










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de fitopatología
versión On-line ISSN 2007-8080versión impresa ISSN 0185-3309
Rev. mex. fitopatol vol.27 no.2 Texcoco ene. 2009
Artículos científicos
Epidemiología de la Antracnosis [Colletotrichum gloeosporioides (Penz.) Penz. and Sacc.] en Mango (Mangifera indica L.) cv. Ataulfo en el Soconusco, Chiapas, México
Anthracnose Epidemiology [Colletotrichum gloeosporioides (Penz.) Penz. And Sacc.] on Mango cv. Ataulfo (Mangifera indica L.) in the Soconusco, Chiapas, Mexico
Graciela Huerta–Palacios1, Francisco Holguín–Meléndez2, Flor Azucena Benítez–Camilo3 y Jorge Toledo–Arreola4
1 Departamento de Entomología Tropical. Correspondencia: ghuerta@ecosur.mx.
2 Laboratorios institucionales.
3 Posgrado ECOSUR.
4 Departamento de Entomología Tropical, El Colegio de la Frontera Sur (ECOSUR), km 2.5 Carr. Antiguo Aeropuerto, Tapachula, Chiapas, México CP 30700.
Recibido: Marzo 22, 2009
Aceptado: Junio 22, 2009
Resumen
Se describe la fluctuación de la antracnosis causada por el hongo Colletotrichum gloeosporioides en follaje de mango cv. Ataulfo, su relación con el desarrollo de brotes en el hospedante y la cantidad de conidios capturados en tres tipos de trampas, bajo las condiciones climáticas del Soconusco, Chiapas, México. El estudio se desarrolló durante los meses de diciembre del 2001 a septiembre de 2002. Los resultados indicaron que la enfermedad crece durante todo el año, afectando hojas jóvenes y maduras, con un patrón de distribución al azar en la copa de los árboles. El mayor incremento de la enfermedad se presentó de diciembre a abril, con igual tasa de crecimiento sobre hojas jóvenes (0.016) y maduras (0.015). Un análisis de regresión múltiple mostró que los parámetros climáticos de mayor correlación con la severidad de la enfermedad fueron la temperatura mínima de punto de rocío (TminPR) y la humedad relativa mínima (HRmin), con correlación positiva y negativa respectivamente. Estas variables explicaron el 83% de las variaciones en el desarrollo de la antracnosis, mientras que las lluvias torrenciales comunes en esta región, coincidieron con una menor incidencia de la enfermedad y solo explicaron el 25% de las variaciones. Valores de TminPR entre 21 a 24°C durante diciembre y abril, propiciaron la condensación de agua sobre hojas, flores y frutos, favoreciendo la infección y dispersión por arrastre de conidios (síntoma de lagrimeo) y salpique.
Palabras clave: clima, hospedero, trampeo de conidios, humedad relativa, temperatura, precipitación pluvial.
Abstract
The present study is aimed to describe the anthracnose fluctuation caused by Colletotrichum gloeosporioides on Ataulfo mango foliage and its relationship with host shoot development and caught conidia quantity, under the climatic conditions from the Soconusco, Chiapas, Mexico. The study was performed from December 2001 to September 2002. The results indicate that the disease increases throughout the year, affecting young and mature leaves with a random distribution pattern into the canopy. The greatest disease increase was observed from December to April, with a similar growth rate between young (0.016) and mature (0.015) leaves. A multiple regression analysis revealed that the most correlated climatic parameters with disease severity were the minimal Temperature of Dew Point (mTDP) and the minimal Relative Humidity (mRH) with a positive and a negative correlation, respectively. Such variables explained 83% of the anthracnose development variations, as the common heavy rains on the region matched with the lowest incidence of the disease and explained only 25% of the variations. mTDP, with values ranging from 21 to 24°C during December and April enabled water condensation over leaves, flowers and fruit and enhanced the infection and dispersion of conidia dragging (tearing symptom) and splashing.
Key words: weather, host, spores trapping, relative humidity, temperature, pluvial precipitation.
INTRODUCCION
La antracnosis del mango (Mangifera indica L.), producida por el hongo C. gloeosporioides (Penz.) Penz. and Sacc., causa graves pérdidas económicas en diversos cultivos de regiones tropicales y subtropicales del mundo (Jeffries et al., 1990; Ploetz, 1994; Prusky et al., 2000). En México, se ha reportado como una enfermedad que afecta hojas, inflorescencias y frutos en las regiones productoras de mango, siendo el daño más importante en poscosecha (García, 1976; Mosqueda–Vázquez et al., 1996; Allende–Molar et al., 2003; Benítez–Camilo et al., 2003). Contrario a esto, en el Soconusco, Chiapas; el daño más fuerte es el atizonamiento de inflorescencias en el cv. Ataulfo, que se presenta durante los meses de noviembre a febrero, después que la época de lluvias termina (Benítez–Camilo et al., 2003).
Una práctica que se generalizó a principios de los 90's fue la inducción forzada de la floración del mango Ataulfo, aplicando nitrato de potasio (2 ó 3 %) durante el mes de octubre, para inducir la floración en noviembre y cosechar los frutos durante enero y febrero. Esto ha permitido obtener un beneficio adicional en el precio de la fruta, por ser el primer mango que llega al mercado. Con el paso del tiempo y de manera gradual se observó una falta de prendimiento de frutos y atizonamiento de flores, a tal punto que en los últimos años la producción de mango temprano ha sido casi nula. Con relación a esto, Benítez–Camilo et al. (2003) mencionan que aplicaciones de cantidades iguales de inóculo de Colletotrichum, sobre panículas presentes en noviembre y enero, causaron en promedio 75.1% y 58.8% de atizonamiento, respectivamente, sugiriendo que esta disminución en severidad fue ocasionada por cambios de temperatura y humedad.
