











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink¿Qué es el endemismo?
La importancia del endemismo radica en la necesidad de conocer y proteger los atributos biológicos e historia evolutiva que representan los taxones endémicos y sus patrones biogeográficos. El endemismo toma un valor ecológico e histórico, si se considera que define zonas geográficas donde hay taxones integrados espacio-temporalmente (Nelson & Platnick 1981; Stattersfield et al. 1998; Morrone 2009).
La palabra “endémico” proviene del griego “endēmios = nativo”. Fue transferido por Candolle (1820) desde la medicina a la botánica para indicar taxones nativos que se distribuyen en un lugar y no en otro (Anderson 1994; Hobohm & Tucker 2014). Aunque los variados usos y aplicaciones del endemismo han ocasionado confusiones en su interpretación (Anderson 1994), las definiciones coinciden en que el “endemismo” es una implicación de restricción geográfica.
Es evidente que para aplicar el término “endemismo” se necesita relacionar un área geográfica de restricción, ya que el endemismo es dependiente de la escala geográfica del área en la que se distribuya el taxón (Cracraft 1985). Por ejemplo, el área de distribución del jaguar (Panthera onca [Linnaeus 1758]) se extiende desde México hasta el norte de Argentina (Caso et al. 2008), por lo tanto esta especie se puede considerar endémica del Neotrópico (Fig. 1). En otro caso el mapache pigmeo (Procyon pygmaeus Merriam 1901), restringido a la Isla de Cozumel en México, es endémico para esa isla (Cuarón et al. 2008) (Fig. 1). En comparación con el jaguar, el mapache pigmeo presenta una distribución reducida a sólo una parte del Neotrópico. Por lo tanto, cada una de las especies es endémica dentro del área geográfica con la que se relaciona su distribución, Neotrópico e Isla de Cozumel, respectivamente.
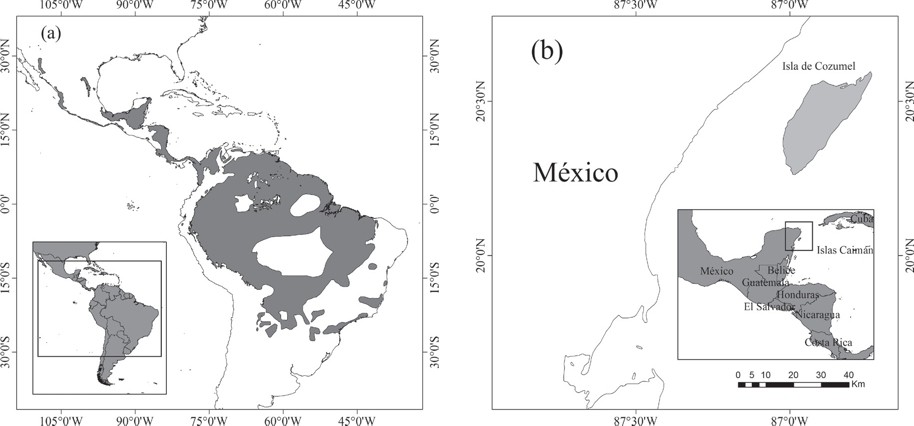
Figura 1 Endemismo a diferentes escalas y diferentes límites geográficos. (a) Distribución geográfica del jaguar (Panthera onca) y (b) mapache pigmeo (Procyon pygmaeus). Cada una de las especies es endémica al área geográfica de referencia, el jaguar al Neotrópico y el mapache pigmeo a la Isla de Cozumel. En cada área geográfica que se tome como referencia hay uno o más taxones que pueden ser considerados endémicos.
Enfoques en la aplicación del concepto
Los múltiples usos y aplicaciones del “endemismo” tienen como base la definición de Candolle (1820), al tomar a la unicidad geográfica para cualificar a un taxón como endémico. Las variantes del concepto son propuestas para la formalización de la relación entre la evolución de un taxón y su área de distribución, o la necesidad de un criterio práctico para definir objetos y/o áreas prioritarias de conservación. Por lo tanto, mantener el sentido que le dio Candolle (1820) al endemismo debería primar en las aplicaciones biogeográficas para todo tipo de organismos. Y cuando sea posible, se deberían emplear unidades geográficas naturales (ecosistemas, cuencas, provincias biogeográficas, regiones biogeográficas, etc.) en los análisis relacionados con el endemismo. Con ello se lograría usar un mismo contexto teórico del endemismo y se aclararía la combinación del “endemismo” con otros términos.
En algunos casos el endemismo ha sido interpretado con base en la rareza de poblaciones o comunidades (Major 1988), esto debido al conocimiento incompleto de las áreas de distribución (Kruckeberg & Rabinowitz 1985). La aplicación del concepto ha cambiado para ser aplicado a taxones (especies, géneros, familias, etc.), debido al progresivo aumento de los sitios de colecta y el desarrollo de hipótesis sobre áreas de distribución. Desde hace unos años el endemismo se relaciona con taxones restringidos a una sola área geográfica o que presentan área de distribución reducida (Polunin 1960; Anderson 1994; Peterson & Watson 1998). Pese a ello, una de las críticas en los procedimientos para la definición de los taxones endémicos fue la ausencia de un criterio práctico (Stattersfield et al. 1998; Bruchmann & Hobohm 2014). En este contexto, se han propuesto múltiples enfoques para determinar el criterio de restricción y/o relacionar el endemismo con la evolución de los taxones.
