










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkActa zoológica mexicana
versión On-line ISSN 2448-8445versión impresa ISSN 0065-1737
Acta Zool. Mex vol.29 no.3 Xalapa dic. 2013
Artículos originales
Comportamiento espacial de trips (Insecta: Thysanoptera), en plantaciones comerciales de aguacate (Persea americana Mill.) en Zitácuaro, Michoacán, México
Spatial behaviour of thrips (Insecta: Thysanoptera), on avocado commercial orchard (Perseaamericana Mill.), in Zitácuaro, Michoacán, México
José Francisco RAMÍREZ DÁVILA,1 Víctor Manuel SOLARES ALONSO,2 Dulce Karen FIGUEROA FIGUEROA1 & Jesús Ricardo SÁNCHEZ PALE1
1 Laboratorio de Investigaciones Entomológicas y Tecnologías en Agricultura de Precisión; Facultad de Ciencias Agrícolas, UAEM. Cerrillo Piedras Blancas s/n km 15 Carr. Toluca - Ixtlahuaca, entronque al Cerrillo, C.P. 50200; Toluca, Estado de México. <jframirezd@uaemex.mx>
2 Programa de Maestría y Doctorado en Ciencias Agropecuarias y Recursos Naturales. <metavic@hotmail.com>
Recibido: 17/08/2012;
aceptado: 25/04/2013.
RESUMEN
México es el principal productor y exportador de aguacate en el mundo. Los trips son considerados como una de las principales plagas del aguacate en México, debido a su asociación con la enfermedad denominada roña del fruto, causada por el hongo Elsinoe perseae. Esta enfermedad representa una fuerte limitante a la producción del cultivo; el hongo penetra la fruta por las heridas causadas por el complejo de varias especies de trips. El análisis del comportamiento espacial de esta plaga proporcionara información que contribuirá a establecer estrategias de manejo adecuadas y dirigidas dentro de las huertas de aguacate. Este trabajo tuvo por objetivo determinar la distribución espacial de las poblaciones de trips en aguacate (Persea americana Mill) Cv. Hass. mediante el uso de técnicas de estadística espacial (Geoestadística y SADIE) que condujeran en el caso de la Geoestadística a la generación de mapas por medio del krigeado. Se logró determinar además, la estabilidad espacial y temporal a corto plazo de las poblaciones de trips. Los resultados demostraron que las poblaciones de trips presentan una distribución de tipo agregada, ajustándose a los modelos de tipo Esférico y Gaussiano. Dicho comportamiento fue corroborado por los índices de SADIE y los mapas de densidad elaborados. Los resultados demuestran que no existe una infestación del 100% en las parcelas de estudio, lo cual resulta de gran interés para dirigir las medidas de control sobre áreas específicas de infestación y conseguir con ello posibles ahorros económicos y medioambientales.
Palabras clave: Geoestadística, trips, krigeado, SADIE.
ABSTRACT
Mexico is the largest producer and exporter of avocados in the world. Thrips are considered a major pest of avocados in Mexico, because of their association with the disease called fruitscab, caused by the fungus (Elsinoe perseae). This disease represents a serious constraint to crop production; the fungus penetrates the fruit through injuries caused by a complex of several species of thrips. The analysis of the spatial behavior of this pest should provide information that will help establish appropriate management strategies, directed in avocado orchards. This work aimed to determine the spatial distribution of populations of thrips in avocado (Persea americana Mill) Cv. Hass. using spatial statistical (Geostatistics and SADIE) that would lead in the case of Geostatistics to generate maps by kriging. The short term spatial and temporal stability of thrips populations was established.The results showed that thrips populations have an aggregate distribution, adjusting to Spherical and Gaussian type models. Such behavior was corroborated by SADIE indices and the density maps produced. The results demonstrate that there is an infestation of 100% in the study plots, which is of great interest to direct control measures on specific areas of infestation and thereby achieve economic and environmental savings.
Key words: Geostatistics, thrips, kriging, SADIE.
