










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de micología
versión impresa ISSN 0187-3180
Rev. Mex. Mic vol.33 Xalapa jun. 2011
Contribuciones
Effect of potassium bicarbonate on fungai growth and sclerotia of Sclerotium cepivorum and its interaction with Trichoderma
Efecto del bicarbonato de potasio en el Sclerotium cepivorum y su interacción fúngico y esclerocios de Trichoderma
Blanca Lilia Ortega–Aguilar, Alejandro Alarcón, Ronald Ferrera–Cerrato
Área de Microbiología, Postgrado de Edafología, Colegio de Postgraduados, Campus Montecillo 56230, Estado de México. México.
Autor para correspondencia:
Alejandro Alarcón
aalarconcp@gmail.com
Recibido 18 de julio 2010;
aceptado 18 de abril 2011.
Resumen
Se estudió el efecto del bicarbonato de potasio (KHCO3) como agente antifúngico en el crecimiento de los hongos Sclerotium cepivorum y Trichoderma cepa R39. Se evaluó el crecimiento del micelio, el número y germinación de esclerocios formados por S. cepivorum, así como el antagonismo de Trichoderma sobre el patógeno, en el medio de cultivo papa dextrosa agar (PDA, Baker®) enriquecido con siete concentraciones de KHCO3 (2, 4, 6, 8, 10, 25 and 50 mM). Con 50 mM de KHCO3, el crecimiento de S. cepivorum fue inhibido en 34.5% y el de Trichoderma en 83.1%. Con la aplicación de 10, 25 y 50 mM de KHCO3, la formación de esclerocios por S. cepivorum fue significativamente inhibida (51.5%, 77.0% y 100%, respectivamente), al igual que la germinación de los esclerocios (18%, 78% y 86%, respectivamente). Al confrontar ambos hongos en el medio de cultivo enriquecido con KHCO3, no se afectó la capacidad antagónica de Trichoderma hacia el patógeno. Después de 144 h, Trichoderma mostró una colonización del 70% sobre el micelio de S. cepivorum, ante 50 mM. Los beneficios potenciales del KHCO3 para inhibir tanto el crecimiento y desarrollo de S. cepivorum fueron evidentes al reducir el crecimiento fúngico, la formación y germinación de esclerocios.
Palabras clave: compuesto antifúngico, antagonismo, fitopatógeno, biocontrol.
Abstract
The effect of potassium bicarbonate (KHCO3) as an antifungal agent was tested on the fungal growth of Sclerotium cepivorum and Trichoderma strain R39. The growth of the fungal colony, the number and germination of sclerotia formed by S. cepivorum, as well as the antagonism of Trichoderma on the pathogen were evaluated in potato dextrose agar (PDA, Baker®) culture media amended with seven concentrations of KHCO (2, 4, 6, 8, 10, 3 25 and 50 mM). At 50 mM, the growth of S. cepivorum was inhibited 34.5%, whilst for Trichoderma inhibition was 83.1%. At the concentrations of 10, 25 and 50 mM of KHCO3 sclerotia formation by S. cepivorum was significantly inhibited (51.5%, 77.0% and 100%, respectively), likewise the sclerotia germination (18%, 78% and 86%, respectively). When both fungi were confronted in a KHCO3 –enriched PDA, this chemical compound did not affect the antagonistic capability of Trichoderma towards the pathogen. After 144 h, Trichoderma showed an invasion of 70% over the colony of S. cepivorum, at 50 mM. The potential benefits of KHCO to inhibit both growth and development of S. cepivorum were 3 evident, since it reduced either fungal growth or both formation and germination of sclerotia.
Keywords: antifungal compound, antagonism, plant pathogen, biocontrol.
Introduction
México at the end of 2010 had 19,225 ha cultivated with onion (Allium cepa L.), and average yield of 15,933 ton ha–1 (SIAP, 2011). However, this crop is affected by white rot, caused by the fungus Sclerotium cepivorum Berk, which spreads rapidly and causes significant crop losses (Pinto et al., 2000; Ponce–Herrera et al., 2008). This fungus forms sclerotia, which may remain in the soil for up to 20 years. The occurrence of the disease is related to the presence and abundance of sclerotia in soil, since these structures remain dormant in the absence of the crop, and its germination is stimulated by root exudates of Allium species, which contain volatile compounds such as allyl sulfide and n–propyl (Entwistle et al., 1982; Tariq and Magee, 1990).