El endemismo en ecología y conservación ha sido usado como distribución restringida (restricted-range) y para definir a las especies endémicas se ha empleado un tamaño de área geográfica como límite práctico (Stattersfield et al. 1998). Por ejemplo, una especie con un área de distribución menor a 100,584 km2 (250 millas en cualquier dirección) (Hall & Moreau 1962); 50,000 km2 (área de distribución sin aumentos desde el año 1,800) (Terborgh & Winter 1983; Stattersfield et al. 1998) o 10,000 km2 (Kier & Barthlott 2001; Bruchmann & Hobohm 2014) tendrá una distribución restringida (Terborgh & Winter 1983; Stattersfield et al. 1998) (Fig. 2). De la aplicación del endemismo bajo el criterio de distribución restringida resultan, por ejemplo, lugares de conservación conocidos como Áreas de Aves Endémicas (Endemic Birds Areas; EBA). Las Áreas de Aves Endémicas son zonas en las cuales dos o más especies cohabitan y presentan áreas de distribución menores a 50,000 km2 (Stattersfield et al. 1998). Por otra parte la flexibilidad del criterio de restricción ha llevado a que los límites administrativos de un país u otras áreas puedan ser usados para definir taxones endémicos (Ceballos et al. 1998; González-García & Gómez de Silva 2003). En el caso de plantas mexicanas, se ha propuesto que especies con áreas de distribución menores, similares o levemente mayores a los límites geopolíticos de México, sean consideradas endémicas (Rzedowski 1991). El endemismo basado en restricción a un país toma un gran valor para los análisis de riqueza de especies y su conservación. De ello resultan otras interpretaciones para definir especies endémicas. Por ejemplo, en México se consideran 157 especies de mamíferos como endémicas (28%, total: 564 especies) (Sánchez-Cordero et al. 2014) y en Colombia 42 especies (8.6%, total: 500 especies) (Solari et al. 2013). Ambos casos representan especies endémicas usando el criterio de restricción dentro del país. No obstante, en México se consideró una clasificación de los mamíferos endémicos restringidos y ampliamente distribuidos. Los mamíferos endémicos restringidos presentaban áreas menores a 50,000 km2 y los confinados dentro de México fueron considerados endémicos ampliamente distribuidos (Ceballos et al. 1998).

Figura 2 Mapas de distribución de cuatro especies de mamíferos considerados como especies restringidas por tener un área menor a 50,000 km2. De acuerdo con este criterio, las cuatro especies pueden ser consideradas endémicas. El primate Callicebus lucifer Thomas, 1914 cumple con los criterios de endemismo de área restringida y restricción dentro de los límites administrativos de un país (Colombia); por lo tanto, es endémico de la Amazonía colombiana y endémico de Colombia.
Variantes del endemismo
Una interpretación adicional del endemismo fue propuesta por González-García & Gómez de Silva Garza (2003), quienes consideran que las especies endémicas pueden ser categorizadas en cuasiendémicas y semiendémicas. Las especies cuasiendémicas “son aquellas que penetran ligeramente a algún país vecino debido a la continuidad de los hábitat o sistemas orográficos”. Por otra parte, las especies semiendémicas se definen como “las especies que son endémicas a un país o a una región durante una época del año” (González-García & Gómez de Silva 2003). Las anteriores variaciones del “endemismo” coinciden con la propuesta de la Unión Internacional para la Conservación de la Naturaleza, con base en la restricción y nativo de una isla pequeña, de un país o de un continente (UICN 2012).
Por otra parte se ha intentado relacionar el endemismo con la historia de los taxones y su distribución. Se ha propuesto el paleoendemismo para definir a los taxones antiguos y restringidos a un área geográfica (Prentice 1976; Major 1988), los cuales inicialmente tuvieron amplias áreas de distribución (Malik 2015). Son taxones sistemáticamente aislados, tales como géneros representados por una única especie. Sus áreas de distribución representan remanentes de áreas de distribución amplias, pero que se han reducido por ejemplo a causa de variaciones climáticas. Desde la perspectiva filogenética los taxones paleoendémicos son taxones que divergieron muy temprano y tienen distribuciones que se superponen con formaciones geológicas antiguas (Ferreira & Boldrini 2011).
En otro lugar está el neoendemismo para los taxones recientes y restringidos (Prentice 1976). Son taxones extremadamente divergentes, con sus distribuciones que se traslapan con formaciones geológicas recientes (Ferreira & Boldrini 2011). Se considera que este tipo de taxones podrían estar listos para expandir sus áreas de distribución y también sus genes (Kruckeberg & Rabinowitz 1985), debido a que en plantas resultan de procesos de hibridación con la consecuente poliploidía. Lo cual les confiere adaptación divergente a diferentes condiciones ambientales (Malik 2015).
La desventaja en la aplicación de los términos paleoendemismo y neoendemismo es que estos son relativos a las edades de los taxones dentro del grupo de comparación. Por ejemplo, según Noguera-Urbano & Escalante (2015) en los Andes del Norte hay cinco especies de mamíferos endémicos (Thomasomys aureus [Tomes 1860], Cuniculus taczanowskii [Stolzmann 1865], Didelphis pernigra J.A. Allen 1900, Marmosops impavidus Tschudi 1845 y Monodelphis adusta [Thomas 1897]). Los cuales están estrechamente relacionados a la evolución de los Andes. Al comparar las edades de las cinco especies, la zarigüeya M. impavidus (2.5 Ma) (Jansa et al. 2013) con la menor edad sería neoendémica, mientras que la zarigüeya de cola corta M. adusta (20.5 Ma.) (Vilela et al. 2015) con la mayor edad podría ser considerada paleoendémica (Fig. 3). Sin embargo, al hacer la comparación entre dos de las cinco especies, el ratón T. aureus (3.1 Ma) (Parada et al. 2015) sería neoendémica y la paca C. taczanowskii (3.8 Ma) (Upham & Patterson 2012) sería paleoendémica.

Figura 3 Aplicación del neoendemismo (n) y paleoendemismo (p) a dos especies de mamíferos andinos. (a) Mapas de distribución de las dos especies. (b) Tiempo aproximado de divergencia de las dos especies. H = Holoceno, Modificado de Noguera-Urbano 2016.