INTRODUCCIÓN
En el 2011 la producción total de aguacate en la República Mexicana alcanzó un poco más de 1 millón 107 mil toneladas. La mayor producción se focaliza en el Estado de Michoacán, lugar donde se producen 950 mil toneladas de la fruta representando el 85% del total (SIAP 2011). Los trips son considerados una de las plagas principales del aguacate; estos insectos se alimentan del fruto en crecimiento (Salgado 1993), causando crestas o protuberancias sobre el pericarpio (González-Hernández et al. 2000). Las deformaciones son más visibles cuando el fruto madura (Fisher & Davenport 1989; González-Hernández et al. 2000). Destacan los géneros Frankliniella y Neohydatothrips; muy probablemente las larvas y adultos son los causantes del deterioro de la calidad comercial de la fruta. Sin embargo, en las especies de Scirtothrips, las larvas y adultos causan daños al follaje tierno, provocando distorsión y cicatrices de color castaño oscuro a lo largo de la nervadura central en el envés de la hoja. Algo similar ocurre con el fruto joven, que se cubre por una cicatriz castaño oscura, aunque comúnmente se observan cicatrices pequeñas, a veces alargadas (Hoddle 2002). Ascensión et al. (1999) afirmaron que este síntoma asociado a trips aparece en el fruto pequeño, aunque no se han precisado las etapas fenológicas más afectadas ni la ubicación espacial dentro de la parcela comercial para realizar un eficiente control. Este trabajo tuvo por objetivo determinar la distribución espacial de las poblaciones de trips en aguacate (Persea americana Mill) Cv. Hass. mediante el uso de técnicas geoestadísticas.
MATERIAL Y MÉTODOS
El estudio se llevó a cabo en el año 2010, en dos parcelas de 4 hectáreas cada una en el municipio de Zitácuaro, dentro de la Región Oriente de Michoacán, todas con antecedentes de trips. Predominando un clima templado con lluvias en verano (Cw), propicio para el cultivo de aguacate (Atlas Geográfico del Estado de Michoacán, 2004).
Cada parcela se dividió en 100 cuadrantes(20 × 20 m), en forma aleatoria se seleccionaron 40 para realizar el muestreo. En cada cuadrante seleccionado se consideraron 3 árboles, para un total de 120 árboles por parcela. Se colocaron 4 trampas por árbol teniendo como referencia cada punto cardinal por árbol. La edad de los árboles de aguacate variedad hass fue de 12 años. Cada árbol se georeferenció utilizando un DGPS (modelo SPS 351, Trimble) para obtener sus coordenadas. Los muestreos se realizaron en forma semanal a partir del mes de febrero hasta noviembre, se contó el número total de ninfas y adultos por árbol. La captura de trips, se logró usando trampas construidas con vasos plásticos No. 8, marca JAGUAR©, de 10 cm de alto, 4.5 cm de diámetro en la base y 7.5 cm de diámetro en el borde superior y de color amarillo canario; de acuerdo con lo sugerido por González et al.(1998) a cada vaso se le colocó un gancho de metal en la base, se les impregno manteca vegetal en la superficie externa, para que los trips quedaran adheridos a la superficie. Las trampas se revisaron semanalmente por 10 meses con una lente de aumento 20X para contar y remover los trips capturados.
Análisis geoestadístico. Se obtuvo el semivariograma experimental a partir del valor promedio mensual de trips capturado en cada muestreo semanal utilizando el programa variowin 2.2 (Software forspatial data analysis en 2D. SpringerVerlag, New York USA). El valor experimental del semivariograma fue calculado con la siguiente expresión (Journel&Huijbregts 1978, Isaaks&Srivastava 1989):
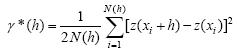
Donde: γ*(h) es el valor experimental del semivariograma para el intervalo de distancia h; n (h) es el número de pares de puntos muestrales separados por el intervalo de distancia h; z (xi) es el valor de la variable de interés en el punto muestral xi y z (xi+h) es valor de la variable de interés en el punto muestra x.+h.
Estimación de los parámetros del modelo de semivariograma. Una vez obtenido el semivariograma experimental, se realizó su ajuste a algún semivariograma teórico (esférico, exponencial, gaussiano, etc.) (Englund&Sparks 1988) utilizando el programa Variowin versión 2.2.
Validación del modelo teórico. Se sigue un procedimiento de validación cruzada, en el cual los parámetros del modelo Co (efecto de pepita), C + Co (meseta) y a (rango o alcance) se van ajustando de forma interactiva (prueba y error) hasta los mejores valores de los estadísticos:
a. Media de los errores de estimación (MEE):
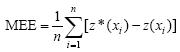
Donde: z*(xi) es el valor estimado de la variable de interés en el punto xi; z (xi) es el valor medido de la variable de interés en el punto xi y n es el número de puntos muéstrales utilizado en la interpolación. La MEE no debe ser significativamente distinta de 0 (Prueba de t), en cuyo caso, indicaría que el modelo de semivariograma permite el cálculo de estimadores no sesgados.
b. Error cuadrático medio (ECM):

Un modelo de semivariograma se considera adecuado si, como regla práctica, el valor del estadístico es cercano a cero (Hevesiet al. 1992).
c. Error cuadrático medio adimensional (ECMA):

Donde: k es la desviación estandar del error esperado en la estimación con el krigeado. La validez del modelo se satisface si ECMA está comprendido entre los valores 12 (2/N)0.5.
d. Otro estadístico para validar el ajuste del modelo consiste en que el valor de la varianza de los errores sea menor a la varianza muestral.
