











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCTION
Rheumatoid arthritis (RA) is a complex systemic autoimmune disease characterized by chronic synovial joint inflammation that leads to the progressive destruction of articular and periarticular structures, disability, and lessened quality of life1. The main clinical feature is polyarthritis, which affects the small joints symmetrically, frequently with altered parts of the musculoskeletal system and extra-hematological manifestations2. During the active phase of RA, elevated values of erythrocyte sedimentation rate (ESR) and C-reactive protein (CRP) can be detected. At the same time, the definitive diagnosis is confirmed by the presence of the rheumatoid factor (RF) and anti-citrullinated protein/peptide autoantibodies (ACPAs)3. The etiology of RA remains undetermined. However, the combined effect of genetic, epigenetic, and environmental factors is concurrent with the disease’s predisposition, pathogenesis, and severity4. Premature immune aging is characteristic of RA patients and is commonly present before symptoms onset5. One of the most distinctive markers of cellular senescence is telomere shortening6. Telomeres are unique structures of assembled repetitive TTAGGG DNA sequences and related proteins located at the end of human chromosome arms. Their primary function is the maintenance of chromosome structure integrity7. The telomere DNA is shortened by 40-200 bp in each cell division8. When telomeric DNA becomes critically short, the cell becomes senescent and enters apoptosis9. Some human cell types with a high cell turnover, including hematopoietic stem cells and peripheral lymphocytes, have functional, active telomerase, a large ribonucleoprotein complex that repairs and extends short telomeres prolonging cell life10. Telomere length (TL) has a high interindividual variation at birth and in adult life, which seems to be genetically determined, presenting high heritability (h2 = 0.70) and a positive correlation with paternal age at conception11. Age-associated telomere shortening is a phenomenon with similar pattern rates in the first two decades of life12. However, the shortening rate in adults is variable and seems to be influenced by environmental factors, even in monozygotic twins13. TL in different subsets of white blood cells, skin cells, umbilical artery cells, vascular wall, liver, spleen, and kidney tissues of the same individual varies with age in a synchronized manner14. Several studies showed the association between telomeres shortening in RA patients and short telomeres found in multiple cell types, including lymphocytes T and B, and neutrophils5,15,16. Moreover, the peripheral hematopoietic precursor cells and bone marrow mesenchymal stem cells from patients with RA usually show shorter TL than healthy individuals of the same age17. Genetic predisposition to RA may also influence TL. Schönland et al. observed that the HLA-DRB1*04 gene allele, a genetic significant factor risk for RA, contributes to premature telomere shortening in peripheral white blood cells5. RA patients carrying HLA-DRB1*04 present premature telomere shortening more frequently than healthy controls, suggesting the participation of this gene in cell senescence. Thus, telomere shortening may appear before RA onset in genetically predisposed individuals.
It is unclear if the telomere shortening is a consequence of the disease or if TL reduction precedes disease development. Probably, it is simultaneous with disease progression. Studies evaluating TL before the onset of clinical symptoms of RA are scarce. Prescott et al., studying incident RA cases in women from three different cohorts with blood samples collected at least 3 months before symptoms onset, did not find telomere shortening before disease onset compared to age-matched controls, suggesting a disease consequence more than a predisposing risk factor18. The present study investigates the relative TL from the early stage of the disease and length changes after follow-up in a cohort of RA patients, considering the individual clinical laboratory characteristics and treatment.