La aplicación de fungicidas de contacto alternados con sistémicos a los 15 días de iniciada la floración y durante la fructificación, es una práctica común en la región. Sin embargo, este programa de aplicaciones no ha proporcionado un control satisfactorio de la enfermedad. Esto coincide con lo observado en Filipinas y Malasia, donde se presentó atizonamiento en flores y una fuerte disminución en el prendimiento de los frutos de mango, cuando la floración coincide con humedad relativa alta (Jeffries et al., 1990). En la isla de Dominica, probaron que aplicaciones de fungicidas posteriores a la floración no controlaron eficientemente dicha enfermedad (Prior y Rider, 1987). Por otro lado, en Florida, E.U.A. se reportó, que las aplicaciones hechas después de la floración no proporcionaron un control adecuado y contribuyeron a la aparición de cepas tolerantes a los fungicidas (Spalding, 1982).
En México, son escasos los estudios sobre la epidemiología de la antracnosis en mango. Algunos antecedentes sobre el tema, fueron generados por Acosta–Ramos et al. (2003), quienes describieron el efecto del manejo integrado sobre la incidencia de enfermedades y calidad del fruto en mango cv. Haden, en el estado de Guerrero. Pero no existen antecedentes acerca de la relación de dicha enfermedad en mango cv. Ataulfo, bajo las condiciones climáticas del Soconusco, Chiapas, México, que puedan apoyar al desarrollo de una estrategia más eficiente de manejo. El presente trabajo tuvo como objetivo estudiar la fluctuación de la antracnosis sobre follaje, su relación con variables climáticas y la producción de follaje nuevo, con la finalidad de observar si hay acumulación de inóculo en follaje antes de la floración.
MATERIALES Y MÉTODOS
Área de estudio. El presente trabajo se llevó a cabo de diciembre del 2001 a septiembre del 2002, en la Finca Pumpuapa, ubicada en el municipio de Tapachula, Chiapas, México, en el km 278, carretera costera Arriaga–Tapachula. En esta localidad hay una estación de lluvias que se presenta de mayo a noviembre, con un rango de precipitación pluvial de 2400–5000mm y temperatura promedio de 26°C (García, 2004). El estudio se realizó en un huerto de árboles injertados con cv. Ataulfo, de 12 años de edad, plantados bajo un arreglo topológico de marco real y una densidad de 56 árboles por ha, con microaspersión durante los meses de enero a mayo, inducción de floración por aspersión de nitrato de potasio (KNO3 al 2 ó 3%) en octubre, y aplicación de fungicidas de noviembre a febrero.
De una población de 135 árboles, se seleccionaron en forma aleatoria 5 árboles y en cada uno de ellos se marcaron 12 ramas, distribuidas hacia los cuatro puntos cardinales y a tres diferentes alturas: 1.5, 2.5 y 4.0 m del suelo, respectivamente.
Descripción de síntomas. Se tomaron muestras de diferentes tipos de lesiones y se colocaron en cámara húmeda por 24 a 48 h, posteriormente fueron observadas al microscopio compuesto para identificar y corroborar la presencia del agente causal. El porcentaje de área foliar afectada por la enfermedad fue medido en las hojas ubicadas entre el primer nudo y la punta de los brotes.
Parámetros de la epidemia. En 60 ramas marcadas, se registró el porcentaje de tejido foliar afectado por antracnosis, el estado fenológico del brote y la cantidad de conidios de C. gloeosporioides presentes en tres tipos diferentes de trampas caza conidios. El follaje enfermo de mango se midió semanalmente, utilizando una escala con diferentes porcentajes de área foliar dañada: 0–1, 3–5, 7–10, 15–20, 25–40, 45–60 y 70100. Se utilizaron fotografías de brotes enfermos, para estimar el área foliar total y el área dañada, según la metodología propuesta por Zadoks y Schein (1979) y Stubbs et al. (1986).
Con el fin de registrar las fechas de brotación y los cambios subsecuentes que tuvieron los brotes, cada semana se registró, el desarrollo de los brotes. Se consideraron brotes jóvenes, aquellos que presentaron hojas de color amarillo–rojizo a verde claro, brillantes, de consistencia suave y flexible y brotes maduros, los de hojas de color verde claro a oscuro, de textura coriácea y poco flexibles. También, se llevó un registró diario de las principales variables climáticas. La cantidad de conidios de C. gloeosporioides, se estimó utilizando tres tipos de trampas: 1)La tipo "roto–rod" marca Ted Brown Associates, para cuantificar conidios diseminados por el viento (Whitney, 1976; Sterling y Lewis, 1998). Esta cuenta con dos varillas de plástico de 6 cm de longitud x 2 mm de ancho, una de las caras fue impregnada con vaselina para que los conidios fueran atrapados (Fig. 1). Para determinar la hora de trampeo al inicio de la época sin lluvias (noviembre) y con lluvia (junio), se hicieron muestreos cada hora a partir de las 6:00 a.m. hasta las 6:00 p.m. Con este fin, se colocó una pértiga de 5 m de largo en el centro del huerto, con tres trampas roto–rod fijadas a 1.5, 2.5 y 4.0 m de altura. La cantidad de conidios por metro cúbico de aire y por hora, se estimó usando las indicaciones del fabricante y con base en estos datos se determinó la hora de trampeo. 2) Para cuantificar los conidios diseminados por salpique o escurrimiento de agua, se colocaron portaobjetos entre las ramas de los árboles, en posición horizontal y con una capa delgada de vaselina en su cara superior y, 3) Trampas tipo "banderitas" de 1 x 1.5 cm, para cuantificar la diseminación de conidios por aire y salpique de agua. Estas fueron hechas con cinta transparente con pegamento en las dos caras, adherida a una aguja entomológica, que al estar insertada en un tubo capilar de 2 cm de alto, permitió que la trampa girara por la acción el viento (Fig. 1).