Nivel de dependencia espacial. La determinación del grado de relación entre los datos o nivel de dependencia espacial se obtuvo al dividir el efecto pepita entre la meseta y expresando en porcentaje su resultado. Si el resultado es menor de 25% el nivel de dependencia espacial es alta, si se encuentra entre 26 y 75% el nivel de dependencia espacial es moderado y si es mayor del 76% el nivel de dependencia es bajo (Cambardellaet al. 1994).
Elaboración de mapas. Se realizó la interpolación de valores a través del krigeado ordinario que permite la estimación insesgada de valores asociados a puntos que no fueron muestreados, las estimaciones obtenidas fueron representadas en forma de mapa para cada fecha de muestreo mediante el uso del programa Surfer 9 (Surface-MappingSystem, Golden Software Inc. 809, 14th Street. Golden, Colorado 804011866. USA).
SADIE (Spatial Analysis by Distance Indices). Dentro de los nuevos desarrollos de la estadística espacial ha aparecido recientemente una nueva metodología llamada SADIE (Análisis Espacial por Índices de Distancia) que ha sido desarrollada por el Dr. Joe N. Perry del Dpto. de Entomología y Nematologia de Rothamsted Experimental Station (Reino Unido). SADIE identifica el modelo espacial para datos bidimensionales, con un índice asociado de la agregación y de una prueba para la desviación de la aleatoriedad basada en un algoritmo de atracción, el cual incorpora un modelo biológico para la dispersión de individuos de un origen en el que a cada individuo se le asigna un territorio dinámico. Es un índice biológicamente más descriptivo e informativo de la distribución espacial de las poblaciones de insectos que el índice de dispersión (cociente media varianza) y el Índice de Green, los cuales dependen directamente de la abundancia de la población (Perry et al. 1996). Perry (1995a) indicó que para datos recolectados en ubicaciones específicas el uso de la distancia para la regularidad es muy adecuado y demostró cómo distinguir no aleatoriedad en la forma de heterogeneidad estadística, de la no aleatoriedad espacial. Perry (1995b) desarrolló y extendió el uso del índice de la distancia para la regularidad (Ia) para el establecimiento de la estructura espacial de las poblaciones de insectos. Además, introdujo dos diagramas de diagnóstico como ayuda a la interpretación y un índice nuevo para estimar el número de focos de agrupamiento de una población, el índice Ja.
Estabilidad espacial y temporal. La comparación se realizó utilizando la modificación propuesta por Syrjala (1996) a la prueba estadística no paramétrica de Cramérvon Mises, cuando los resultados se obtuvieron con la prueba de krigeado; mientras que para el caso de los resultados obtenidos mediante el método SADIE, la comparación para establecer la estabilidad espacio temporal de los mismos se llevó a cabo mediante el índice de asociación del SADIE (Im) (Perry &Klukowsky 1997) considerando que si Im> 0, es indicativo que existe una asociación o una estabilidad espacio-temporal entre los mapas. La determinación de la estabilidad se realizó con el programa SADIE 1.22.
RESULTADOS
Con los datos obtenidos de los muestreos fue posible generar la modelización espacial y el mapeo de las poblaciones de trips en forma mensual dentro de la parcela comercial de aguacate. Se estableció el porcentaje estimado de infestación en cada fecha de muestreo y se logró determinar la estabilidad espacio-temporal de este insecto a corto plazo. También se pudo establecer que los géneros de trips presentes en las plantaciones de aguacate son Frankliniella y Scirtothrips, y que las especies más abundantes fueron F. fallaciosa Priesner y S. perseae Nakahara respectivamente.
La media de la población de trips en la parcela 1 vario de 2.983 individuos (septiembre) a 140. 258 individuos (junio). En la parcela 2 la densidad media fluctuó entre 17.525 individuos (septiembre) y 98. 991 individuos (mayo). Las densidades más bajas se presentaron en septiembre y las más altas entre mayo y junio, lo anterior relacionado con el inicio del aumento de la temperatura (Tabla 1).
La distribución espacial obtenida para las poblaciones de trips fue de tipo agregada para cada una de las fechas de muestreo en las dos plantaciones de aguacate cultivar hass (Tabla 1).
