











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
El género Lupinus L. (Leguminosae, Papilionoideae) cuenta con aproximadamente 267 especies (Drummond et al., 2012); la mayoría crecen en América y solo 13 de ellas en tierras altas de África y la cuenca del Mediterráneo. De acuerdo con Atchison et al. (2016) y Kurlovich (2002), desde 2000 años A.C. algunas especies fueron domesticadas para consumo humano y/o animal de forma independiente, tanto en la cuenca del mediterráneo (L. albus L., L. angustifolius L. y L. luteus L.), como en la región de los Andes de América del Sur (L. mutabilis Sweet). La importancia de estas especies se basa en el alto contenido de proteínas en sus semillas (Sujak et al., 2006), así como en su capacidad para obtener nitrógeno atmosférico (N2) por medio del proceso denominado fijación biológica del nitrógeno (FBN). De acuerdo con Weisany et al. (2013) el proceso de FBN que ocurre mediante la asociación simbiótica entre raíces y bacterias nativas del suelo (familia Rhizobiaceae) permite que estas bacterias tomen N del aire y lo transformen en amonio (NH4 +).
Esta simbiosis ocurre mediante la formación de órganos radiculares especializados denominados nódulos, los cuales son colonizados por las bacterias fijadoras de N (Masson-Boivin y Sachs, 2018). En México no se cultivan especies del género Lupinus; sin embargo, se tienen aproximadamente 110 especies nativas distribuidas en varios estados (Bermúdez et al., 2000). Especies como Lupinus montanus Kunth en el Estado de México, así como L. elegans Kunth y L. spp. en Michoacán, fueron utilizadas en programas forestales, ensayos de rehabilitación de suelos y abono verde, respectivamente (Lemus et al., 1986; Lindig-Cisneros et al., 2007; Ramírez-Contreras y Rodríguez-Trejo, 2009). Aunque estos investigadores no realizaron mediciones sobre la cantidad de N que pueden fijar y aportar al suelo, los resultados obtenidos en estos estudios son atribuidos al papel que desempeñan estas plantas en la FBN.
Se han desarrollado diferentes técnicas para cuantificar la FBN: incremento de la producción de materia seca, método de la diferencia de N total, nodulación, solutos en el xilema, actividad enzimática y uso del 15N (Hardason y Danso, 1993). De acuerdo con Radic y McAdam (2012), la técnica de las diferencias del N es una medición confiable para evaluar la fijación. Carlsson y Huss-Danell (2003) señalan que el principio de esta técnica asume que la cantidad de N fijado es igual a la diferencia entre el N total de una planta fijadora y un cultivo de referencia (especie no fijadora de N), sembradas ambas especies en las mismas condiciones. En este sentido, la elección de la planta control o no fijadora de N es de importancia y entre las especies que se han utilizado como cultivo de referencia se encuentran las gramíneas Lolium perenne L., Dactylis glomerata L., Panicum maximum Jacq., Triticum aestivum L. y Zea mays L., entre otras (Boller y Nösberger, 1994; Schulz et al., 1999; Radic y McAdam, 2012; Martín et al., 2017). Aunque la técnica de la dilución isotópica de 15N generalmente es considerada la más confiable para hacer este tipo de investigaciones, de acuerdo con Cueto-Wong et al. (2013), son pocos los estudios que utilizan fertilizantes marcados con 15N, debido a sus altos costos y al de los análisis de las muestras que los contienen.
La información sobre la cantidad de N que fijan las especies cultivadas del género Lupinus es amplia, se han estimado valores de 0 a 463 kg N fijado ha-1, dependiendo de la especie y condiciones edafo-climáticas de los sitios donde se han realizado los experimentos (Schulz et al., 1999; Unkovich et al., 2010); sin embargo, en especies silvestres a nivel mundial estos estudios son escasos. De acuerdo con lo anterior y considerando que en México no se han realizado estudios para cuantificar la fijación de N atmosférico (N2) en especies nativas del género Lupinus, se realizó una investigación con el objetivo de estimar por el método de la diferencia la cantidad de N fijado (kg ha-1) y proporción de N derivado de la atmosfera (% Ndda) en Lupinus exaltatus Zucc., Lupinus rotundiflorus M.E. Jones y Lupinus mexicanus Cerv. ex Lag. en suelos de Zapopan, Jalisco.