Los dos últimos tipos de trampas fueron colocados a una altura de 1.5 m, en dirección a los puntos cardinales en cada uno de los cinco árboles sujetos a estudio y ahí permanecieron por siete días, al término de los cuales se estimó la cantidad de conidios por cm2, contando 15 campos de microscopio por trampa (Whitney, 1976; Pérez et al., 1999).
Los factores climáticos que se utilizaron fueron humedad relativa máxima y mínima, días con humedad relativa igual o mayor a 90%, oscilación de la humedad relativa, temperatura máxima, mínima y promedio, temperatura máxima y mínima de punto de rocío, precipitación pluvial total por semana y promedio por hora, evaporación, tensión de vapor, insolación, presión atmosférica y su oscilación semanal. Estos datos fueron registrados por la estación meteorológica nacional del observatorio sub–centro de la CNA–SAGARPA en Tapachula, Chiapas; ubicada en las mismas isobaras, isoyetas e isotermas del sitio de estudio.
Distribución de la enfermedad en los árboles. Los porcentajes de tejido dañado por antracnosis, en cada una de las 60 ramas, se acumularon y se estandarizaron a una escala con máximo de 1000, considerando como máximo la cantidad de enfermedad acumulada por rama. Esta operación se hizo para las diferentes alturas (1.5, 2.5 y 4.0 m) y en cada uno de los puntos cardinales.
Con estos datos, se calculó: a) La velocidad de crecimiento (r = 1/ t2–t1[Ln(X2/1–X2)–Ln(X1/1–X1)]; Van der Plank, 1963) durante la fase exponencial de la enfermedad, b) El área bajo la curva del progreso de la enfermedad (ABCPE = Σ[(X1 + X2)/ 2](t2–t1)) (Jeger y Viljanen–Rollinson, 2001) y c) La enfermedad máxima acumulada.
Los datos de severidad y la velocidad de crecimiento de la enfermedad por punto cardinal (4) y altura (3), fueron sometidos a un análisis de varianza factorial 4 x 3 (ANOVA) con distribución en bloques al azar. El análisis se hizo con el programa estadístico "Statistica 7", versión para Windows (Stat Soft, 2003).
Desarrollo fenológico del hospedante y su relación con la enfermedad. La formación del tejido nuevo se registró cada semana, se agruparon de acuerdo a las fechas de emisión de hojas nuevas, y se correlacionaron con la cantidad de enfermedad que prevaleció durante su desarrollo. En la fase logarítmica de cada curva de crecimiento se calculó la velocidad de crecimiento y mediante un ANOVA se determinó si había diferencias entre la cantidad de enfermedad observada sobre brotes jóvenes y maduros.
Fluctuación del inóculo y su relación con la enfermedad. Para encontrar la mejor relación entre la incidencia de la enfermedad en el tiempo y la cantidad de conidios capturados, se recorrieron los datos, hacia atrás en el tiempo, de una a cuatro semanas con respecto a los datos de la enfermedad y se hizo un análisis de correlación.
Efecto de los factores climáticos en el desarrollo de la enfermedad. Para detectar las mejores relaciones entre el desarrollo de la enfermedad y los elementos del clima, se recorrieron los datos de las variables climatológicas hacia atrás en el tiempo, de una a cuatro semanas con respecto a los datos de la enfermedad, se hizo un análisis de regresión simple, para seleccionar los elementos que explicaron mejor las variaciones del fenómeno. Por último, se hizo un análisis de regresión múltiple, para definir cuales parámetros fueron los que contribuyeron en mayor grado al desarrollo de la antracnosis. Todos los análisis estadísticos se hicieron con el programa estadístico "Statistica 7" (Stat Soft, 2003).
RESULTADOS
Daños por antracnosis en mango cv. Ataulfo. En brotes jóvenes se presentaron lesiones de antracnosis, < a 1 mm de diámetro, de color café amarillento, que se tornaron a color café oscuras, casi negras y de bordes regulares. Al unirse forman conglomerados de lesiones con aspecto de salpicadura o escurrimiento. Este tipo de lesiones ocasionaron el enrollamiento ó distorsión de las hojas, la muerte del ápice de hojas tiernas y del brote completo. Las hojas tiernas se distorsionan, se ponen de color negro y se secan; quedan adheridas a las ramas por un tiempo y después caen.
En hojas maduras o viejas se observaron lesiones típicas que se produjeron cuando las hojas fueron jóvenes y manchas necróticas de 3 a 4 cm de diámetro, con círculos concéntricos de color café claro y oscuro. Este tipo de lesiones causó un necrosamiento de aproximadamente 25% en la superficie de las hojas, lo que provocó una severa defoliación durante los meses de septiembre a noviembre. Bajo condiciones de alta humedad relativa, se observó crecimiento de micelio en los bordes de las manchas, que invadió el tejido sano incrementando el tamaño de las lesiones (Fig. 2A). Fue común observar exudados de color rosa salmón, sobre las lesiones.

La flor del mango cv. Ataulfo es un tejido muy susceptible al ataque de C. gloeosporioides, puede presentar lesiones en forma de pizca de color café obscuro, sobre el receptáculo, el pedúnculo, el pistilo, los estambres y/ó los sépalos, pero rara vez sobre los pétalos (Fig. 2B, C y D). A pesar de un daño poco evidente a simple vista, causa la muerte repentina de las flores, las cuales caen al formarse un anillo de abscisión a la mitad del pedúnculo. La diferenciación entre flores caídas por muerte fisiológica y la provocada por antracnosis, solo puede establecerse, al colocar flores marchitas sobre hojas jóvenes sanas, e inducir el desarrollo de las lesiones incubándolas en cámara húmeda, o mediante la detección de acérvulos al microscopio estereoscópico. Cuando las flores caídas, quedaron retenidas sobre hojas jóvenes, desarrollaron lesiones típicas de antracnosis.