En botánica se propuso una clasificación que relaciona el endemismo con la edad de los taxones, inferida a partir del conjunto de cromosomas (Contandriopoulos 1962; Major 1988). En dicha clasificación se asume que las plantas endémicas con baja ploidía (conjunto de cromosomas), posiblemente divergieron más tempranamente que los que presentan alta ploidía (Favarger & Contandriopoulos 1961; Bruchmann & Hobohm 2014). Favarger y Contandriopoulos (1961) propusieron que los paleoendémicos y patroendemismos son los grupos antiguos, mientras que los apoendémicos y esquizoendémicos son los jóvenes o recientes. La poliploidía puede provocar la formación de nuevas especies, así el patroendemismo define a las plantas diploides cuyos progenitores son poliploides que presentan distribución amplia (Fig. 4). El apoendemismo define a plantas poliploides derivadas de diplodes ampliamente distribuidos (Fig. 4) y esquizoendemismo para las plantas cuyo número de cromosomas se ha conservado (ploidía) (Fig. 4) (Contandriopoulos 1962; Major 1988).

Figura 4 Esquemas que representan los tipos de endemismo de acuerdo con la evolución cromosómica de plantas. (a) Especie poliploide ampliamente distribuida. (b) Especie diploide ampliamente distribuida. Los rectángulos y cuadros representan áreas de distribución hipotéticas, los cuales cambian de tamaño por un evento de vicarianza (triángulos). n= número de cromosomas.
Aunque la evolución cromosómica es una estrategia evolutiva muy importante en plantas y otros grupos, se poco claro cómo la ploidía aporta en la definición de endemismo (Bruchmann & Hobohm 2014). Por ejemplo las especies de plantas Allium dirphianum Brullo & al., A. calamarophilon Phitos & Tzanoukadis, Asperula euboea, Campanula constantini Beauverd & Topali, C. cymaea Phitos, Chaerophyllum euboeum Halácsy e Hypericum fragile Heldr. & Sart son consideradas endémicas de la las Islas del oeste del Egeo (Trigas & Iatrou 2006). A la vez A. calamarophilon, Asperula euboea, Campanula constantini y C. cymaea están clasificadas como esquizoendémicas (Trigas & Iatrou 2006).
Al momento la clasificación del endemismo con base en los cromosomas ha sido superada por clasificaciones basadas en el tiempo y el lugar de divergencia. De ello resulta que se haya relacionado el concepto de endemismo con el sitio de evolución de las especies (van Steenis 1964; Merckx et al. 2015). Se considera el “endemismo céntrico” para definir a las especies endémicas que han tenido una evolución in situ (Fig. 5) (Merckx et al. 2015) y el “endemismo excéntrico” es usado para las especies que han migrado de sus sitios de divergencia (Merckx et al. 2015) (Fig. 5). En un sentido práctico de los anteriores conceptos, Merck et al. (2015) definieron como endémicas a aquellas especies con poblaciones restringidas a su área de muestreo (Parques Kinabalu y Crocker Range en Malasia) y relacionaron el sitio de dispersión tomando como base de comparación el piso altitudinal que ocupan la especie endémica y su congénere cercana (Merckx et al. 2015). Los autores concluyeron que las especies endémicas en estudio, pueden evolucionar en el sitio actual de desarrollo o a partir de áreas vecinas (Merckx et al. 2015).

Figura 5 Representación esquemática del endemismo céntrico y excéntrico. El triángulo representa un área geográfica, mientras que los rectángulos indican las áreas de distribución de dos especies. En la especie endémica excéntrica las flechas indican la movilidad de la distribución desde el tiempo de divergencia (Tiempo 1) hasta el Tiempo 3, cuando la especie se acerca parcialmente a su sitio de divergencia.
Una propuesta que ha sido levemente discutida es la de aplicar el endemismo según el concepto de especie de referencia (González-García & Gómez de Silva 2003); por ejemplo el “endemismo biológico” toma en cuenta el concepto biológico de especie (Dobzhansky 1935; Mayr 1942), en el cual es posible reconocer subespecies endémicas; y el “endemismo filogenético” se basa en el concepto filogenético (Cracraft 1989). Con la aplicación del concepto filogenético de especie muchas subpoblaciones consideradas como subespecies con el concepto biológico de especies, pasan a ser especies. Por lo tanto, con el endemismo filogenético, sólo se reconocen especies endémicas (González-García & Gómez de Silva 2003). Considero que la definición de los taxones con base en linajes, grupos monofiléticos, etc. contribuyen a la cuantificación del endemismo, no obstante, la diferenciación de los dos tipos de endemismo aporta poco a la base conceptual del endemismo, la cual sigue siendo la restricción geográfica a un área. Por otra parte, para las especies extintas la información filogenética puede ser incompleta o ausente. Así el endemismo filogenético de González-García y Gómez de Silva (2003) se vuelve impráctico. Si se toma el endemismo para considerar subespecies, como es el caso de taxones fósiles, el único criterio empleado es la restricción a un área geográfica determinada. Por ejemplo el ciervo siciliano Cervus elaphus siciliae es una subespecie endémica de Sicilia, registrada a partir de depósitos del Pleistoceno medio en Sicilia (Mangano 2005).
La necesidad de integrar múltiples dimensiones de la diversidad biológica en la interpretación del endemismo, ha llevado a que se interprete el termino dentro de un contexto filogenético. En este sentido Faith et al. (2004) propusieron el endemismo filogenético (EF) con base en la diversidad filogenética (DF), para representar la historia evolutiva singularmente representada por un conjunto de taxones en un área geográfica (Faith et al. 2004). Aunque Faith et al. (2004) fueron ambiguos en su definición, Rosauer et al. (2009) incluyen a la restricción geográfica como determinante del endemismo filogenético, una medida filogenética de endemismo absoluto. Una ventaja del endemismo filogenético es que permite incluir directamente datos filogenéticos para la interpretación de patrones espaciales del endemismo.