In order to reduce the damage caused by this plant pathogen, several methods have been used, but the most common is the chemical control. However, the excessive use of fungicides has caused pollution problems like leaving residues in the harvested crop, and most important it has also induced pathogen resistance (Fushiwaki et al. 1990; Ma and Michailides, 2005; Klose et al., 2010; Van Dyk and Pletschke, 2011). Therefore, less environmentally harmful control alternatives have been sought. For example, biological control which is based on the use of antagonistic microorganisms and on the use of innocuous chemical compounds, such as bicarbonates (Quiroz–Sarmiento et al., 2008; Ordóñez–Valencia et al., 2009; Ibarra–Medina et al., 2010). The aim of these alternatives is to contribute on soil conservation, as well as to ensure quality food sources for humans and to maintain a healthy environment (Bombelli and Wright, 2006).
Laboratory and greenhouse studies have successfully proven the effects of microbial antagonists on plant pathogens (Arzate–Vega et al., 2006). In this way, species of the genus Trichoderma are efficient and long–lasting agents for controlling pathogenic fungi such as Phytophthora, Sclerotinia, Sclerotium, Rhizoctonia, Fusarium, among others (Tsahouridou and Thanassoulopoulos, 2002; Vinale et al., 2008; Ibarra–Medina et al., 2010), by producing toxic metabolites, inhibiting or parasitizing mycelium of soil–borne fungi (Zago et al., 2001).
On the other hand, bicarbonates are commonly used in food industry to avoid fermentation, control pH, and develop adequate textures or flavors (Karabulut et al., 2001; Bombelli and Wright, 2006). Their efficiency has also been proven in the control of many phytopathogenic fungi (Palmer et al., 1997; Gamagae et al., 2003). The bicarbonate ion has been identified as the probable cause of growth inhibition in some bacteria and fungi. In the case of fungi, hyphal walls collapse in the presence of bicarbonate, then causing either growth inhibition or fungal death (Palmer et al., 1997; Ordoñez–Valencia et al., 2009).
There are reports about the effectiveness of potassium bicarbonate (KHCO3) on the reduction of foliar damage produced by fungi, as well as the benefit of Trichoderma species for controlling soil–borne fungi (Tsahouridou and Thanassoulopoulos, 2002; Vinale et al., 2008). However, the effect of KHCO3 on the growth of S. cepivorum and on its interaction with Trichoderma is unknown. This study evaluated the effect of KHCO3 on the growth and development of S. cepivorum, and on its interaction with Trichoderma strain R39.
Materials and methods
Phase I. Effect of potassium bicarbonate on the mycelial growth of Trichoderma and Sclerotium cepivorum
Cultures of Trichoderma sp. strain R39 and Sclerotium cepivorum were obtained from the Departamento de Microbiologia, Colegio de Postgraduados. Sclerotium cepivorum was isolated from a garlic crop field at Guanajuato, and the Trichoderma strain was selected due to its antagonistic and mycoparasitic ability against a wide range of species of pathogenic fungi of cultivated plants, including S. cepivorum (unpublished data).
Disks of medium and mycelium (0.9 cm of diameter) of either Trichoderma sp. R39 or S. cepivorum were placed at the center of Petri dishes containing Potato Dextrose Agar (PDA, Baker®) enriched with seven concentrations of KHCO3 (2, 4, 6, 8, 10, 25, and 50 mM) whose pH was 7.0, 7.0, 7.1, 7.3, 7.4, 7.9 and 8.0, respectively. Five Petri dishes were used for each concentration of KHCO3. PDA–dishes cultures without KHCO3 were used as control (pH 6.5). Fungal cultures were incubated at room temperature (20°C) for four days, and the fungal growth of each fungus was measured each 24 h. Afterwards, the Petri dishes with S. cepivorum were kept at room temperature (20°C) for 7 days (168 h), to count new–formed sclerotia at each concentration of KHCO3.