En la parcela 1, los semivariogramas experimentales se ajustaron al modelo gaussiano, excepto para los meses de septiembre y octubre cuando se ajustaron al modelo esférico (Tabla 1). Los semivariogramas de la parcela 2, se ajustaron al modelo esférico en los meses de febrero, marzo, septiembre, octubre y noviembre; las restantes fechas se ajustaron al modelo gaussiano (Tabla 1). Para todos los modelos ajustados se presentó un efecto pepita igual a cero, dicho valor es indicativo de que la escala de muestreo utilizada en el presente trabajo fue la adecuada y el error de muestreo fue mínimo. Los valores del rango, en la parcela 1, se ubicaron entre 9.596 y 41.908 metros, mientras que para la segunda parcela sus valores fluctuaron entre 18.392 y 35.288 metros. Siendo el rango la distancia máxima hasta la cual existe relación espacial entre los datos. En todos los modelos se presentó un alto nivel de dependencia espacial para cada una de las fechas de muestreo (Tabla 1). Los modelos de la distribución espacial de trips en las dos parcelas analizadas se lograron validar con los parámetros estadísticos al ubicarse dentro del rango permisible (Tabla 2).
Los mapas de la superficie infestada en la parcela 1, mostraron que las poblaciones de trips se ubicaron en centros de agregación, es decir, su distribución se localiza en puntos específicos en la región central durante los meses de febrero a junio, siendo este último donde se evidencia una mayor intensidad; para los meses de julio y agosto los centros de agregación se ubican en la parte inferior derecha y superior izquierda respectivamente. Para los meses de septiembre a noviembre los centros de agregación se localizan en la parte central del mapa, siendo septiembre el que se presenta de forma irregular (Fig. 1).
Sin embargo, los mapas generados en la parcela 2 mostraron que para los meses de febrero y marzo los centros de agregación presentan una tendencia de ubicarse en la región inferior izquierda y hacia la región inferior derecha, abril y mayo muestran una tendencia central y parte inferior derecha respectivamente. Los mapas de junio a septiembre presentan una ubicación irregular, lo mismo sucede con el mes de noviembre, por último el mes de octubre se ubican en la región central, con una ligera tendencia a la parte inferior (Fig. 2).
En la parcela 1, la menor superficie infestada estimada (63%), se presentó durante el mes de septiembre (Tabla 3) con una media de 2.983 trips por árbol (Tabla 1). Mientras que la mayor superficie infestada (90%) se presentó en el mes de agosto (Tabla 3) con una densidad media de 41.558 ejemplares por árbol (Tabla 1). Por otro lado en la parcela 2, la menor superficie infestada estimada (67%) se presentó en el mes de septiembre (Tabla 3) con una media de 17.525 ejemplares por árbol (Tabla 1), mientras que la mayor superficie infestada (90%) se presentó durante el mes de octubre (Tabla 3) con una densidad de 27.891 ejemplares por árbol (Tabla 1).

Los valores obtenidos con el índice de SADIE se indican en la Tabla 4. En todos los casos el índice Ia fue significativamente superior a 1, lo cual señala que la distribución de trips es de tipo agregada, en la parcela 1 y 2. Resultados similares se encontraron con el índice Ja, su valor no fue significativamente superior a 1, lo que permite determinar que la distribución espacial de las poblaciones del insecto se concentró en diferentes centros de agregación en las dos parcelas estudiadas.
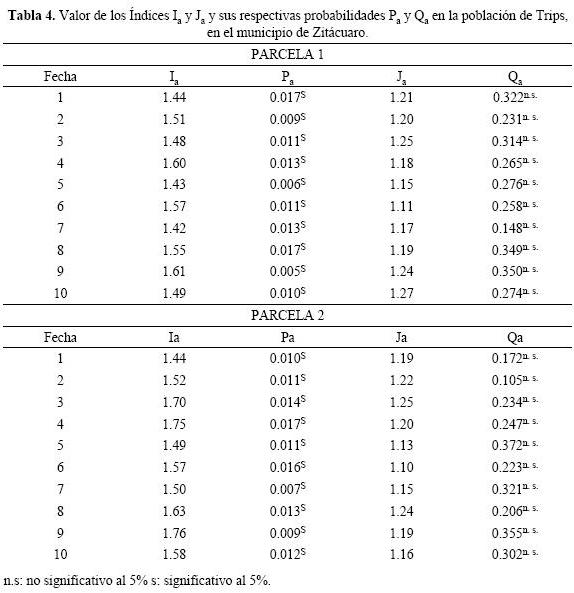
La distribución agregada de las poblaciones (Figuras 1 y 2) del insecto se manifiestan en cada uno de los mapas obtenidos, lo que corrobora lo establecido por los índices Ia y Ja. En los mapas realizados se alcanzan a apreciar los diferentes focos de agregación de las poblaciones, en las diez fechas, para las dos parcelas. Los numerosos focos de agregación detectados en cada fecha de muestreo, corroboran lo detectado por el índice Ja en las diez fechas de muestreo.