En el raquis principal de la panícula y en los raquídeos también se observaron lesiones típicas de antracnosis, pero en pocas ocasiones hubo atizonamiento. Esta caída repentina de las flores evita el prendimiento de los frutos.
Cuando la infección ocurre después del prendimiento, los frutos pequeños mostraron síntomas de roña y quebraduras causadas por Colletotrichum y posteriormente cayeron.
Las infecciones en frutos de mayor tamaño, permanecen latentes hasta la maduración. Cuando estos frutos son infectados por escurrimiento de esporas provenientes del raquis, de ramas o de flores marchitas que quedan en contacto con el fruto, se produce un manchado por lagrimeo (tear–stained, Fig. 2E y F).
Distribución de la enfermedad en árboles de mango cv. Ataulfo. El análisis de varianza de enfermedad máxima acumulada (durante 168 días), tasa de la enfermedad (r), y el área bajo la curva del progreso de la enfermedad (ABCPE), reveló que no existen diferencias significativas entre los puntos cardinales, ni entre alturas, hecho que indicó que la enfermedad se encuentra distribuida al azar en la copa de los árboles, y no existe preferencia, ni condiciones microclimáticas que favorezca el desarrollo del patógeno, en algún sitio en particular (Cuadro 1).
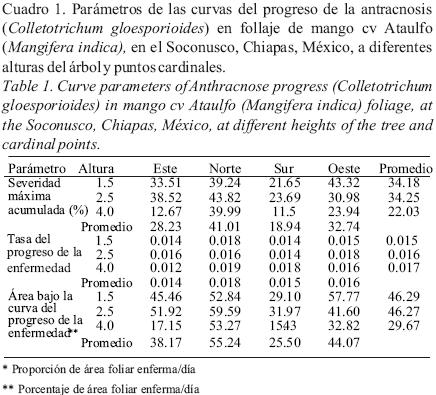
Desarrollo fenológico del hospedante y su relación con la severidad de la enfermedad. Durante el periodo de estudio, solo el 42% de las ramas marcadas emitieron brotes nuevos, que se presentaron en tres flujos vegetativos: El primero se observó del 28 de diciembre al 1 de marzo, el segundo del 31 de mayo al 21 de junio, y el tercero del 9 de agosto al 23 de agosto. De manera general, cuando hubo emisión de brotes, la enfermedad se incrementó de manera significativa. Aun así, la severidad de la enfermedad fue ligeramente mayor sobre brotes maduros que en brotes jóvenes (Cuadro 2), este efecto fue producido por la frecuente caída de hojas jóvenes fuertemente dañadas, mientras que el tejido infectado en brotes maduros permanece. La estimación de la tasa de crecimiento en tejido joven (0.016) fue ligeramente mayor que en brotes maduros (0.015), pero la diferencia no fue significativa. La floración (no representada en la Fig. 3) se inicio en noviembre y finalizó a mediados de febrero del siguiente año. Este hecho indica que hay tejido susceptible de ser infectado por C. gloeosporioides durante todo el año (Fig. 3).
Un análisis de correlación simple entre presencia de brotes jóvenes, maduros y la severidad de la enfermedad, mostró que ambos tipos de brotes explican el 67% de las variaciones del porcentaje de severidad de la enfermedad sobre el follaje (R2= 0.67, P = 0.000 ).
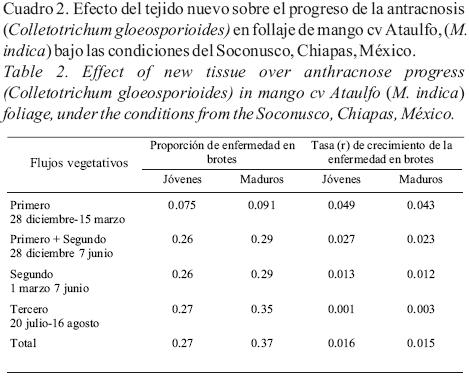
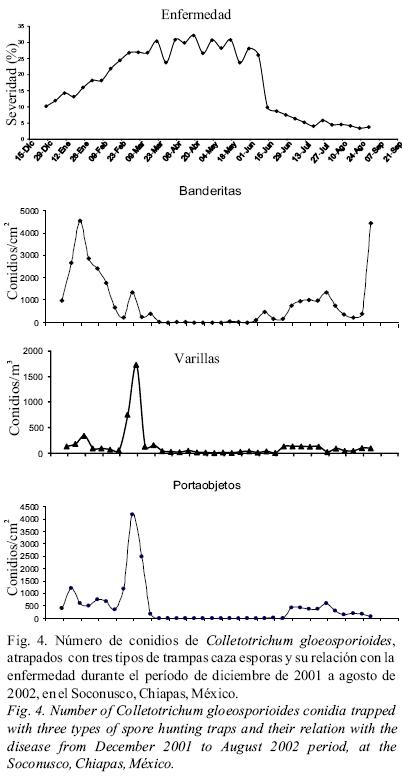
Fluctuación del inóculo y su relación con la antracnosis. El trampeo de esporas con los tres tipos de trampas reflejaron la fluctuación de inóculo durante el año, pero solo en el caso de la trampa "tipo banderitas" se obtuvo una correlación positiva y significativa, que explicó el 23% de las variaciones de la enfermedad (r = 0.48; R2= 0.23, P = 0.001). La presencia de conidios de C. gloeosporioides se detectó desde diciembre (cuando se inició el estudio) hasta la tercera semana del mes de marzo, cuando la enfermedad presentó un crecimiento exponencial (Fig. 4). Durante los meses de abril y mayo no se detectó la presencia de coinidios y coincidió con el período de bajo crecimiento de enfermedad, registrándose una fluctuación entre 25 y 35%. Una vez que las lluvias se establecieron en junio, la presencia de conidios volvió a detectarse, pero la enfermedad tuvo una disminución drástica de 23 a 3%. La cantidad de conidios atrapados con los diferentes tipos de trampa, fue muy disímil. La trampa tipo roto–rod capturó un máximo de 1,722 conidios/m3/hora, el 23 de febrero de 2002; con trampa de portaobjetos 41,819 conidios/cm2/ semana, el 23 de febrero y con las de banderitas 908 conidios/cm2/semana, el 11 de enero.