METHODS
Participants and sample description
Patients included were from the Early Arthritis Clinic (EAC) of the Department of Immunology and Rheumatology, National Institute of Medical Sciences and Nutrition in Mexico City. EAC adult patients (≥18 years of age) are only accepted if they have a disease duration of < 1 year at the first interview. A complete clinical study, including disease history, demographic data, RF, and ACPA results, is registered at admission. A blood sample for DNA extraction is collected at admission from each RA patient, preserved, and used in different studies, including TL analysis. In addition, a follow-up blood sample for DNA extraction was obtained not before 1.5 years of regular surveillance to evaluate telomeres changes after disease exposure time (DET), which was variable, although similar in some RA cases. Furthermore, at admission and at follow-up, we investigated the possible association between the relative leukocyte TL (rLTL) changes and the patient’s clinical characteristics, such as age, DET, HLA-DRB1*04 allele carriage, ESR, CRP, disease activity score (DAS28-ESR)19, RF, ACPA status, and treatments (disease-modifying anti-rheumatic drugs [DMARDs], nonsteroidal anti-inflammatory agents [NSAIDs], and GCCs). The Institutional Research and Ethics Committee approved the study, and signed informed consent was obtained to have the patient’s charts reviewed, presented in scientific forums, and published. The second signed informed consent was requested from the patients and the reference group to obtain their samples for DNA studies. A reference group of healthy individuals, donors of the Department of Transfusion Medicine from the Institute, matched by sex and age to the AAP, was included mainly to evaluate if our sample of patients had a shorter rLTL at the early RA disease stage. Except for the age and sex of healthy subjects, other personal or clinical data were unavailable to preserve confidentiality and the privacy of donors.
DNA samples
DNA from whole leukocytes was obtained with the Wizard® Genomic DNA Purification Kit (Promega, Madison, WI, USA), and the concentrated DNA was preserved at −20°C. DNA quantity and quality were assessed with a Nanodrop 1000 (Thermo Scientific, Wilmington, DE, USA) and agarose gel electrophoresis.
Telomere assay
The rLTL was estimated by monochrome multiplex quantitative polymerase chain reaction (MM-qPCR) developed by Cawthon, with minor modifications20. Primers used for telomere measurements were synthesized by Integrated DNA Technologies, Inc. (IDT, Coralville, IA, USA). We used 7.5 µL of the QuantiTect SYBR® Green PCR ×2 Master Mix (Qiagen, Hilden, Germany), plus 0.3 µL of dNTPs 10 mM (Qiagen, Hilden, Germany), 7.5 ng of gDNA, and RNase-free water for a total of 15 µL. Three reactions were run simultaneously for each sample. In addition, a mix made in our laboratory of five DNA samples from healthy 20-year-old women was included to reference relative TL values. Serial dilutions of the standard DNA sample were also included in each run for PCR efficiency control.
Furthermore, two DNA samples, from a healthy man and woman, were analyzed to control interassay variation in each run. The estimated variation coefficient was 13.68% on 20 assays performed on different days. We used the Rotor-Gene 6000 analyzer (Qiagen, Hilden, Germany) with thermal cycling conditions specified by Cawthon. rLTL was calculated by the 2-ΔΔCt method and expressed as folds of the TL reference sample21. To evaluate the longitudinal rLTL changes in RA patients, we estimated the rLTL relative change as follows: rLTL relative change = ([rLTLFUP–rLTLAAP] ÷ [rLTLAAP]) × 100%.
HLA-DRB*04 allele genotyping
PCR amplification of the HLA-DRB1*04 allele was done as reported22. The amplified products were run in 1.5% agarose gels and the *04 risk allele was documented as a discrete band. Unfortunately, genotyping from two patients failed, limiting the statistical comparisons to 86 individuals instead of 88.
Classification of continuous variables
TL was characterized based on the distribution of rLTL values in a reference group of healthy blood donors. We considered as shorter telomeres, those with an rLTL lower than the 25th percentile (≤ 0.59), and longer telomeres, those with an rLTL over the 75th percentile (≥ 0.92), representing an interquartile of ≥ 33 rLTL units. The rLTL relative change was classified as shortened or unchanged, always considering the qPCR technique’s variation coefficient (13.68%). DET was categorized in percentiles and compared cases with rLTLs under the p25 (4.8 years) with cases over the p75 (9.4 years). We grouped patients by age into young (≤ 40 years) and old (> 40 years), following the previously reported differences in TL observed between RA patients and healthy controls17. Disease activity denoted by the DAS28 was classified into two categories: low to moderate (≤ 5.1) and high (> 5.1)23. The anthropometric variable, body mass index (BMI), was classified as normal weight (< 25), overweight (≥ 25 - < 30), and obesity (≥ 30) following international recommendations.