Área endémica, área de endemismo y centro de endemismo
La aplicación del endemismo ha llevado a crear otros términos relacionados: el área endémica, el centro de endemismo y el área de endemismo. Inicialmente la presencia de un taxón endémico permitió caracterizar a una zona geográfica como área endémica (Polunin 1960; Nelson & Platnick 1981; Rosen 1988; Axelius 1991; Parenti & Ebach 2009). Polunin (1960) (p. 205) afirma que “un área endémica es el área de una especie u otro taxón que, en su distribución, se limita a cierta región natural o hábitat únicos por causas históricas y/o ecológicas”. En este contexto un taxón endémico presentaba un área endémica, así que cualquier taxón nativo y restringido podía ser endémico a alguna área (Anderson 1994). El endemismo es relativo al área geográfica que se tome para la comparación (Cracraft 1985), así que un “área endémica” podía ser un concepto equivalente al área de distribución (Polunin 1960).
La puesta en práctica del endemismo en estudios ecológicos y conservación, ha requerido generalizar el área endémica para describir zonas habitadas por varias especies endémicas restringidas. De modo que la superposición, recurrencia o concentración de áreas de distribución de especies endémicas definen a un patrón de endemismo. Por ejemplo un área de aves endémicas será aquella área ocupada por varias especies con distribuciones restringidas (menores a 50,000 km2) (Stattersfield et al. 1998). Desde esta perspectiva el área endémica revisada por Polunin (1960) toma otro sentido en conservación, al representar un patrón de endemismo conformado por varias especies restringidas y que se codistribuyen sólo en esa área.
Por otra parte, un área de endemismo es una zona geográfica definida por la congruencia espacial de las áreas de distribución geográfica de dos o más especies (Platnick 1991; Espinosa et al. 2001; Noguera-Urbano 2016). La congruencia espacial es explicada por procesos diferentes al azar (Morrone 1994) y representa la integración de los taxones en el espacio y tiempo en estratos temporales (Fig. 6) (Noguera-Urbano 2016). Tanto las áreas endémicas de Stattersfield et al. (1998) como las áreas de endemismo representan patrones de endemismo definidos por límites geográficos, sin embargo las áreas de endemismo pueden presentar cualquier tamaño en su área. De acuerdo a lo anterior se podrían tomar algunos conceptos como equivalentes. Es decir, si el endemismo de Candolle (1820) en el contexto histórico define el “área endémica” como aquella área donde se encuentra un taxón endémico, el “área endémica” (Polunin 1960) estaría representando una unidad geográfica similar a la de Candolle. Para generalizar el uso de los términos y evitar malas interpretaciones sobre el endemismo, considero que se debería homologar los conceptos de Candolle (1820) y Polunin (1960) para área endémica, mientras que se podría generalizar el uso de “áreas de endemismo” para definir los patrones de endemismo.

Figura 6 Modelo simplificado de un área de endemismo. (a) Vista actual de las áreas de distribución integradas en el espacio-tiempo. (b) Como se verían las distribuciones separándolas con respecto a la edad de los taxones endémicos. Se indica el espacio geográfico (EG) con el cual se referencia el área de endemismo, un centro de congruencia (CC), especies endémicas (u, v, w, x, y, z) y dos cenocrones o estratos temporales (C1, C2). Modificado a partir de Noguera-Urbano (2016).
Otra manera de representar el endemismo son los centros de endemismo (Crisp et al. 2001; Linder 2001a). Los centros de endemismo son sitios con alta concentración de especies que presentan áreas de distribución restringida (Williams et al. 1996; Crisp et al. 2001; Linder 2001b). El supuesto de los centros de endemismo es que las áreas de distribución de los taxones restringidos se superponen unas sobre otras y que los patrones de endemismo que conforman se traslapan unos sobre otros en la naturaleza (Crisp et al. 2001). Sin embargo, se ha propuesto que es difícil comprobar que los taxones sean endémicos a los centros de endemismo, o que las distribuciones de las especies tengan congruencia espacial (Linder 2001a).
Desde mi punto de vista, los tres términos relacionados con endemismo: área endémica (según Stattersfield et al. 1998), centro de endemismo y área de endemismo, toman en cuenta que las áreas de distribución de los organismos proveen información sobre la relación compleja entre los ambientes físicos y los atributos biológicos de los organismos. Sin embargo, emplean criterios diferentes para delimitar el endemismo. Las áreas endémicas y los centros de endemismo se basan en la restricción a un área de tamaño y límites arbitrarios, mientras que las áreas de endemismo se basan en la superposición de las áreas de distribución geográfica.
Con respecto a la integración evolutiva de los taxones, Linder (2001a) ha mencionado que en los centros de endemismo los taxones presentan historias múltiples y que las áreas de endemismo contienen biota con historia única. En mi opinión, en las áreas endémicas según Stattersfield et al. (1998), los centros de endemismo y las áreas de endemismo, los taxones se encuentran integrados en el espacio, pero tienen múltiples historias. Pero en algunos casos los taxones pueden conformar subgrupos y tener historias compartidas en el tiempo. Por ejemplo en el área de endemismo Andina la integración de los taxones endémicos ha ocurrido en diferentes tiempos en la evolución de América del Sur, pero todos han sido afectados por el levantamiento de los andes y la orogenia de la zona (Fig. 6) (Noguera-Urbano 2016).
Se considera que las áreas de endemismo y los centros de endemismo pueden ser representados valorando tanto la concentración de las áreas de distribución de las especies endémicas (continuidad del patrón de endemismo) como sus límites geográficos (Cracraft 1985; Anderson 1994; Crisp et al. 2001; Noguera-Urbano 2016). En una perspectiva integradora de los dos conceptos “centros y áreas de endemismo”, se podría considerar que las áreas de endemismo al tener un centro de congruencia con valores máximos de endemismo (Fig. 6) (Noguera-Urbano 2016), podrían contener centros de endemismo. En un área de endemismo los centros de endemismo estarían conformados por la alta congruencia de las especies endémicas. Además, las áreas de endemismo pueden estar superpuestas conformando diferentes estratos de integración temporal, estratos temporales (Fig. 6), y estarían representando historias múltiples como los centros de endemismo.