Elementos del clima y severidad de antracnosis. El análisis de correlación simple entre algunas de las variables climáticas y la severidad de la enfermedad, mostró que entre los parámetros que explican mejor las fluctuaciones de la enfermedad se encuentran, la humedad relativa mínima (HRmin) y máxima (HRmax) presentes 14 días antes de la medición de severidad, con una correlación negativa que explica el 50 % y 49% de las variaciones de severidad observadas (r = –0.71, R2 = 0.50 ), (r = –0.70 y R2= 0.49), y la precipitación pluvial que explica el 25% (r= 0.50 R2= –0.25).
El mayor progreso de la enfermedad se registró cuando la HRmin (observada durante el día) se mantuvo entre 40 y 53% durante los meses de diciembre a marzo (período sin lluvias) y disminuyó cuando esta fue mayor a 55%, de junio a octubre (período con lluvias). Con respecto a la HRmax, el incremento de la severidad coincidió con la presencia de 80–90% de humedad, durante las noches del período sin lluvia y cae drásticamente cuando esta pasa del 90% o es menor de 80% de junio a octubre, período con lluvias (Fig. 5). En esta misma figura se observa que estas dos variables tuvieron un patrón de distribución similar al patrón de precipitación, que consta de 6 meses sin lluvia, de noviembre a inicios de mayo, y 6 meses con lluvia tipo monzón, de mayo a octubre. Además, la reducción drástica de antracnosis que se registró en junio, coincide con la presencia de lluvias y el mayor progreso de la antracnosis con la ausencia de éstas, de diciembre a marzo, período durante el cual se mantuvo la humedad relativa en el ambiente.
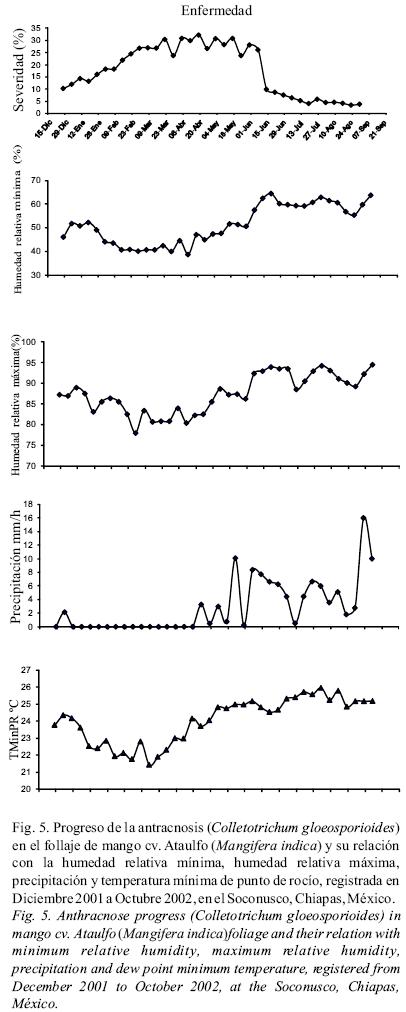
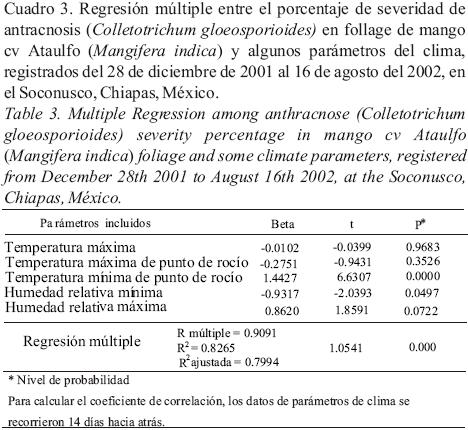
El análisis de regresión múltiple por el método de stepwise, seleccionó después de cinco pasos, la temperatura mínima de punto de rocío (TminPR), correlacionada positivamente con la enfermedad y la humedad relativa mínima (HRmin), con correlación negativa, como las variables más influyentes (Cuadro 2). La ecuación de regresión múltiple generada usando estas dos variables explica el 83% de las variaciones de severidad encontradas. En la figura 5, se puede observar que la fase de crecimiento de la epidemia (representada en % de severidad), coincide con una TminPR entre 21–24°C y una HRmin no menor de 40%, ni mayor de 53%, se mantiene y cae cuando la TminPR fue mayor de 24.5°C y la HRmin fue mayor de 55%.