Statistical analysis
Shapiro–Wilk and skewness/kurtosis tests were used for normality assessment, and the Mann–Whitney and Wilcoxon sign-rank tests were used to compare non-parametric independent quantitative and paired quantitative data. In addition, Pearson’s Chi-square test and Fisher’s exact test were applied for categorical data analysis and the McNemar test for paired data. Finally, Spearman’s correlation analysis assessed the age effect on TL in cases and controls. All statistical analyses were done using Stata Statistical Software: release 12 (StataCorp LP, College Station, TX, USA), and p < 0.05 was considered statistically significant.
RESULTS
Table 1 shows the demographic and clinical characteristics of RA patients at admission and follow-up. All AAP had disease activity, DAS28 over 2.8, with a median of 6.1. At follow-up, 76% of patients decreased to < 2.6, indicating clinical remission. In addition to DAS28, the disease activity-related variables, ESR and CRP, were significantly lower in follow-up patients (FUP) (p < 0.001). The proportion of RF-positive patients had a significant reduction of 14% (p = 0.001) at follow-up, while the percentage of ACPA-positive cases had a non-significant reduction of 4% (p = 0.317). On admission, 81% of patients have treatment regimens without DMARDs (NSAIDs and CCGs). In contrast, DMARDs were part of the treatment regimens in 89% of RA cases at follow-up.
Table 1. Demographic and clinical characteristics of 88 RA patients
| Variables | AAP | FUP | p |
|---|---|---|---|
| Sex, male n (%) | 10 (11.4) | – | |
| HLA-DRB4*04, n (%) | 56 (65.1)a | – | |
| DET, years; median (p25, p75) | 7.3 (4.8, 9.4) | – | |
| Age, years; mean ± SD | 38.7 ± 12.9 | 45.4 ± 13.4 | < 0.0001b |
| BMI; median (p25, p75) | 27 (23, 30) | 28 (24, 31) | 0.0006c |
| Normal weight (< 25), n(%) | 35 (40) | 28 (32) | 0.539d |
| Overweight (≥ 25-< 30) , n(%) | 29 (33) | 32 (36) | – |
| Obesity (≥ 30) , n(%) | 24 (27) | 28 (32) | – |
| Smokers, n (%) | 20 (23) | – | |
| ESR, mm/h; median (p25, p75) | 23 (16, 43) | 8 (3, 18) | < 0.0001c |
| CRP, mg/L; median (p25, p75) | 0.9 (0.4, 3.3) | 0.4 (0.2, 0.9) | < 0.0001c |
| DAS28; median (p25, p75) | 6.1 (5.1, 7.1) | 1.7 (1.2, 2.5) | < 0.0001c |
| Remission (< 2.6), n (%) | 0 | 67 (76) | NA |
| Low activity (2.6-3.2), n (%) | 4 (5) | 4 (5) | NA |
| Moderate activity (3.2-5.1), n (%) | 17 (19) | 14 (16) | NA |
| High activity (> 5.1), n (%) | 67 (76) | 3 (3) | NA |
| RF +, n (%) | 80 (91) | 68 (77) | 0.0013e |
| ACPA+; n (%) | 82 (93) | 78 (89) | 0.3173e |
| DMARDs, n (%) | 17 (19) | 78 (89) | < 0.0001e |
| GCCs, n (%) | 36 (41) | 46 (52) | 0.1228e |
| NSAIDs, n (%) | 72 (82) | 71 (81) | NA |
| Treatment free, n (%) | 10 (11) | 1 (1) | NA |
aGenotype results were available for 86 RA patients only; bPaired t-test; cWilcoxon signed-rank test; dχ2 test; eMcNemar’s χ2. AAP: at-admission patients; FUP: at follow-up patients; DET: disease exposure time; NA: no statistical analysis because of 0 groups, ESR: erythrocyte sedimentation rate; CRP: C-reactive protein; RF: rheumatoid factor; ACPA: anti-citrullinated protein/peptide autoantibodies; BMI: body mass index.