Métodos de cuantificación del endemismo y patrones basados en restricción
Una vez definidas las especies distribuidas en algún área geográfica, es posible aplicar diferentes tipos de descriptores para cuantificar el endemismo (Apéndice I) e identificar áreas de endemismo (Apéndice II). La cuantificación del endemismo se relaciona con la estimación de la riqueza de taxones endémicos por unidad de área (país, provincias, celdas) con el objetivo de hacer comparaciones, conocer la distribución del endemismo en un área geográfica, evaluar la correlación del número de especies endémicas con la riqueza de especies o dirigir esfuerzos de conservación. Por otra parte, la identificación de áreas de endemismo se dirige a conocer y describir patrones evolutivos con base en las distribuciones de las especies, en ellas se toma como argumento teórico que la tierra y la biota evolucionaron juntos. Con los métodos de identificación de áreas de endemismo se busca proponer hipótesis biogeográficas que puedan ser interpretadas para explicar la integración espacio-temporal de las especies endémicas.
En los métodos usados para cuantificar el endemismo se aplica el endemismo como “distribución restringida” (Apéndice I). Estos métodos son sencillos en su aplicación, pero pueden ser poco útiles en la comparación entre regiones de diferente tamaño (Major 1988). Por ejemplo la estimación de la Riqueza de endemismo y Proporción de endemismo son medidas directas del número de especies endémicas dentro de un área o con relación a un área respectivamente (Apéndice I). Ambas métricas brindan poca información sobre el área de distribución de las especies y no toman en cuenta el tamaño de las unidades de comparación (Kier & Barthlott 2001).
Otros métodos como el Índice de Bykow (Major 1988; Hobohm & Tucker 2014), la densidad de endemismo (Hobohm & Tucker 2014) y la relación área-endemismo (Major 1988; Harte & Kinzig 1997; Hobohm & Tucker 2014) son empleados para evaluar la concentración de endemismos entre unidades geográficas (continentes, países, celdas, etc.). Los tres índices incluyen en sus ecuaciones el tamaño de las áreas de distribución y el área geográfica analizada (Apéndice I), no obstante, su interpretación se debe realizar con cuidado, ya que el supuesto básico es que las especies están bien muestreadas en el área de estudio (Kier & Barthlott 2001). En el caso de la densidad de endemismo y la relación área-endemismo su aplicación se vuelve poco práctica, cuando las áreas en comparación tienen tamaños diferentes (Hobohm & Tucker 2014).
La selección de taxones endémicos con criterios arbitrarios y basados en un valor absoluto del endemismo, son factores condicionantes al momento de estudiar y comparar sitios de interés. Así que se desarrollaron índices para la evaluación de la distribución espacial del endemismo y la identificación de sitios de alta concentración de endemismos (Apéndice I), los cuales son representados en mapas de endemismo. Los patrones de concentración de especies endémicas fueron denominados como áreas de rareza (Williams et al. 1996) o centros de endemismo (Crisp et al. 2001; Linder 2001a), los cuales son conceptos equivalentes, si se toma en cuenta que representan concentraciones de sitios con los mayores valores de endemismo (Williams et al. 1996; Crisp et al. 2001; Linder 2001b).
Para la identificación de centros de endemismo fueron desarrollados los índices de Endemismo Ponderado (WE) y Endemismo Ponderado Corregido (CWE), que toman como unidad de comparación celdas de una gradilla (Apéndice I). El CWE es una modificación del WE, con el cual se busca disminuir la correlación entre la riqueza de especies y el endemismo (Crisp et al. 2001; Linder 2001b). El CWE facilita la identificación de centros de endemismo pobres en especies (Crisp et al. 2001), pero en sitios con bajo muestreo de especies ampliamente distribuidas el índice puede resultar inflado (Slatyer et al. 2007).
Otra modificación al WE, denominada Riqueza de endemismo (Cs), fue propuesta por Kier & Barthlott (2001) con el objetivo de generalizar el índice para cualquier tipo de UGO (gradillas, corotipos, biomas, ecoregiones, etc.). En este índice se considera que cada especie contribuye al inventario general del área, se incluye el tamaño del área de distribución y su valor complementario dentro del área estudiada como factores de la formula (Apéndice I). Los tres métodos coinciden en descartar el uso de un criterio a priori para identificar a las especies endémicas (Williams et al. 1996; Lovett et al. 2000).
En búsqueda de aumentar el peso de las especies endémicas en los índices, se ha propuesto el uso de ventanas (v.g. 3x3 celdas) para estimar el endemismo. Sobre las ventanas de análisis se calcula el índice WE y su valor se relativiza con la riqueza de especies por ventana (Slatyer et al. 2007). Laffan, Lubarsky & Rosauer (2010) propusieron otros cambios a los índices WE y CWE. Algunos índices del endemismo se encuentran disponibles en el programa Biodiverse (Laffan et al. 2010), incluyendo los WE y CWE normalizados con el número de celdas que componen el área de estudio (Apéndice I). Otras aplicaciones incluyen la identificación de endemismo con el uso de ventanas para diferentes niveles taxonómicos (orden, familia, género) en el llamado Endemismo Completo con Partición Jerárquica (Laffan et al. 2013) (Apéndice I), bajo el supuesto que cada nivel taxonómico contribuye al endemismo de un taxón.