DISCUSIÓN
Daño por antracnosis en mango cv. Ataulfo. Esta variedad de mango se originó en forma natural en el Soconusco, Chiapas, México, y cuenta con la denominación de origen otorgada por Instituto Mexicano de la Propiedad Industrial (IMPI) y su producción es principalmente para exportación. A pesar de que en los manuales técnicos generados para mejorar la producción del cv. Ataulfo, se menciona el daño causado por el hongo C. gloesporoides en poscosecha, pero no se asoció a este patógeno con el marchitamiento de flores, falta de prendimiento, caída y roña de frutos pequeños, como daño en precosecha (Mosqueda–Vásquez, 1996; Sandoval et al., 1993). En este trabajo se determinó que el daño provocado por la antracnosis es particularmente severo sobre las emisiones de brotes nuevos y durante la floración, cuando coincide con períodos de alta humedad relativa, que se presentan después de finalizar el período de lluvias. La condensación de agua que se produce sobre la superficie de las hojas, durante la noche, hace que las esporas pasen del follaje a las flores, donde el hongo puede causar marchitez y caída de flores, que da como resultado un bajo prendimiento de frutos. Un daño similar fue reportado por Dodd, et al. (1991), al estudiar el efecto de los factores climáticos sobre el desarrollo de la antracnosis de mango cv.Carabao, en Filipinas.
Las infecciones causadas por C. gloeosprioides posteriores al amarre de los frutos, producen lesiones hundidas sobre frutos jóvenes, causando usualmente su caída (Estrada et al., 2000, Dodd et al., 1991). En el caso del mango cv. Ataulfo, las lesiones en frutos pequeños (< 2 cm), son similares a la roña, presentan abundantes acérvulos de Colletotrichum y también caen. El bajo prendimiento de frutos que se presenta en la variedad de mango Ataulfo, en las floraciones inducidas y naturales que se presentaron en los meses de noviembre–diciembre, correspondieron a pérdidas de precosecha, que según Jeffries et al. (1990), causan las especies de Colletotrichum, en frutales desarrollados principalmente en los trópicos.
Efecto del hospedero sobre la enfermedad. Los resultados de este trabajo mostraron que las interacciones entre el hospedero, clima e inóculo, que prevalecieron en las diferentes partes de los árboles de mango, favoreció de manera similar el desarrollo de la enfermedad, por lo que su distribución sobre el follaje fue uniforme. La presencia continua de tejido susceptible (follaje joven y maduro) a través del tiempo, indicó que C. gloeosporioides no tiene limitaciones en cuanto a disponibilidad de tejido hospedante. Este hecho explicó en parte, el incremento de antracnosis conforme aumenta la edad del cultivo y coincidió con lo que se ha reportado por otros investigadores, al estudiar la epidemia de antracnosis en mango cv. Brown's en Alstonville, en Estados Unidos (Fitzell y Peak, 1984; Fitzell et al., 1984). La disminución del porcentaje de antracnosis, durante la emisión de brotes nuevos, fue ocasionada en parte, por la caída de hojas jóvenes con fuerte daño de antracnosis, y porque el rápido incremento de tejido nuevo libre de antracnosis, disminuyó la relación de tejido enfermo/tejido sano. Algo similar fue observado cuando se estudió la epidemia de antracnosis en flores de cítricos (Denham y Waller, 1981).
Cuando la floración del mango cv. Ataulfo es inducida mediante la aplicación de nitrato de potasio durante el mes de octubre, la emisión de yemas florales coincide con la fase de crecimiento acelerado de la enfermedad presente en el follaje joven, en los meses de diciembre– abril.
Cabe señalar que la aplicación de fungicidas cada 15 días, después de haber iniciado la floración, no evita o detiene las infecciones por C. gloeosporioides, lo que induce una falta de prendimiento de fruto severa, en la floración que se produce a finales de diciembre. En las floraciones subsecuentes de fines de enero, febrero y marzo, el daño causado por antracnosis, disminuye gradualmente a medida que la humedad relativa disminuye y la TminPR aumenta, durante febrero y marzo.
Efecto del patógeno sobre la enfermedad. El incremento del número de conidios capturados, durante los meses de diciembre a febrero coincidieron con la ausencia de lluvias torrenciales y con la formación de una mayor cantidad de rocío sobre el follaje de mango, lo cual favoreció la diseminación del hongo por los escurrimientos, salpique y goteo de agua con conidios, dando origen a nuevas lesiones. Durante esta época fue frecuente observar hojas con múltiples lesiones pequeñas de color negro, que adquieren la forma del agua al escurrir. Esto fue también reportado por Jeffries et al. (1990). El ligero incremento en la cantidad de conidios atrapados con los portaobjetos y las varillas, durante el mes de julio, coincidió con el inicio de la temporada de lluvias, lo que se caracterizó por la presencia de lluvias fuertes, aunque discontinuas. Esto último también fue observado por Fitzell y Peak en 1984, al estudiar el desarrollo de la epidemia de antracnosis en mango, en Australia. No obstante, al incrementarse la frecuencia de lluvias torrenciales durante los meses de septiembre y octubre, los conidios fueron lavados. Este efecto produjo una marcada reducción en el número de conidios capturados. Una tendencia similar fue reportada por Madden et al. (1996), en una epidemia de antracnosis en el cultivo de fresa. La reducida cantidad de esporas capturadas de marzo a mayo, pudo estar influenciada por la escasa esporulación del tejido enfermo, y la baja humedad relativa que prevaleció durante este período; es difícil señalar fuentes de inóculo inicial en la epidemia de antracnosis sobre mango, debido a que se producen continuamente nuevas lesiones sobre el tejido susceptible, el cual se forma durante todo el año. Aun así podemos mencionar, que el follaje enfermo es la fuente de inoculo que inicia la epidemia sobre las inflorescencias del mango; y que las primeras flores enfermas que caen, inducen nuevas infecciones al entrar en contacto con flores, hojas y frutos jóvenes. En este sentido, un daño similar fue reportado en aguacate por Fitzell (1987), y en cítricos por Denham y Waller (1981).