rLTL evaluations in RA patients follow-up
We compared the TL between AAP and FUP. The median rLTL was higher in the first group (0.64; 0.50-0.78) than in FUP (0.61; 0.49-0.70). Paired comparison revealed SSD (p = 0.017; Fig. 1A). It was observed that the telomere shortening was already present in the early stage of the disease, since the median rLTL in AAP was lower than in the reference group (0.77; 0.59-0.92), presenting SSD (p = 0.003; Fig. 1A). Results of rLTL measurements showed a poor inverse statistically significant correlation with chronological age in AAP group (rS = −0.280; 95% CI: −0.491, −0.068; p = 0.008). When patients were stratified by age, those of younger age (≤ 40 years, n = 51) showed a similar median rLTL in AAP (0.66; 0.52-0.87) and at FUP (0.65; 0.52-0.72; p = 0.099) (Fig. 1B). Similar findings were observed in patients over 40 years of age (n = 37) at follow-up (0.55; 0.47-0.66) than at admission (0.57; 0.49-0.71; p = 0.069; Fig. 1C). No statistical difference was found for the time of follow-up between younger (≤ 40 years) and older patients (> 40 years), with median DET of 7.1 years (3.7-9.3) and 7.8 years (4.9-9.5), respectively.
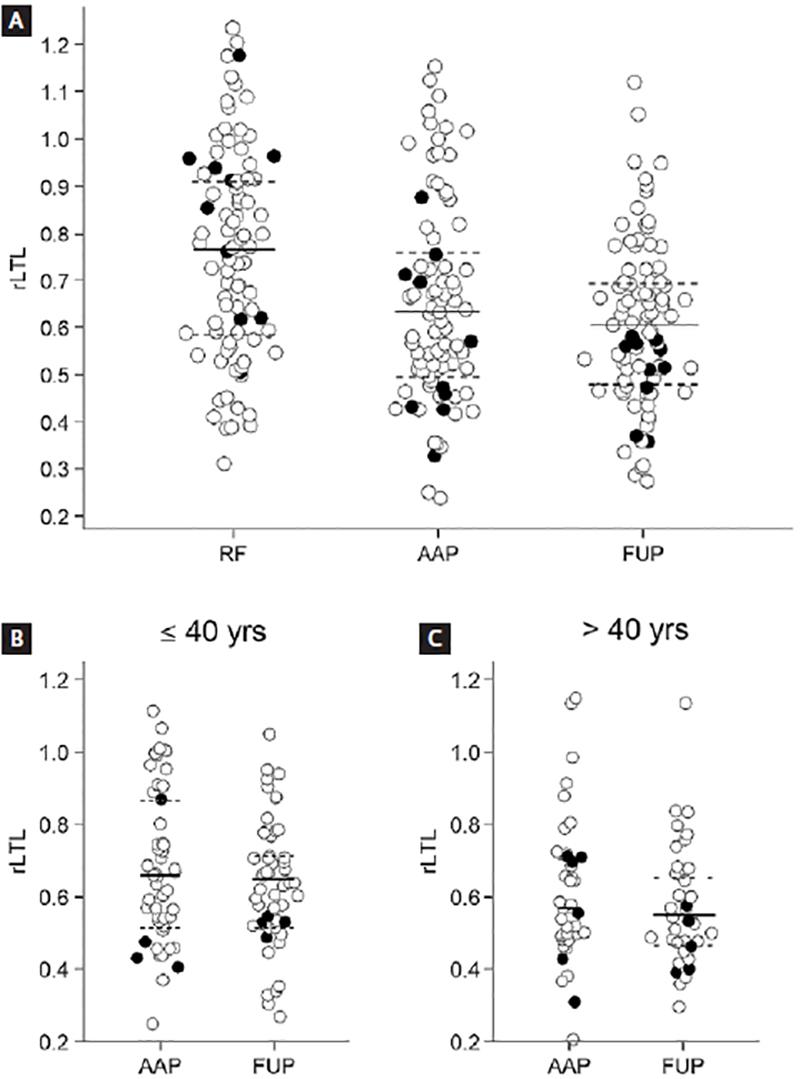
Figure 1. Distribution of relative leukocyte telomere length (rLTL) by comparative groups. (A) rLTL in the reference group (RF), RA patients at-admission (AAP), and at follow-up patients (FUP). (B) rLTL in younger (age ≤ 40 years) AAP and FUP. (C) rLTL in older (age > 40 years) AAP and FUP. Black dots – males, white dots – females, solid lines – median value; dash lines – interquartile range.