A completely randomized experimental design was used for the experiment which included eight treatments for each fungus with five replicates each (n=5). Data were analyzed using analysis of variance and the mean comparison test (Tukey, á=0.05) using SAS statistical program (SAS Institute Inc, 2002).
Phase II. Effect of potassium bicarbonate germination of sclerotia of Sclerotium cepivorum
Sclerotia of S. cepivorum were collected from Petri dishes without KHCO3 from the previous experiment. Sclerotia were surface disinfected with 80% alcohol for 30 seconds, followed by several rinses with sterile distilled water. The sclerotia were transferred to a gentamicine solution (20 |j.g mL–1) and incubated at 4°C for 24 h. Excess of antibiotic solution was removed by blotting dry sclerotia on sterile paper. Ten sclerotia were placed in each Petri dish containing PDA and KHCO3 at the concentrations and pH described in the previous section. Plates were incubated at 20°C, and thus, germinated sclerotia were counted daily during seven days.
A completely randomized experimental design was used for the experiment that included eight treatments with five replicates each (n=5). Data were analyzed using analysis of variance and the mean comparison test (Tukey, α=0.05) using SAS statistical program (SAS Institute Inc, 2002).
Phase III. Effect of potassium bicarbonate on the antagonism in vitro of Trichoderma R39 on Sclerotium cepivorum
Petri dishes containing PDA and KHCO3 at the concentrations and pH indicated in the previous section were inoculated with PDA disks with mycelium. Trichoderma R39 and Sclerotium cepivorum were placed in opposite sides of the Petri dishes. Sclerotium cepivorum was inoculated first, due to its slow growth, and Trichoderma R39 was placed 24 h later. Fungal cultures were incubated at 20 °C and the growth of both fungi was measured every 24 h for four days. The invasion percentage of Trichoderma over the fungal colony of the pathogen was estimated according to the following equation (Rollan et al., 1999): C=[(DCAP/DSP)x100], where: C=Invasion percentage; DCAP=distance covered by the antagonist on the pathogen colony over the axis which separates both fungi, DSP=distance between sowing points (6.5 cm).
A completely randomized experimental design was used for the experiment that included eight treatments with five replicates each (n=5). The data obtained were analyzed using analysis of variance and the mean comparison test (Tukey, α=0.05) using SAS statistical program (SAS Institute Inc, 2002).
Results
Phase I. Effect of potassium bicarbonate on the mycelial growth of Trichoderma and Sclerotium cepivorum
Concentrations of KHCO3 produced gradual increases on the pH in culture media (from 7.0 to 8.0) when compared to the control (pH=6.5). After 24 h, concentrations of 2 and 6 mM of KHCO3, significantly (P<0.001) inhibited the growth of Trichoderma by 20.6% and 50.3%, respectively; the concentrations of 10 mM and 25 mM caused an inhibition of 58.1% and 84.2%, while in 50 mM the growth of the fungal culture was entirely inhibited (Figure 1A). After 96 h, concentrations of 2 and 6 mM inhibited fungal growth by 4.0% and 7.3% while concentrations of 10, 25 and 50 mM caused an inhibition of 15.7%, 17.5 and 67.9%, respectively in comparison to the control (Figure 1A).

After 24 h, concentrations greater than 2 mM significantly (P<0.001) reduced the fungal growth, in comparison to the control (Figure 1B). In contrast, after 96 h, the application of 2, 10 and 25 mM, caused fungal growth inhibition of 1.5%, 5.8% and 6.4%, respectively; however, at 50 mM the fungal growth reduction was 34.5% in comparison with the control (Figure 1B).
KHCO3 caused significant (P<0.001) reductions on the number of sclerotia. After 168 h, fully formed sclerotia were observed in the concentrations from 0 to 10 mM; in contrast, at 25 mM the sclerotia formation just started, while sclerotia did not form at 50 mM (Figure 2). At the concentrations of 2 and 10 mM of KHCO3, the number of sclerotia decreased 9.1% and 51.5% respectively, while at 25 and 50 mM, the inhibition on the number of sclerotia was 77.0% and 100% respectively, in comparison to the control (Figure 2).