Cabe señalar, que durante los meses de julio y agosto se presentó un incremento de antracnosis sobre brotes jóvenes, asociado al daño producido por diabróticas (Coleoptera: Crisomelidae). Este hecho y la abundancia de trips en las floraciones tempranas, hacen suponer que con su actividad, contribuyen a la diseminación de la enfermedad. A pesar de que varios autores han reportado que los insectos promueven la diseminación de C. gloeosporioides, solo Nemeye et al. (1990), demostró que el parasitoide bracónido (Heterospilus prosopidis), parásito de la broca del café (Hypothenemus hampei Ferrari) fue capáz de diseminar los conidios de C. coffeanum y C. gloeosporioides, agentes causales del CBD en café y la antracnosis en mango.
Efecto del clima sobre la antracnosis. Al finalizar la época de lluvias y al inicio del período sin lluvia, la alta humedad relativa (> 80%) que prevaleció en el campo, combinado con valores de TminPR entre 21 a 24°C durante la noche, propició la condensación de agua sobre el follaje de mango, a tal grado que hubo escurrimiento y salpique, que promovió la dispersión de conidios de C. gloeosporioides, aumentando la severidad de antracnosis sobre el follaje, flores y frutos en desarrollo. Durante los meses de marzo, abril y mayo, la humedad relativa media fue de 80%, pero un ligero aumento en TMinPR 24–25°C, disminuyó la cantidad de agua condensada, pero mantiene la diseminación de conidios por arrastre, que produce síntomas de "lagrimeo" sobre hojas jóvenes y frutos en desarrollo. Este resultado coincide con reportes previos de antracnosis en mango, que indican que el salpique de las gotas de lluvia y rocío, así como el arrastre de conidios por gotas de agua, son el principal medio de diseminación de Colletotrichum (Fitzell y Peak 1984; Fitzell et al., 1984; Jeffries et al., 1990). En el Soconusco, Chiapas, la formación de gotas se debió a alta humedad relativa, y a bajas temperaturas durante la noche (18 a 23°C), coincidiendo los resultados de nuestra investigación con lo mencionado por Denham y Waller (1981), quienes señalaron que períodos prolongados de humedad relativa durante la noche, incrementaron la incidencia de antracnosis de 30 a 65% sobre flores de cítricos en Belice. Por otro lado, en presencia de rocío la germinación de conidios aumentó (Wastie, 1972). Sin embargo, son contrarios a otros reportes que mencionan que la lluvia es necesaria para el desarrollo de la antracnosis en mango y que la formación de rocío depende de la hora en que ocurre la lluvia (Fitzell y Peak 1984; Griffiths et al., 1971). En este trabajo se observó que durante la época de precipitaciones fuertes (> 7.5 mm/h), la severidad de la antracnosis sobre el follaje disminuyó, esto parece estar relacionado con el lavado del inoculo que ocasiona la lluvia.
La región del Soconusco, Chiapas tiene un clima cálido húmedo con abundantes lluvias en verano, con un mínimo de 2500 mm al año, llegando en algunos lugares a 5000 mm. La intensidad de la lluvia de esta región es fuerte, produce restricción de visibilidad y el impacto de las salpicaduras sobre una determinada superficie, puede alcanzar varias pulgadas (AIOGDPC, 2003). Durante el desarrollo de este trabajo se observó que al inicio de la época de lluvias (junio y julio), se presentaron lluvias continuas de aproximadamente 0.2 a 30.5 mm/h, que limitaron la dispersión de conidios del hongo; en agosto se presentó un período de 2 a 3 semanas, sin lluvia (denominado periodo de canícula); durante septiembre y octubre se registraron lluvias de mayor intensidad (43.2 mm/h), y de noviembre a mayo deja de llover, aunque hay posibilidad de que se presenten algunas lluvias esporádicas en diciembre y abril.
Agradecimientos. A Juan Manuel D'Amiano–Poumián, por permitir el acceso a la Finca Pumpuapa para realizar esta investigación y a Jesús Vázquez–Aguilar, Jefe del Observatorio del Servicio Meteorológico Nacional, por los datos climatológicos proporcionados. A la Fundación PRODUCE Chiapas A.C., Por el finaciamiento parcial del proyecto "Influencia de la inducción uniforme de brotes vegetativos y reproductivos sobre la producción de mango Ataulfo y la antracnosis". Esta investigación es parte de la tesis de la Maestría en Ciencias en Recursos Naturales y Desarrollo Rural con Orientación en Biotecnología Ambiental del tercer autor, en El Colegio de la Frontera Sur.