We observed that patients with a DET < 4.8 years, 25th percentile (n = 22), had shorter rLTL at follow-up (0.57; 0.51-0.78) than at admission (0.65; 0.54-0.91), showing SSD (p = 0.019). Patients exposed to the disease for ≥ 9.4 years, 75th percentile (n = 21), had at-admission median rLTL (0.58; 0.47-0.72) similar to after follow-up (0.58; 0.46-0.69), with no SSD (p = 0.375). Regarding disease activity variables, the median rLTL of AAP with high disease activity (n=21; 0.62, 0.50-0.82) did not differ significantly from those with low or moderate disease activity (n = 67; 0.64, 0.50-0.75). Moreover, no correlation was observed between rLTL and DAS28 values in AAP (rS = 0.082; p = 0.443) or FUP (rS = 0.022; p = 0.837). Likewise, the AAP rLTL did not correlate with the inflammation biomarker ESR (rS = 0.082; p = 0.443) nor with CRP (rS = −0.009; p = 0.937). We did not observe SSD for rLTL at admission between ACPA-negative (n = 6; 0.53, 0.51-0.56) and ACPA-positive patients (n = 82; 0.65, 0.50-0.82; p = 0.249). On the other hand, comparing median rLTL between AAP and FUP, we observed significantly shorter telomeres at FUP only in ACPA-positive patients (p = 0.002), but not in those ACPA negative (p = 0.207).
Concerning the BMI, the median rLTL was lower in obese AAP (n = 24; 0.54, 0.45-0.69) than in the normal weight AAP (n = 35; 0.66, 0.56-0.89), reaching SSD (p = 0.027). No SSD were found in rLTL between normal weight and overweight AAP (p = 0.487). At follow-up, rLTL was lower in obese FUP (0.53, 0.45-0.67) than in normal weight FUP (0.63, 0.51-0.75), but did not reach SSD (p = 0.099). The smoking status effect on rLTL was considered. The AAP rLTL in a group of smokers (n = 20; 0.61, 0.49-0.82) and never smokers (n = 68; 0.64, 0.51-0.76) did not reach SSD (p = 0.698).
Concerning HLA-DRB1*04 genotype, no rLTL significant changes occurred between AAP (0.59; 0.49-0.73) and at FUP (0.61; 0.49-0.70) in 04* allele carriers (n = 56; z = 1.640; p = 0.101; Fig. 2B). In non-carrier AAP (n = 30), rLTL was higher than in FUP (0.67; 0.54-0.89 vs. 0.62; 0.51-0.79) but not reaching SSD (z = 1.954; p = 0.051; Fig. 2A). At early stage of disease, within the AAP group, HLA-DRB*04 allele carriers (n = 56) showed lower median rLTL value of 0.59 (0.50-0.73) than non-carriers (n = 30; 0.67; 0.54-0.89), although without SSD (z = -1.33, p = 0.183; Fig. 2C). Similarly, when comparing the median rLTL between AAP stratified by age in ≤ 40 years *04 carriers (n = 34; 0.59, 0.52-0.75) and non-carriers (n = 16; 0.70, 0.66-0.99), no SSD was present (z = -1.654; p = 0.098; Fig. 2D). Furthermore, in the group of > 40 years, among *04 carriers (n = 22; 0.62, 0.48-0.71) and *04 non-carriers (n = 14; 0.57, 0.51-0.82), the respective rLTL medians did not significantly differ z = -0.325; p = 0.746; Fig. 2E). HLA-DRB1*04 genotype analysis was also done in the reference group, identifying 37/86 (43.0%) positives, which was significantly lower than in patients (65.1%), indicating a higher genetic predisposition of developing RA being a carrier for the HLA-DRB1*04 allele (OR = 2.47, 95% CI: 1.34-4.57; p = 0.004).

