |
6-8 |
5-7 |
4.6-8.3 |
6-8 |
4 -6 |
| Loculi |
- |
- |
- |
8-20 |
- |
8-23 |
| Unilocular sporangia Shape |
Spherical to ovoid |
Spherical to ovoid |
Spherical to ovoid |
Spherical to oval regular or
asymmetrical |
ovoid |
- |
| Unilocular sporangia size (μm) |
18-37 |
18-37 |
49-62 x 17-40 |
20-50 x 18-39 |
20-40 |
- |
| Phaeophycean or colorless hairs, diam (μm) |
8-10 |
8-10 |
8-10 |
- |
8-10 |
8-10 |












 nova página do texto(beta)
nova página do texto(beta) Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
Permalink