A partir del concepto de endemismo filogenético de Rosauer et al. (2009), se han propuesto índices que integran la diversidad filogenética y los índices WE y CWE, por ejemplo el Índice de Endemismo Filogenético (EF) (Apéndice I). El EF representa la cantidad de diversidad filogenética de un taxón o clado restringido a un área geográfica con base en dos criterios (Rosauer et al. 2009): a) mide la variación del endemismo a través de cada unidad (celdas) dentro del área de estudio; y b) integra todos los niveles filogenéticos representados en la filogenia usados en el análisis. Los dos criterios hacen que el EF pueda ser medido para cualquier área de interés, a diferentes escalas y además que dependa poco de los niveles taxonómicos de los taxones analizados. Cerca de otras 12 modificaciones se han desarrollado al EF para incorporar las correcciones realizadas a los índices WE y CWE (Laffan et al. 2010). Dos ejemplos de índices usados para describir el EF son los Rareza Filogenética Corregida-Ponderada y el Endemismo Filogenético Ponderado Corregido (variante central). El primer índice incluye la abundancia de los taxones, buscando dar mayor peso a las especies raras. Mientras que el segundo considera grupos de celdas (ventanas) para estimar la restricción de la DP. Otras modificaciones al EF ingresan como factor la simulación de pérdida de clados en la filogenia, con la intensión de evaluar el efecto de la disminución de taxones superiores sobre la PD y por lo tanto el endemismo. Las anteriores variantes de la DP se encuentran también implementadas en el programa Biodiverse (Laffan et al. 2010).
Métodos de identificación de áreas de endemismo
En biogeografía evolutiva se toma al endemismo como restricción a un área sin importar el tamaño. Las áreas de endemismo se identifican a partir de la evaluación de la congruencia espacial de las áreas de distribución de las especies, para retener a aquellas áreas soportadas por las especies con alta congruencia espacial. Sin embargo, la descripción de patrones de endemismo ha sido controversial por la ausencia de un valor cuantificable de la congruencia espacial (Henderson 1991). Esto ha llevado a pensar que el uso de la congruencia espacial como criterio para la identificación de las áreas de endemismo, puede ser poco útil cuando se trata de especies endémicas sin congruencia espacial que habitan islas (Harold & Mooi 1994).
En biogeografía evolutiva de acuerdo con la interpretación de la congruencia espacial y la puesta a prueba de su validez, se han desarrollado aproximadamente 11 métodos para la identificación de patrones de endemismo “áreas de endemismo” (Apéndice II): (a) los que generalizan las presencias a unidades geográfica operativas (celdas, ecosistemas o tipos de vegetación, etc.) y luego las comparan para identificar grupos, medidos a partir de la alta congruencia espacial de los taxones; (b) los que toman como distribuciones a las localidades de presencia y por lo tanto la congruencia se mide a ese nivel; y (c) aquellos métodos que buscan probar la significancia matemática de la congruencia espacial, a partir de la evaluación de la no aleatoriedad de las áreas de endemismo.
En el primer grupo se encuentran el Análisis de Parsimonia de Endemismos (PAE) (Rosen & Smith 1988), el Análisis de Parsimonia de Endemismos para celdas (PAE) (Morrone 1994) y el Análisis de Endemicidad o Criterio de Optimización (AE) (Szumik et al. 2002; Szumik & Goloboff 2004). La identificación de áreas de endemismo en el PAE puede incluir localidades, provincias, celdas y otros UGOs, mientras que en el AE las distribuciones son representadas solo por localidades o el uso de matrices de taxones por celdas.
Se ha mencionado que los métodos que jerarquizan UGOs desestiman la representación de los sistemas naturales (Szumik et al. 2002; Casagranda et al. 2009), así que la diferencia entre el PAE y el AE, se fundamenta en esa filosofía. Con el PAE se identifican sistemas jerarquizados de áreas de endemismo, mientras que con el AE se obtienen áreas de endemismo superpuestas. En cuanto a la aplicación metodológica, el PAE ha sido criticado por la inclusión indirecta del componente espacial. Por esta razón en el AE se comparan celdas y se obtiene un índice de endemismo para cada especie y área de endemismo, esto involucra el componente espacial dentro de la evaluación del endemismo (Szumik et al. 2002; Casagranda et al. 2009).
El segundo grupo está integrado por el Método de Análisis de Redes (NAM) (Dos Santos et al. 2008) y la Interpolación Geográfica del Endemismo (GIE) (Oliveira et al. 2015). En el NAM se evalúa la congruencia a nivel de localidades (Dos Santos et al. 2008), haciendo implícito el componente espacial para identificar áreas de endemismo. Sin embargo, los patrones con poca congruencia detectadas en el NAM han sido criticados por considerar que cumplen vagamente con el criterio de área de endemismo (alta congruencia espacial) (Casagranda et al. 2009). La comparación de las distribuciones en el NAM, se realiza considerando la conexión entre localidades y la posterior identificación de patrones con análisis de redes. El análisis de redes permite incluir a las especies y sus relaciones espaciales (Dos Santos et al. 2008), esto lo diferencia del PAE y el AE que son dependientes de celdas de grillas.
El GIE es una adaptación de la interpolación espacial Kernel para la representación del endemismo (Oliveira et al. 2015). Este método define áreas circulares de influencia alrededor de los puntos de presencia, las cuales son usadas para evaluar su traslape con una función de densidad Kernel y representar el endemismo como un continuo en el espacio geográfico. Las selecciones de los radios para comparar las distribuciones son definidos por el usuario (Oliveira et al. 2015), por lo tanto, la distribución de los valores de endemismo podría depender del radio que se tome.
En cuanto a la implementación de los métodos, el PAE puede ser ejecutado en programas que tengan parsimonia como método filogenético, el AE ha sido desarrollado en el programa NDM/VNDM (Goloboff 2014) y NAM se encuentra en el paquete SyNet del programa R (Dos Santos et al. 2008; Dos Santos 2011). Mientras que el GIE se encuentra como extensión de un programa comercial de Sistemas de información geográfica (Oliveira et al. 2015).
En el tercer grupo se encuentran el reconocimiento de áreas de endemismo de Harold & Mooi (1994), el criterio de optimización para evaluar métodos de endemismo (Linder 2001a), el Análisis de Co-presencia significante de taxones (Sigcot) (Mast & Nyffeler 2003), el Análisis de Endemismo de Áreas Anidadas (NAEA) (Deo & DeSalle 2006) y Sigcot con PAE (Giokas & Sfenthourakis 2008). Harold & Mooi (1994) criticaron el uso de la congruencia espacial como único criterio para identificar áreas de endemismo y propusieron la inclusión de información filogenética para la identificación de áreas de endemismo (criterio de congruencia espacial y filogenética). El protocolo es teórico y hasta el momento se encuentra sin implementar.