Estabilidad espacial y temporal. En la parcela 1 (Tabla 5), se indican los resultados de la comparación entre los mapas de la distribución espacial de las poblaciones de trips para cada fecha de muestreo, los valores Im del SADIE fueron mayores que 0, indicando que no existió diferencia significativa, es decir se detectó asociación espacial entre los mapas comparados para trips, por lo que fue posible detectar una estabilidad espacio temporal a corto plazo de las poblaciones del insecto entre las fechas comparadas; excepto para las comparaciones 5 vs 6, 7 vs 8 y 8 vs 9, los valores fueron menores que 0, esto indica que no existió asociación espacial. Para la parcela 2, todas las fechas comparadas mostraron una asociación espacial entre los mapas (Tabla 5).

Los resultados obtenidos con la prueba estadística bivariable de Cramér-von Mises, indican diferencia no significativa en la estabilidad espacial y temporal de corto plazo al comparar la mayoría de las fechas de muestreo (Tabla 5), para las dos parcelas analizadas, es decir, existió estabilidad espacial y temporal en las poblaciones de trips entre las fechas analizadas. Dicho de otra manera, los mapas presentan similitud estadística entre la dispersión de las poblaciones del insecto. Cabe resaltar que al comparar fechas 5 vs 6, 7 vs 8 y 8 vs 9 de la parcela 1 (Tabla 5) si existió diferencia en ese patrón de comportamiento, reflejándose una diferencia significativa entre los mapas, lo cual muestra que no hubo estabilidad espacial y temporal en esos casos.
DISCUSIÓN
La mayor densidad poblacional de trips en huertos de aguacate se presentó en los meses de julio a septiembre, descubrimiento que contrasta con lo determinado por González et al. (1998) quienes encontraron que en Michoacán, las poblaciones de trips presentaron picos entre marzo y mayo, mientras que durante el periodo de lluvias (junio-septiembre) la población bajo notablemente, para posteriormente tener un nuevo aumento a partir de octubre.
La determinación de una estructura agregada en la modelización de la distribución espacial de trips en aguacate se logró con el uso de la geoestadística. Los métodos geoestadísticos proporcionan una medida más directa de la dependencia espacial, ya que tienen en cuenta la naturaleza bidimensional de la distribución de los organismos a través de su exacta localización espacial y es independiente de la relación entre la media y la varianza. La Geoestadística además, permite elaborar mapas de gran utilidad de la distribución espacial de un organismo (Rossi et al. 1992). El uso de dicha técnica para modelizar la distribución espacial de insectos, ha sido reportado en diferentes trabajos tal como el de Sciarretta et al. (2008) quienes determinaron la distribución espacial de Lobesia botrana en predios de uva y el de Bressanet al. (2010) con Pentastiridius leporinus L., en campos de trigo.
Al respecto de la estructura espacial Weiszet al., (1996a) mencionan que debido a la movilidad innata de los insectos adultos y de sus estadíos preimaginales, la estructura espacial hallada debe de considerarse "dinámica". Si bien compartimos lo anterior plenamente, debemos señalar que las características del insecto pueden matizar el "dinamismo" de la estructura espacial.
El comportamiento de tipo agregado en las poblaciones de trips en aguacate permite sugerir que la reducción de las infestaciones se puede lograr al focalizar las estrategias de manejo hacia puntos específicos o focos de infestación en donde se ubican los puntos de agregación, además de poder visualizarlos a través de los mapas generados, concordando con lo propuesto por Rong et al. (2006) quienes propusieron realizar el manejo de Locusta migratoria manilensis (Meyen) en puntos específicos. Resultados similares fueron determinados con los índices Ia y Ja de SADIE, los cuales indicaron la estructura espacial agregada de las poblaciones de trips. Además el índice Ja señaló la existencia de varios centros de agregación, tal como se visualiza en los mapas obtenidos. Dichos resultados concuerdan con la distribución espacial determinado en otros organismos por Conrad et al. (2006) y Ramírez & Porcayo (2009).