Figure 2. Distribution of relative leukocyte telomere length (rLTL) by HLA-DRB*04 allele carrier status. (A) distribution of rLTL at-admission patients (AAP) and at follow-up patients (FUP) in *04 non-carriers. (B) distribution of rLTL at-admission patients (AAP) and at follow-up patients (FUP) in *04 carriers. (C) distribution of rLTL by HLA-DRB1*04 allele carrier status at the early stage (in AAP). (D) distribution of rLTL by HLA-DRB1*04 allele carrier status in younger (≤ 40 years) AAP. (E) distribution of rLTL by HLA-DRB1*04 allele carrier status in older (> 40 years) AAP. Solid lines – median value; dash lines – interquartile range.
Treatments in the patients were complex combinations of different drugs depending on the individual’s disease activity response. Therefore, to test the treatment effect on rLTLs, we evaluated drug groups individually, DMARDs, NSAIDs, and GCCs. Only the use of GCCs in the AAP group was inversely associated with the frequency of telomere shortening (χ2 = 4.778; p = 0.029); however, when adjusted by at-admission rLTLs, the statistical significance disappeared.
The individual rLTL changes between at-admission and at follow-up cases showed telomere shortening in 34/88 patients (38.6%). However, when analyzed by initial TL categories, shortening after follow-up occurred in 7/41 (17.1%) AAP with shorter telomeres; in 17/34 (50.0%) AAP with average telomeres; and 10/13 (76.9%) AAP with longer telomeres (χ2, 2df = 17.93, p < 0.001), showing those cases with initially longer telomeres a higher frequency of shortening. The clinical and demographic characteristics of patients with shortened and unchanged telomeres are exhibited in table S1. Other variables such as smoking status, RF, ACPA status, and high disease activity (DAS28 > 5.1) did not show any detectable effect on the telomere shortening after follow-up (Table S1). Spearman’s test to assess for a possible correlation of the rLTL changes with clinical and demographic patient characteristics revealed that telomeres’ main shortening is associated with their at-admission TLs (rS = −0.65; p < 0.001), indicating that RA AAP with longer telomeres underwent length reduction in a higher proportion than those with shorter or average telomeres (Fig. 3A). DET had a mild positive correlation (rS = 0.21; p = 0.032; Fig. 3B). No correlation was present between the rate of rLTL changes and the AAP’s age, DAS28 values, ESR, and CRP (Table S2).

DISCUSSION
The present study investigated telomeres length variation and temporal changes in RA patients recruited during the early stage of the disease, < 12 months from symptoms onset, and after a follow-up of different DETs. Significant findings were: (1) premature telomere shortening in patients at the early disease stages; (2) a patient’s initial TL was an influencing factor for telomere shortening; (3) heterogeneous changes in telomere shortening were observed, independently of the follow-up time lag; and (4) neither HLA-DRB1*04 allele carrier status, disease activity markers, nor treatments correlated with TL changes in the present patient cohort.