El Sigcot permite filtrar los taxones antes de aplicar cualquier método para la identificación de las áreas de endemismo. Se fundamenta en la evaluación de la congruencia espacial, con modelos nulos. Para ello se aleatoriza las distribuciones y se mide la congruencia entre las distribuciones generadas. Si la congruencia observada se encuentra con frecuencia alta en las comparaciones de las distribuciones generadas, entonces la congruencia podría ser explicada por el azar (Mast & Nyffeler 2003). En el criterio de Sigcot (Linder 2001a) y Sigcot con PAE se busca poner a prueba las áreas de endemismo y la congruencia espacial, y se ejecutan antes o después de identificar las áreas de endemismo. En el Sigcot (Linder 2001a) se desarrolló el índice de congruencia (CON, Apéndice II) el cual pondera el tamaño del área de distribución con el tamaño del área de endemismo. Este índice mide el ajuste de la distribución de las especies endémica al área de endemismo. El Sigcot con PAE, sigue los principios básicos del Sigcot pero en lugar de ejecutar los modelos nulos sobre la matriz completa, la comparación se hace con los taxones en cada área de endemismo (Giokas & Sfenthourakis 2008).
El NAEA es una adaptación del análisis de clados anidados (Deo & DeSalle 2006), empleado en filogeografía para el estudio de flujo de genes y poblaciones (Templeton 1998). Una de las ventajas del NAEA, es la evaluación de la significancia de la congruencia espacial para identificar las áreas de endemismo (Deo & DeSalle 2006). La desventaja general para este y otros métodos que generalizan las distribuciones a celdas, es la dependencia del resultado al tamaño de celda, especies endémicas con un tamaño de celda particular podrían no serlo con otro tamaño de celda (Morrone & Escalante 2002). Con respecto a la implementación del NAEA, este método se puede ejecutar en el programa original para el análisis filogeográfico de clados anidados (Deo & DeSalle 2006).
Problemas asociados a la cuantificación del endemismo y a la descripción de patrones de endemismo
Los resultados de los cálculos de los índices para el análisis del endemismo pueden ser afectados por variaciones en las variables de las fórmulas (riqueza de especies, tamaño del área de estudio, tamaño de celdas, sesgos de muestreo, etc.). En el caso de los métodos de cuantificación del endemismo, la evaluación de la concentración del endemismo o la identificación de áreas de endemismo que son dependientes de celdas, el efecto del tamaño y divisiones de la gradilla pueden ocultar los patrones (Linder 2001a; Morrone & Escalante 2002). Para el PAE, NDM, entre otros, la modificación del tamaño de celdas altera la generalización de las distribuciones a las celdas, así que un taxón que es endémico a una escala, podría no serlo en otra (Morrone & Escalante 2002).
La extensión del área de estudio puede condicionar la ubicación de los centros de endemismo o la identificación de las áreas de endemismo. Si se incluyen áreas de distribución incompletas, los índices de cuantificación del endemismo cambian, debido al efecto que tiene el número de celdas sobre la ponderación del endemismo (Lovett et al. 2000). En los métodos de identificación de áreas de endemismo este punto ha sido poco explorado, no obstante, se podría esperar que se identifiquen áreas de endemismo falsas al encontrar congruencia espacial alta entre un área de distribución completa y el fragmento de otra (Fig. 7).

Figura 7 Mapas de distribución de tres especies de mamíferos. (a) Áreas de distribución completas de las tres especies, Allouta pigra Lawrence, 1933 y Cryptotis mayensis (Merriam 1901) pueden ser consideradas que soportan a la Península de Yucatán como área de endemismo. (b) La distribución de Carollia sowelli Baker, Solari & Hoffmann, 2002 está incompleta en el mapa, durante un análisis de endemismo que incluya la panorámica presentada las tres especies podría soportar a la Península de Yucatán como área de endemismo. En este caso C. sowelli es un falso endemismo de la Península, resultado de incluir una distribución incompleta.
En los índices que consideran ventanas o radios para estimar la densidad durante la ponderación, se ha mencionado que el movimiento y tamaño de las ventanas incrementan los valores de endemismo (Laffan et al. 2013). Esto se relaciona con el uso de la riqueza para corregir el índice de endemismo, donde la riqueza alrededor de la celda objetivo puede ser variable. Por ejemplo, usar una ventana de cuatro celdas podría involucrar conteos incompletos, en comparación con una ventana de nueve o más celdas. En el caso del GIE, modificaciones en el tamaño del radio de análisis, ocasiona cambios en la densidad del endemismo.
La dependencia de todos los métodos al conjunto de taxones analizados es alta, la adición o sustracción de especies afecta la evaluación del endemismo (Linder 2001b). Los índices de riqueza de endemismo, relación endemismo-riqueza total y densidad, se vuelven incomparables si existen vacíos en información sobre la riqueza total y distribuciones de los taxones. Por ejemplo, si se comparan las riquezas de mamíferos endémicos en el presente y su representatividad en el Cuaternario (Cuadro 1), se observa que los valores más altos de endemismo se encuentran en el presente. Esta es una conclusión poco acertada, considerando que el conocimiento sobre la riqueza de mamíferos endémicos del Cuaternario y otros periodos es incompleto. Por lo tanto, se considera que los patrones de endemismo son hipótesis que deben ser probadas con nuevos datos (Harold & Mooi 1994), analizadas con diferentes enfoques metodológicos y generalizadas a partir de comparaciones con otros patrones (Morrone 1994). El desarrollo de protocolos para la evaluación del endemismo y los patrones que conforma aún es un campo en desarrollo.