Phase II. Effect of potassium bicarbonate germination of sclerotia of Sclerotium cepivorum
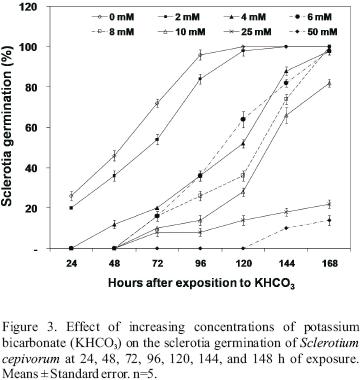
KHCO3 significantly inhibited (P<0.001) the germination of sclerotia. After 24 h, sclerotia germination percentage was 26% and 20% in the control (0 mM) and in the concentration of 2 mM, respectively. After 48 h, sclerotia germination at control, 2 and 4 mM was 46%, 36% and 12% germination, respectively, whilst from concentrations from 6 to 50 mM, germination was not observed (Figure 3). After 168 h, 100% germination of sclerotia was observed for concentrations between 0 and 8 mM, while at 10, 25 and 50 mM, germination was 80%, 22% and 14%, respectively (Figures 3 and 4).
Phase III. Effect of potassium bicarbonate on the antagonism in vitro of Trichoderma R39 on Sclerotium cepivorum
Concentrations of KHCO3 up to 6 mM resulted in significant differences (P<0.001) in the antagonistic capability of Trichoderma towards the pathogen (Figure 5). After 96 h, invasion of Trichoderma on the colony of S. cepivorum at concentrations of 0 to 8 mM was approximately of 58%. At 10 and 25 mM the average percentage invasion of Trichoderma was 45%, while at 50 mM was 30% (Figure 5). After 120 h, the antagonism of Trichoderma on S. cepivorum persisted at concentrations from 0 mM (control) to 25 mM, in which the average invasion percentage was 78% (Figure 5). At 50 mM, the invasion percentage was 53% (Figure 5) although the pathogen showed less growth than Trichoderma (Figure 6). After 144 h, Trichoderma grew over the colony of the pathogen by 96% at concentrations from 0 to 10 mM, while at 25 and 50 mM, the invasion of the antagonist on S. cepivorum was 80% and 70%, respectively (Figure 5).

Discussion
Results show that KHCO3 exerted significant effects on the growth of Trichoderma R39 and S. cepivorum. For Trichoderma R39, the concentration of 50 mM inhibited in 67.9% the fungal growth. In contrast, the growth of S. cepivorum at 50 mM was inhibited in 34.5%. These results denote that Trichoderma was more sensitive to the presence of bicarbonate in comparison to S. cepivorum. This inhibitory effect can be explained partly due to the pH changes that bicarbonate concentrations induced in the culture media, thus affecting the acidic requirements for fungal growth (Steyaert et al., 2010). For instance, Harman and Taylor (1994) indicated that a pH 4.5 was more suitable for the growth of Trichoderma sp. in vitro, whilst at pH of 2.0 and/or 8.0 the fungal growth is slow. The antifungal effect of KHCO3 observed on S. cepivorum is in accordance to findings that show the growth inhibition of S. rolfsii and Botrytis cinerea under in vitro conditions, when bicarbonates were applied at 20 or 40 mM (Palmer et al., 1997; Corral et al., 1998; Bombelli and Wright, 2006).
This study also shows that the number of sclerotia formed by S. cepivorum was significantly inhibited at KHCO3–concentrations higher than 10 mM. Punja and Grogan (1982) and Palmer et al., (1997) found that the carbonates and bicarbonates applied in a concentration of 50 mM inhibited the formation and germination of sclerotia formed by S. rolfsii and B. cinerea. The decrease in the number of sclerotia observed in this study can be related to the pH increase due to KHCO3. In this sense, several findings indicate that the pH affects both growth and sclerotia formation by fungal phytopathogens such as B. cinerea, S. rolfsii, Sclerotinia sclerotiorum and Rhizoctonia solani (De Pasquale and Montville, 1990; Ordóñez–Valencia et al., 2009).