Los valores de cero en el efecto pepita de los diferentes modelos ajustados, permiten afirmar que la escala de muestreo fue correcta, y que el error de muestreo fue mínimo (Oliver & Webster 1991) por lo que los modelos ajustados tienen un 98% de fiabilidad, lo anterior da pie a deducir más del 90% de la variación total era debida a la dependencia espacial encontrada en la escala de muestreo utilizada, dicho de otra manera, más del 90% de la variación de la distribución de las poblaciones del insecto se logró explicar por la estructura espacial establecida con los semivariogramas (Liebhold & Sharov 1998). Además, las funciones estadísticas permitieron validar los modelos del tipo esférico y gaussiano en ambas parcelas en las diferentes fechas de muestreo.
El ajuste de las poblaciones de trips al modelo gaussiano, en la parcela 1 para la mayoría de los casos, es indicativo de que el comportamiento espacial de la agregación se expresa en forma continua dentro de la plantación de aguacate. Lo que implica un continuo avance de las infestaciones de trips a arboles de aguacate vecinos, esto corrobora además, la alta preferencia de ataque de este insecto sobre la variedad hass. La existencia de condiciones ambientales similares en toda la extensión de la parcela no resulta una limitante para la distribución del insecto plaga.
En la parcela 2, el ajuste de la distribución espacial al modelo esférico, durante los meses de febrero, marzo y septiembre a noviembre, indica que las agregaciones del insecto se presentan en mayor cantidad en ciertas zonas de la parcela respecto al resto de puntos considerados en el muestreo. Es decir, los centros de agregación son aleatorios dentro de la zona de infestación de la parcela, posiblemente como resultado de la diseminación del insecto a través del viento lo que origina infestaciones en zonas específicas, dichas zonas se observan en los mapas obtenidos. La agregación ajustada al modelo gaussiano en las fechas de abril hasta agosto, nos muestra una manifestación continua en su distribución, avanzando hacia otras partes de la parcela de aguacate, a pesar de que nunca alcanzó una infestación del 100%.
El alto nivel de dependencia espacial resultó de dividir el valor del efecto pepita entre el valor de la meseta que fue menor a 25% para todos los semivariogramas. Los valores del efecto pepita indicaron una alta dependencia espacial, lo cual permite suponer que las poblaciones de trips dependen entre si y su nivel de agregación es alto (Rossi et al. 1992).
En los mapas de densidad obtenidos con la técnica de krigeado se observan los centros de agregación de las poblaciones de trips, que desde el punto de vista de protección de cultivos permite dirigir de forma precisa las diferentes medidas de control del insecto (Fleischer et al. 1997; Rong et al. 2006; Weisz et al. 1996) generando beneficios económicos al realizar con menor frecuencia las aplicaciones de plaguicidas, menor uso de combustible, menor impacto al medio ambiente, retardar el desarrollo de resistencia del insecto debido a la creación de refugios temporales dinámicos, al no tratar toda la superficie del cultivo (Fleischer et al. 1999a), además nos permite visualizar la variabilidad poblacional e identificar áreas infestadas y libres de infestación. Dicho mapas pueden conducir al manejo con precisión de la plaga, al dirigir las medidas de control hacia las áreas específicas de infestación (Fleischer et al. 1999b). El manejo de focos de infestación permitirá obtener frutos de aguacate con una mayor inocuidad, tal como lo solicitan los mercados de exportación, según la Asociación de Empacadores y Exportadores de Aguacate del Estado de Michoacán (ASEEAM 1998). La mejora de la sanidad permitirá una mayor productividad del cultivo de aguacate para el beneficio de los productores. Los mapas permitieron identificar áreas infestadas y libres de infestación, tal apreciación permite indicar que el trips no invade el 100% de la superficie de las parcelas analizadas en la región de Zitácuaro, Michoacán. La determinación de áreas libres de infestación de trips dentro de las parcelas estudiadas concuerdan con lo reportado por Ramírez & Porcayo (2008) quienes obtuvieron mapas con áreas libres de infestación de Jacobiasca lybica en viñedos. La estabilidad espacio temporal encontrada en el presente estudio es de gran utilidad ya que nos permite conocer la permanencia del insecto en puntos específicos dentro de la parcela, en donde se ubican sus nichos o sitios de reproducción a través de tiempo. Dicha estabilidad nos permite realizar acciones de control de tipo preventivas y focalizadas, para mantener bajos niveles de infestación. La estabilidad a corto plazo encontrada puede deberse a el hecho de la poca movilidad de los trips, lo cual nos puede indicar que no existen nuevos centros de agregación entre las comparaciones, o bien, no son significativas, también nos muestra que las actividades que realiza el productor en la parcela no tuvieron un efecto en la reducción de la población o centro de agregación entre una fecha y otra, o bien, no lograron romper tal ciclo de reproducción del insecto. Finalmente, otra posible causa que origina la estabilidad espacial es la permanente presencia de maleza, dentro de las parcelas de aguacate, que fungen como reservorio de los trips, tal como lo menciona Jiménez (1987) quien recomienda el mantener los huertos libres de maleza, ya que sirven de refugio o como hospederas alternantes de los trips. El barbechar el suelo después de la cosecha y antes de la próxima floración, principalmente debajo del dosel del árbol, puede contribuir a eliminar las pupas de los trips, ya que se exponen al efecto del sol, el viento o las altas temperaturas, lo que ocasiona su desecación y muerte.