Perspectivas
La mayoría de índices iniciaron con la descripción de patrones espaciales del endemismo, pero se ha progresado en el desarrollo de índices como el endemismo filogenético, con el objeto de darle peso a la historia evolutiva de los taxones. Sin embargo, se considera que la inclusión de las dimensiones espacio-tiempo y forma podría ser un nuevo enfoque para el análisis de los patrones de endemismo (Noguera-Urbano 2016).
En general el término endémico se refiere a la exclusividad de un taxón a un área geográfica. Por ejemplo el roedor Neotoma cinerea (Ord 1815) es considerado endémico, porque está restringido al norte del Pacífico de América del Norte (Zabel & Anthony 2003). Sin embargo, los registros fósiles del Pleistoceno indican que la especie se encontraba en México (Ferrusquía-Villafranca et al. 2010). Este caso refleja el sentido variable de la aplicación del endemismo, al ser tomado como restricción geográfica. Las especies que en la actualidad son consideradas endémicas, pudieron tener mayores o menores distribuciones en el pasado o estar restringidas en zonas geográficas diferentes a las actuales. La identificación de patrones de endemismo requiere un conocimiento aceptable de las distribuciones de los taxones. En el análisis de provincialismo de mamíferos pleistocénicos en México (Ferrusquía-Villafranca et al. 2010), los autores integraron la información de mamíferos extintos y existentes para identificar patrones. A partir de ello se propusieron ocho corredores que permitieron el flujo de mamíferos a lo largo y dentro del territorio mexicano. Esto demuestra que los patrones biogeográficos pueden ser recuperados usando distribuciones multitemporales integradas. En la actualidad son pocos los análisis que incluyen fósiles para estudiar patrones de endemismo, este puede ser un nuevo camino a explorar.
Generalmente en los índices de endemismo filogenético se estima la historia evolutiva de los taxones sobre una o más celdas y luego se comparan entre conjuntos de celdas. En este proceso se deja un lado el análisis de la historia independiente de cada taxón endémico. La composición de los taxones en los patrones de endemismo es un punto que empieza a ser explorado. Por ejemplo, para las áreas de endemismo se sabe que su estructura puede ser asincrónica con respecto a la edad de cada taxón (Noguera-Urbano, 2016). Se piensa que en las áreas de endemismo pueden ocurrir múltiples tiempos de integración, dependiendo de la evolución de los taxones endémicos en cada área de endemismo. Por lo tanto, es necesaria su evaluación integral para dejar de percibir al endemismo como restricción, e interpretarlo dentro del contexto evolutivo (unicidad). Además, hace falta explorar e incorporar en los índices la forma de los taxones representada en atributos biológicos. La inclusión de mayor información biológica podría mejorar la interpretación del endemismo en las tres dimensiones (espacio- tiempo-forma).
Conclusiones
En ecología y conservación el endemismo se interpreta con relación a un área geográfica de referencia y dentro un marco temporal determinado. Mientras que en biogeografía evolutiva se considera al endemismo como restricción a un área natural sin importar el tamaño, además permite identificar patrones que han perdurado a través del tiempo.
Tradicionalmente en ecología el endemismo bajo criterios prácticos es relacionado con la restricción de una especie a un área determinada a priori o los límites geopolíticos. Esto permite hacer comparaciones entre áreas o priorizar sitios donde el endemismo toma alto valor. Mientras que en la perspectiva evolutiva generalmente se usan límites naturales para definir taxones endémicos y la congruencia espacial como criterio práctico para la determinación de taxones endémicos.
Las áreas endémicas, áreas de endemismo y centros de endemismo representan unidades geográficas derivados de la aplicación de diferentes criterios de endemismo. En el primero se usa como criterio “distribución restringida”, mientras que la presencia de dos o más especies que pueden tener o no congruencia espacial soportan el área endémica. En las áreas de endemismo los taxones endémicos son aquellos que habitan áreas de cualquier tamaño, pero que tienen alta congruencia espacial con al menos otro taxón. En los centros de endemismo los taxones endémicos son identificados con base en el criterio de distribución restringida, y el endemismo es relativizado en el espacio geográfico de acuerdo al tamaño de área de distribución de cada especie endémica.
Todo lo anterior demuestra la necesidad de un concepto unificado del endemismo, es decir, se podría generalizar la aplicación de la definición de Candolle (1820) para referirse a taxones nativos y de distribución única, aplicable a área de endemismo. Y si se toma en cuenta que las áreas endémicas y los centros de endemismo se basan en áreas de distribución restringida, entonces los dos tipos de unidades geográficas se podrían llamar áreas con especies restringidas y centros de distribuciones restringidas respectivamente.
La identificación de las áreas endémicas, áreas de endemismo y centros de endemismo están directamente relacionadas con la estimación y delimitación de las áreas de distribución o con la definición del área de estudio. Por ejemplo, la comparación de áreas de distribución parciales podría resultar en la identificación de áreas de endemismo falsas. Por otra parte, la escala espacial puede tener efecto sobre la caracterización de los patrones. Por ejemplo, en los métodos dependientes de grillas, los índices de endemismo y la forma de los patrones de endemismo puede variar con el cambio en el tamaño de celda. Por ejemplo celdas grandes podrían homogenizar el endemismo dentro del área de estudio, mientras que celdas chicas podrían desagregar el patrón.
Finalmente, la poca inclusión de información de taxones extintos en los análisis biogeográficos de áreas de distribución, puede ocasionar sesgos o pérdida de los patrones de endemismo (Wagner & Marcot 2013). Aunque cuando los patrones son robustos y la calidad de las distribuciones es aceptable, las distribuciones de especies actuales permiten hacer inferencias sobre procesos evolutivos en múltiples marcos temporales (Martínez-Meyer et al. 2004; Wagner & Marcot 2013). Por lo tanto, la inclusión de distribuciones de taxones extintos obtenidas de registros fósiles podría mejorar la identificación e interpretación de patrones biogeográficos propuestos para describir el endemismo.