In addition, the present research shows the effects of KHCO3 on the fungal confrontation between Trichoderma sp. and S. cepivorum. The antagonic effect of Trichoderma on S. cepivorum was maintained in all KHCO3–concentrations, but the pathogen showed more growth inhibition at 50 mM. Although the antagonism of Trichoderma species on fungal pathogens is well documented (Vinale et al., 2008), the in vitro confrontation of this antagonist with S. cepivorum under culture media amended with KHCO3 has not been previously reported.
The growth of Trichoderma was more sensitive to 50 mM of KHCO3 than S. cepivorum in single cultures. In contrast, when both fungi were confronted, the growth of S. cepivorum was more limited. This effect can be in part explained due to the fungal antagonism in combination with the increased pH in the culture media by KHCO3. In this regards, Trichoderma species preferably grow up in acid environments than in alkaline ones (Benitez et al., 2004; Steyaert et al., 2010), and their antagonistic capabilities may be influenced by the pH in the culture media (Bell et al., 1982; Vinale et al., 2008). Likewise, the enrichment of culture media with KHCO3 may result in modifications on growth patterns of the fungal colony. For example, Trichoderma at concentrations from 0 to 25 mM had greater sporulation, whereas S. cepivorum, exposed to 50 mM showed less abundance of mycelia, which has finer filaments (data not shown).
Our results agree with those from Tsahouridou and Thanassoulopoulos (2002) and De Souza et al. (2008) in the sense that Trichoderma species have antagonistic effects against pathogens such as S. rolfsii and Moniliophthora perniciosa. There are no earlier reports about the inhibitory effects of KHCO3 on the antagonistic capability of Trichoderma sp. on S. cepivorum, therefore the importance of this study.
An inhibitory effect of KHCO3 on the formation and germination of sclerotia at concentrations up to 10 mM was observed. An 80% inhibition in sclerotia germination of S. rolfsii in the presence of 40 mM of KHCO3 was reported (Ricker and Punja, 1991). This effect shows the potential of bicarbonates to control not only the growth of S. cepivorum but also to inhibit the germination of sclerotia. In this regards, Wilson (1999) showed that carbonates and bicarbonates (sodium and potassium) are effective chemical compounds against B. cinerea which is the causal agent of postharvest diseases in grapes. Likewise, applying 50 mM of ammonium bicarbonate (NH4HCO3) reduced the number of sclerotia en S. rolfsii under in vitro conditions (Punja and Grogan, 1982), while Palmer et al. (1997) indicated that B. cinerea did not produce sclerotia at concentrations greater than 50 mM of KHCO3. The negative effects of bicarbonates on the formation and/or germination of spores have been demonstrated for fungal plant pathogens such as Colletotrichum gloesporiodes, Penicillium spp. (Korsten et al., 2000; Gamagae et al., 2003).
Conclusions
KHCO3 had negative effects on the growth of both S. cepivorum and Trichoderma R39 in vitro. The formation of sclerotia by S. cepivorum was inhibited by 77% and 100% with 25 and 50 mM of KHCO3. Sclerotia germination of S. cepivorum was significantly inhibited as the concentration of bicarbonate in the culture media increased. Potassium bicarbonate had significant effects during the confrontation of both fungi, nevertheless Trichoderma kept its antagonistic effect on S. cepivorum, even at the highest concentration (50 mM).
Acknowledgements
This work was financially supported by the grant SEP–CONACYT 58594. Sincere thanks to the two anonymous reviewers for invaluable comments and thorough revisions.