AGRADECIMIENTOS
Al apoyo del Consejo Nacional de Ciencia y Tecnología, al Ing. Juan X. Mora y a los productores de aguacate en Michoacán: Comunidad de Zitácuaro, Jorge Arredondo y Homero Carmona.
LITERATURA CITADA
Ascensión-Betanzos, G., Bravo, M. H., González, H. H., Johansen, R.& Becerril, R. E. 1999. Fluctuación poblacional y daño por trips en aguacate cv. Hass. IV Congreso Mundial del Aguacate. Revista Chapingo serie Horticultura, 5: 291-296. [ Links ]
ASEEAM, 1998. El aguacate mexicano: producción y destino. Uruapan, Michoacán. 43 pp. [ Links ]
Atlas Geográfico del Estado de Michoacán. Secretaría de Educación en el Estado de Michoacán, Universidad Michoacana de San Nicolás de Hidalgo. 2004. EDDISA. 143-150. [ Links ]
Bressan, A., Moral, J. F., Sémétey, O. & Boudon-Padieu, E. 2010.Spatio-temporal pattern of Pentastiridiusleporinus migration in an ephemeral cropping system. Agricultural and Forest Entomology, 12: 59-68. [ Links ]
Cambardella, C., Moorman, T., Novak, J., Parkin, T., Karlen, D., Turco, R. & Konopka, A. 1994. Field scale variability of soil properties in central Iowa soils. Soil Science Society of American Journal, 58: 1501-1511. [ Links ]
Conrad, K. F., Perry, J. N., Woiwod,I. P. & Alexander, C. J. 2006. Large scale temporal changes in spatial pattern during declines of abundance an occupancy in common moth. Journal of Insect Conservation, 10: 53-64. [ Links ]
Englund, E. & Sparks, A. 1988.GEO-EAS (Geostatistical Environmental Assessment Software) User's Guide. U.S. Environmental Protection Agency. Document EPA/600/4-88/033. Environmental Monitoring Systems Laboratory, Las Vegas, NV, USA, 186 pp. [ Links ]
Fischer, J. B. & Davenport, T. L. 1989. Structure and development of surface deformation on avocado fruits. HortScience, 24: 841-844. [ Links ]
Fleischer, J., Blom, E. P. & Weisz, R. 1999a. Sampling in Precision IPM: When the objetive is a map. Phytopathology, 89: 115-118. [ Links ]
Fleischer, J., Blom, E. P., Emmen, D. & Hower, A. 1999b.Dynamics in the spatial continuity of insect density. Fourth International Conference on Presicion Agriculture,pp. 133- 141. [ Links ]
Fleischer, S., Weisz, R., Smilowitz,Z. & Midgarden, D. 1997.Spatial variation in insect populations and site-specific integrated pest management, pp. 101-130. In: Pierce, F. J. & Sadlerb, E. J. (Eds.).The state of Site-Specific Management for Agriculture, ASA miscellaneous Publication.Madison, WI, 430 pp. [ Links ]
González H., H., Sosa T., C.M., González R., M. & Valle de la Paz, A.R. 1998. Selección del color de trampas adhesivas para muestreo de trips en el cultivo del aguacate (Persea americana Mill.), pp. 209-210. In: Avances de investigación, 1997. Instituto de Fitosanidad, Colegio de Postgraduados. Montecillo, México. [ Links ]
González Hernández, H., Johansen, R., Gasca, L., Equihua, A., Salinas, A., Estrada, E., Duran, F. & Valle, A. 2000. Plagas del aguacate. In: Téliz., D. (Cd.). El Aguacate y su Manejo Integrado. Ediciones Mundi Prensa. México, DF. pp: 177-186. [ Links ]
Hevesi, J., Istok, J. & Flint, A.1992. Precipitation estimation in mountainous terrain using multivariate geostatistics. Part I. Structural analysis. Journal of Applied Meteorology, 31: 661-676. [ Links ]
Hoddle, M. S. 2002. Development and reproductive biology of Scirthothripsperseae (Thysanoptera: Thripidae). Review avocado post in California, Bulletin Entomology Research, 92: 279-285. [ Links ]
Isaaks E. H. & Srivastava, R. M.1989. An Introduction to applied Geostatistics. Oxford University Press. New York. 561 pp. [ Links ]
Jiménez, R. P. 1987. Plagas y enfermedades que limitan la producción del aguacate y su control en el estado de Michoacán. UCAIM. Revista No. 10. Uruapan, Mich. México. [ Links ]