The current results show shorter peripheral leukocyte telomeres in RA patients at an early stage of the disease than in the healthy sex- and age-matched reference group, probably a sign of premature cell senescence, as confirmed in a recent meta-analysis24. Two different hypotheses were proposed, one, that telomere shortening is the consequence of the inflammation process, characteristic of the disease15, and two, that genetically determined shorter telomeres predispose to RA5. However, the previously reported lack of telomere shortening before RA symptoms onset rejects the hypothesis that short telomeres represent RA risk18. Although patients included in the present study were of different ages at recruitment, they were all within 1 year of symptoms onset. Our results agree with the previous reports, which observed shorter telomeres in RA patients at the initial active disease stage than in age-and sex-matched controls25,26. Telomeric shortening was also previously reported in very young RA patients, 20-30 years17, although these patients were at different disease stages. Our results show shorter telomeres in early-stage RA in younger patients (≤ 40 years) and older patients (> 40 years) without significant differences, not supporting our results with an additive effect between the rLTL in AAP and DET and patient’s age.
The initial disease telomeres shortening in a relatively high proportion of patients could be the effect of disease-related inflammatory events, as observed particularly at follow-up in our sample of ACPA-positive patients, which agrees with the finding that patients at a younger age of disease onset could be more predisposed to severe progressive disease and ACPA positivity, indicating a higher inflammation level27. In addition, obesity contributed to the shorter rLTL in our patients, mainly at admission, with the same tendency at follow-up, according to a previously reported effect of obesity-induced inflammation on TL28.
Our sample of patients of Mexican Mestizo ancestry is characterized by a genetic structure product of the admixture of two main different ethnic groups, indigenous (~50%), Spanish (~45%), and a lesser amount of African ancestry (5%)29. Despite reported distinctive serological, clinical disease features and epidemiological behavior in Latin-American RA patients, such as female preponderance, younger presentation age, and less severe clinical expression than in Caucasians, we observed shorter telomeres length in our Mexican-Mestizo sample of patients than in the reference group, as reported in patients of other ethnic backgrounds30,31.
Telomere shortening in our sample of patients with RA occurred preferentially early during the disease, independently of the age of the patients, which agrees with evident length decay in younger as well as in older patients. On the other hand, the expected age-dependent shortening typically observed during normal aging was not evident in our patients, probably because RA patients had already substantially shortened their telomeres, and because an increase in shortening could induce cell death. Such premature telomere attrition has been reported in peripheral granulocytes, CD4+ T, CD8+ T, and CD34+ hematopoietic precursor cells from RA patients between 20 and 40 years of age5,17,25. However, in patients older than 60 years, the telomeres were similar to controls of similar age32. We observed that RA patients at an early disease stage show shorter telomeres, although they appeared not to be age related.
The present results confirmed the well-established association between HLA-DRB1*04 gene shared epitope in heterozygous and homozygous genotypes and RA. The *04 is the most common HLA-DRB1 allele group in the Mexican population, also reported as one of the shared epitopes containing gene variants frequently found in Mexican RA patients33. In addition, a possible association between the HLA-DRB1*04 allele, RA, and telomere shortening has been reported5. Although the frequency of carriers of the HLA variant was significantly higher than in the reference group, our results showed that rLTL in the HLA-DRB1*04 carriers RA predisposed patients, statistically did not differ from non-carriers independently of age at admission. Moreover, after follow-up, the HLA-DRB1*04 allele carrier status did not affect individual TL. Furthermore, although disease activity modifying drugs, such as methotrexate and chloroquine, frequently used in RA therapy, have been reported to interfere in the mean TL of peripheral leukocytes, this was not observed in our patient cohort.
The present results support that the reported association between short telomeres, RA, and rLTL shortening after follow-up is not associated with the clinical disease activity of the patients, the DET, drug treatment, or HLA-DRB1*04 allele carrier status.
Some limitations of the present study regarding variables that can probably affect telomeres length, including diverse environmental factors, lifestyle, and comorbidities except for smoking status and BMI, were not considered in the analysis in AAP and after follow-up. In addition, no other potentially telomere modifying variables were evaluated.
In conclusion, our observation of significantly shorter rLTL in RA patients at follow-up than at admission, particularly in those AAP with longer telomeres, disregarding disease activity and treatments, denotes an rLTL shortening effect influenced by age, DET, and native telomere rLTL.