References
Arzate–Vega, J., A.C. Michel–Aceves, V.M. Domíngez–Márquez, O.A. Santos–Eméstica, 2006. Antagonismo de Trichoderma spp. sobre Mycosphaerella fijiensis Morelet, agente causal de la sigotoka negra del plátano (Musa sp.) in vitro e invernadero. Revista Mexicana de Fitopatología 24:98–104. [ Links ]
Bell, D.K., H.D. Wells, C.R. Markhman, 1982. In vitro antagonism of Trichoderma species against six fungal plant pathogens. Phytopathology 72:379–382. [ Links ]
Benitez, T., A.M. Rincon, M.C. Limón, A. Codón, 2004. Biocontrol mechanism of Trichoderma sp. strains. International Microbiology 7:249–260. [ Links ]
Bombelli, E.C., E.R. Wright, 2006. Tomato fruit quality conservation during post–harvest by application of potassium bicarbonate and its effect on Botrytis cinerea. Ciencia e Investigacion Agraria 33:167–172. [ Links ]
Corral, L.G., L. Post, T.J. Montiville, 1998. Antimicrobial activity of sodium bicarbonate. Journal of Food Science 53:981–982. [ Links ]
De Pasquale, D.A., T.J. Montville, 1990. Mechanism by which ammonium bicarbonate and ammonium sulfate inhibit mycotoxigenic fungi. Applied Environmental Microbiology 56: 3711–3717. [ Links ]
De Souza, J.T., B.A. Bailey, A.W.V. Pomella, E.F. Erbe, C.A. Murphy, H. Bae, P.K. Hebbar, 2008. Colonization of cacao seedlings by Trichoderma stromaticum, a mycoparasite of the witches' broom pathogen, and its influence on plant growth and resistance. Biological Control 46:36–45. [ Links ]
Entwistle, A.R., P.R. Merriman, H.L. Munasinghe, P. Michel, 1982. Diallyldisulphide to reduce the numbers of sclerotia of Sclerotium cepivorum in soil. Soil Biology and Biochemistry, 14:229–232. [ Links ]
Fushiwaki, Y., N. Tase, A. Saeki, K. Urano, 1990. Pollution by the fungicide pentachloronitrobenzene in an intensive farming area in Japan. The Science of the Total Environment 92:55–67. [ Links ]
Gamagae, S.U., S. Sivakumar, R.S.W. Wijeratnam, R.L.C. Wijesundera, 2003. Use of sodium bicarbonate and Candida oleophila to control anthracose in papaya during storage. Crop Protection 22:775–779. [ Links ]
Harman, G.E., A.G. Taylor, 1994. Evaluation of Trichoderma koningii and T. harzianum fom New York soils for biological control of seed rot caused by Phytium spp. Phytopathology 74:107–111. [ Links ]
Ibarra–Medina, V.A., R. Ferrera–Cerrato, A. Alarcón, M.E. Lara–Hernández, J.M. Valdez–Carrasco, 2010. Isolation and screening of Trichoderma strains antagonistic to Sclerotinia sclerotiorum (Lib) de Bary and Sclerotinia minor Jagger. Revista Mexicana de Micologia 31:53–63. [ Links ]
Karabulut, O.A., S. Lurie, S. Droby, 2001. Evaluation of the use of sodium bicarbonate, potassium sorbate, and yeast antagonists for decreasing postharvest decay of sweet cherries. Postharvest Biology and Technology 23: 233–236. [ Links ]
Klose, S., B.M. Wu, H.A. Ajwa, S.T. Koike, K.V. Subbarao, 2010. Reduced efficacy of rovral and botran to control Sclerotinia minor in lettuce production in the Salinas Valley may be related to accelerate fungicide degradation in soil. Crop Protection 29:751–756. [ Links ]
Korsten, L., E.S. De Jager, I. Paul, J. Obagwu, A. Ghaouth, 2000. Alternative control of citrus postharvest diseases. Citriculture 30:77–89. [ Links ]
Ma, Z; T.J. Michailides, 2005. Advances in understanding molecular mechanisms of fungicide resistance and molecular detection of resistant genotypes in phytopathogenic fungi. Crop Protection 24:853–863. [ Links ]