LITERATURA CITADA
Acosta–Ramos, M., Noriega–Cantú, D.H., Nieto–Angel, D., y Téliz–Ortiz, D. 2003. Efecto del manejo integrado del mango (Mangifera indica L.) en la incidencia de enfermedades y en la calidad de frutos. Revista Mexicana de Fitopatología 21:46–55. [ Links ]
Allende–Molar, R., García, R.S., y Carrillo, A. 2003. Enfermedades post–cosecha en mangos cultivados en Sinaloa. http://www.ciad.mx/boletin/mayjun02/mangos.pdf (acceso el 10 de junio de 2003). [ Links ]
[AIOGDPC] Asociación Iberoamericana de Organismos Gubernamentales de Defensa y Protección Civil. 2003. Aplicación en red para casos de emergencia (ARCE): Glosario de términos de protección y defensa civil. http://www.proteccioncivil.org/arce/glosario/glosarce–1.htm; Marzo 20, 2003. [ Links ]
Benítez–Camilo, F.A., Huerta–Palacios, G, Holguín–Melendez, F., y Toledo–Arreola, J. 2003. Efecto de Colletotrichum gloeosporioides (Penz.) Penz. y Sacc. en la caída de frutos de mango cv. Ataulfo en el Soconusco, Chiapas, México. Revista Mexicana de Fitopatología 21:223–227. [ Links ]
Dodd, J.C., Estrada, A.B., Matcham, J., Jeffries, P. and Jeger, M.J. 1991. The effect of climatic factors on Colletotrichum gloeosporioides, causal agent of mango anthracnose, in the Philippines. Plant Pathology 40:568–575. [ Links ]
Denham, T.G, and Waller, J.M. 1981. Some epidemiological aspects of post–bloom fruit drop disease (Colletotrichum gloeosporioides) in citrus. Annual Applied Biology 98:65–77. [ Links ]
Estrada, A.B., Dodd, J.C., and Jeffries, P. 2000. Effect of humidity and temperature on conidial germination and appressorium development of two Philippine isolates of the mango anthracnose pathogen Colletotrichum gloeosporioides. Plant Pathology 49:608–618. [ Links ]
Fitzell, R.D. 1987. Epidemiology of anthracnose disease of Avocados. South African Avocado Growers' Association Yearbook 10:113–116. [ Links ]
Fitzell, R.D., and Peak, C.M. 1984. The epidemiology of anthracnose disease of mango: inoculum sources, spore production and dispersal. Annals Applied Biology 104:53–59. [ Links ]
Fitzell, R.D., Peak, C.M., and Darnell, R.E. 1984. A model for estimating infection levels of anthracnose disease of mango. Annals Applied Biology 104:451–458. [ Links ]
García, A.M. 1976. Enfermedades de las plantas en la república Mexicana. Editorial Limusa, México, D.F. 75 p. [ Links ]
García, A.E. 2004. Modificaciones al sistema de clasificación climática de Kópen (Adaptado a las condiciones de la República Mexicana) 5a. Ed. Edit. por la Universidad Nacional Autónoma de México. México, D. F. 90 p. [ Links ]
Griffiths, E., Gibbs, J.N., and Waller, J.M. 1971. Control of coffee berry diseases. Annals of Applied Biology 67:45–74. [ Links ]
Jeger, M.J., and Viljanen–Rollinson, S.L.H. 2001. The use of the area under the disease–progress curve (AUDPC) to assess quantitative disease resistance in crops cultivars. Theoretical Applied Genetic 102:32–40. [ Links ]
Jeffries, P., Dodd, J.C., Jeger, M.J., and Plumbley, R.A. 1990. The biology and control of Colletotrichum species on tropical fruit crops. Plant Pathology 39:343–366. [ Links ]
Madden, L.V., Yang, X., and Wilson, L.L. 1996. Effects of rain intensity on splash dispersal of Colletotrichum acutatum. Phytopathology 86:864–874. [ Links ]
Mosqueda–Vázquez, R., de los Santos–de la Rosa, F., Becerra–Leor, E.N., Cabrera–Mireles, H., Ortega–Zaleta, D.A., y del Angel–Pérez, A.L. 1996. Manual para cultivar mango en la planicie costera del Golfo de México. Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias. Centro de Investigación Regional Golfo Centro. Veracruz, Ver. México. Folleto Técnico No. 15. 131 p. [ Links ]
Nemeye, P.S., Moore, D., and Prior, C. 1990. Potential of parasitoid "Heterospilus prosopidis" (Hymenoptera: Braconidae) as a vector of plant–pathogenic Colletotrichum spp. Annals of Applied Biology 116:11–19. [ Links ]
Pérez, E., Isea, F., Montiel, A., Marin, M., y Sandoval, L. 1999. Efecto de la distancia de siembra y la altura de muestreo en la colección de conidios de hongos en una plantación de guayabo. Revista de la Facultad de Agronomía (LUZ) 16 Supl. 1:43–48. [ Links ]
Ploetz, R.C. 1994. Anthracnose. p. 35–36. In: R.C. Ploetz, GA. Zentmyer, W.T. Nishijima, K.G. Rohrbach and H. D. Ohr (eds). Compendium of tropical fruit diseases. APS Press. The American Phytopathological Society, St. Paul, Minnesota, USA. 88 p. [ Links ]
Prior, C., and Rider, K. 1987. Effect of low volume copper sprays with polyisobutene sticker on mango blossom blight (Glomerella cingulata) in Dominica. Tropical Pest Management 33:350–352. [ Links ]
Prusky, D., Freeman, S., and Dickman, M.B. 2000. Colletotrichum. Host specificity, pathology, and host–pathogen interaction. APS Press, The American Phytopathological Society. St. Paul, Minnesota, USA. 393 p. [ Links ]
Sandoval, E.A., Hernández, O.J., Montecillo, T.J.L., y Quilantán, C.J. 1993. Manual de producción de mango en la costa de Chiapas. INIFAP. Ocozocoautla, Chiapas, México. 21 p. [ Links ]
Spalding, D.H. 1982. Resistance of mango pathogens of fungicides used to control post–harvest diseases. Plant disease 66:1185–1186. [ Links ]
StatSoft, Inc., 2003. STATISTICA (data analysis software system), Version 7. www.statsoft.com. [ Links ]
Van der Plank, J.E., 1963. Plant Diseases: Epidemics and Control. Academic Press, New York, USA. 349 p. [ Links ]
Wastie, R.L. 1972. Secondary leaf fall of Hevea brasiliensis: Factors affecting the production, germination and viability of spores of Colletotrichum gloeosporioides. Annals Applied Biology 72:273–283. [ Links ]
Whitney, P.J. 1976. Microbial Plant Pathology. Hutchinson, Great Britain; London, England. 161 p. [ Links ]
Zadoks, J.C., and Schein, R.D. 1979. Epidemiology and Plant Disease Management. Oxford University Press. New York, USA. 427 p. [ Links ]