Journel, A. G. & Huijbregts, C. J. 1978. Mining Geostatistics. Academic Press, London, U.K. 600 pp. [ Links ]
Liebhold, A. & Sharov,A.1998. Testing for correlation in the presence of spatial autocorrelation in insect count data, p. 1-17. In: Baumgartner, J., Brandmayr, P. & Manly, B. F. J. (Eds.). Population and community Ecology for insect management and conservation. Balkema, Rotterdam, 435 pp. [ Links ]
Oliver, M. & Webster, R. 1991. How geostatistics can help you. Soil use and Management, 7: 206-217. [ Links ]
Perry, J. 1995a. Spatial aspects of animal and plant distribution in patchy farmland habitats, pp. 221– 242. In: Glen,D.M., Greaves,M.A. & Anderson, H.M. (Eds.). Ecology and Integrated Farming Systems, Wiley, Chichester, England. [ Links ]
Perry, J. 1995b. Spatial Analysis by distance indices. Journal of Animal Ecology, 64: 303-314. [ Links ]
Perry, J. N. & Hewitt, M. 1991. A new index of aggregation for animal counts. Biometrics, 47: 1505– 1518. [ Links ]
Perry, J., Bell, E., Smith,R. & Woiwod, I. 1996. SADIE. Software to measure and model spatial pattern. Aspects of Applied Biology, 46: 95-102. [ Links ]
Perry, N. &Klukowsky, Z. 1997. Spatial distributions of counts at the edges of sample areas, pp. 103-108. In: VI Conferencia de la sociedad de biometría. Córdoba, España. [ Links ]
Ramírez, J. F. & Porcayo, E. 2008. Distribución espacial de las ninfas de Jacobiasca líbica (Hemiptera: Cicadellidae) en un viñedo en Andalucía, España. Revista Colombiana de Entomología,34: 169–175. [ Links ]
Ramírez, J. F. & Porcayo, E. 2009. Estudios de la distribución espacial del muérdago enano (Arceuthobimsp.) en el Nevado de Toluca, México, utilizando el Método de SADIE. Madera y Bosques, 34: 93–111. [ Links ]
Rong, J., Dian-Mo, L., Bao-Yu, X., Zhe,L. & Dong-Li,M.2006. Spatial Distribution of Oriental Migratory Locust (Orthoptera: Acrididae) Egg Pod Populations: Implications for Site-specific Pest Management. Environmental Entomology, 35: 1244-1248. [ Links ]
Rossi, R., Mulla, J., Journel,G.& Franz, H. 1992.Geostatical Tools for Modeling and interpreting Ecological Spatial Dependence. EcologicalMonographs, 62: 277-314. [ Links ]
SIAP.SAGARPA. 2011. Servicio de Información Agroalimentaria y Pesquera. http://www.siap.gob.mx/ (Consulta: 7 de febrero 2011). [ Links ]
Salgado S., M. L. 1993. Problemas fitosanitarios del aguacate en Coatepec Harinas. In: Memorias Centro de Investigaciones científicas y tecnológicas del aguacate en el Estado de México. pp. 191-212. [ Links ]
Samper, F. J. & Carrera, J. 1996. Geoestadística: Aplicaciones a la Hidrología subterránea. 2nd ed. Centro Internacional de Métodos en Ingeniería, Barcelona, España, 484 p. [ Links ]
Sciarretta, A. & Trematerra, P. 2006. Geostatistical characterization of the spatial distribution of Grapholitamolesta and Anarsiaineatella males in an agricultural landscape. Journal of Applied Entomology, 130: 73–83. [ Links ]
Sciarretta, A., Zinni, A., Mazzocchetti, A. & Trematerra, P. 2008. Spatial Analysis of Lobesiabotrana (Lepidoptera: Tortricidae) Male Population in a Mediterranean Agricultural Landscape in Central Italy. Environmental Entomology, 37: 382-390. [ Links ]
Syrjala, E. 1996. A statical test for a difference between the spatial distributions of two population. Ecology, 77: 75-80. [ Links ]
Weisz, R., Fleischer,S. & Smilowitz, Z. 1996. Site-specific integrated pest management for high value crops: Sample units for map generation using the Colorado potato beetle (Coleoptera: Chrysomelidae) as a model system. Journal Economic Entomology, 88: 1069-1080. [ Links ]