Ordóñez–Valencia, C., A. Alarcón, R. Ferrera–Cerrato, L.V. Hernández–Cuevas, 2009. In vitro antifungal effects of potassium bicarbonate on Trichoderma sp. and Sclerotinia sclerotiorum. Mycoscience 50:380–387. [ Links ]
Palmer, C.L., R.K. Horst, R.W. Langhans, 1997. Use of bicarbonate to inhibit in vitro colony growth of Botrytis cinerea. Plant Disease 81:1432–1438. [ Links ]
Pinto, C.M.F., L.A. Maffia, V.W.D. Casali, R.D. Berger, A.A. Cardoso, 2000. Production components and yield loss of garlic cultivars planted at different times in a field naturally infested with Sclerotium cepivorum. International Journal of Pest Management 46:67–72. [ Links ]
Ponce–Herrera, V., R. Garcia–Espinosa, M. P. Rodriguez–Guzman, E. Zavaleta–Mejia, 2008. Temporal analysis of white rot (Sclerotium cepivorum Berk.) in onion (Allium cepa L.) under three pathogen inoculums densities. Agrociencia 42:71–83. [ Links ]
Punja, Z.K., R.G. Grogan, 1982. Effects on inorganic salts, carbonate –bicarbonate anions, ammonia, and the modifying influence of pH on sclerotial germination of Sclerotium rolfsii. Phytopathology 72:635–639. [ Links ]
Quiroz–Sarmiento, V.F., R. Ferrera–Cerrato, A. Alarcón, M.E. Lara–Hernández, 2008. Antagonismo in vitro de cepas de Aspergillus y Trichoderma hacia hongos filamentosos que afectan al cultivo de ajo. Revista Mexicana de Micología. 26:27–34. [ Links ]
Ricker, M.D., Z.K. Punja, 1991. Infuence of fungicide and chemical salt dip treatments on crater rot caused by Rhizoctonia carotae in long–term storage. Plant Disease 75:470–474. [ Links ]
Rollan, M.C., A.I. Nico, C. Mónaco, 1999. Efecto de la temperatura sobre la interacción in vitro entre especies de Trichoderma y Sclerotinia sclerotiorum, S. minor y S. rolfssi. Investigación Agraria: Producción y Protección Vegetal 14:33–48. [ Links ]
SAS Institute, 2002. The SAS System for Windows, Ver. 9.0. SAS Institute Inc, Cary, N.C. [ Links ]
Steyaert, J.M., R.J. Weld, A. Stewart, 2010. Ambient pH intrinsically influences Trichoderma conidiation and colony morphology. Fungal Biology 114:198–208. [ Links ]
SIAP, 2011. Avence de siembras y cosechas, Otoño–Invierno 2011 (Riego+Temporal). Situación al 31 de enero del 2011. Servicio de Información Agroalimentaria y Pesquera (SIAP). http://www.siap.gob.mx/. Consultada el 14 de marzo del 2011. [ Links ]
Tariq, V.N., A.C. Magee, 1990. Effect ofvolatiles from garlic bulb extract on Fusarium oxysporum f.sp. lycopersici. Mycological Research 94:617–620. [ Links ]
Tsahouridou, P.C., C.C. Thanassoulopoulos, 2002. Proliferation of Trichoderma koningii in the tomato rhizosphere and the suppression of damping–off by Sclerotium rolfsii. Soil Biology and Biochemistry 34:767–776. [ Links ]
Van Dyk, J.S., B. Pletschke, 2011. Review on the enzymes for the detection of organochlorine, organophosphate and carbamate pesticides in the environment. Chemosphere 82:291–307. [ Links ]
Vinale, F., K. Sivasithamparam, E.L. Ghisalberti, R. Marra, S.L. Woo, M. Lorito, 2008. Trichoderma–Plant–Pathogen interactions. Soil Biology and Biochemistry 40:1–10. [ Links ]
Wilson, C.L., 1999. Biological control of postharvest diseases of fruits and vegetables: an emerging technology. Annual Review of Phytopathology 27:425–441. [ Links ]
Zago, E., L.C. Zago, A.C. Ferreira, 2001. Selección de cepas nativas Trichoderma sp. para el control de Sclerotium sclerotiorum in vitro. Ciencia Rural 31:885–887. [ Links ]