Materiales y Métodos
Localización del sitio experimental
Se estableció un experimento de noviembre de 2015 a marzo de 2016 en terrenos del campo agrícola experimental del Centro Universitario de Ciencias Biológicas y Agropecuarias (CUCBA) de la Universidad de Guadalajara, el cual se encuentra en la localidad de las Agujas, Nextipac, municipio Zapopan, Jalisco, México. Las coordenadas geográficas del lugar son 20°43' latitud norte y 103°23' latitud oeste; tiene una altitud de 1600 m. La mayor parte del municipio de Zapopan (85.4%) se caracteriza por su clima semicálido semihúmedo con una precipitación media anual es de 943 mm (INEGI, 2001). La temperatura media anual es de 20.5 °C, mientras que su máxima y mínima promedio oscilan entre 32.1 y 8.4 °C respectivamente (Ruiz et al., 2003).
El suelo del terreno donde se estableció el experimento es clasificado como Regosol, de acuerdo con los análisis físicos y químicos realizados en el laboratorio de análisis de suelos del Centro Universitario de Ciencias Biológicas y Agropecuarias (CUCBA), presenta 51.9% de arena, 20.8% de arcilla, 27.8% de Limo, pH de 5.0, materia orgánica 1.7%, N 0.08%, P 68.52 mg kg-1. El contenido de los cationes Ca, Mg, K y Na fue de 1.36, 0.18, 0.83 y 0.35 cmol kg-1, respectivamente. Durante la época de evaluación solo se registraron datos de temperatura, ya que la precipitación no es común durante esta época.
Material vegetal
Las semillas o germoplasma de Lupinus exaltatus, L. mexicanus y L. rotundiflorus utilizadas en el experimento se recolectaron en los primeros meses del año 2015 en poblaciones silvestres localizadas en el estado de Jalisco (México), en las localidades Ciudad Guzmán (20o15'N, 103º32'O), Lagos de Moreno (21º20'N, 101º54'O) y Chiquilistlán (20º03'N, 103º51'O), respectivamente (Fig. 1). La identificación taxonómica de las plantas se realizó mediante la revisión de bibliografía especializada de flora (McVaugh, 1987) y a través de cotejos con las colecciones del herbario IBUG (números curatoriales 184176, 204527, 37505, 34446, 56754 y 97977).

Establecimiento del experimento en campo
Antes de la siembra, las semillas fueron desinfectadas con hipoclorito de sodio (2%) y se sometieron a un proceso de escarificación (inmersión en ácido sulfúrico concentrado 98% durante 30 minutos), debido a que generalmente las semillas de las especies nativas del género Lupinus presentan una testa endurecida (Medina-Sánchez y Lindig-Cisneros, 2005). Después del tratamiento de escarificación y previa preparación convencional del terreno (barbecho, rastreo y surcado), éste se dividió en tres bloques y cuatro parcelas por bloque de 5 m de ancho × 6 m de longitud (área total por parcela de 30 m2). Las semillas de las tres especies de Lupinus se sembraron el 6 de noviembre de 2015 en forma manual a una profundidad de 3 cm, con una distancia de 15 cm entre plantas y una distancia de 80 cm entre surcos.
Con el propósito de cuantificar la FBN por el método de la diferencia de N también se sembró cebada en forma aleatoria entre las especies del género Lupinus (Hardarson y Danso, 1993; Schulz et al., 1999; Martín et al., 2007; Radic y McAdam, 2012). Se utilizó un diseño experimental de bloques al azar con arreglo factorial de 3 × 3, tres especies, tres muestreos destructivos (93, 108 y 122 días después de la siembra) y tres repeticiones. La densidad de plantación estimada para las especies del género Lupinus fue de aproximadamente 120,000 plantas por ha y 300,000 plantas por ha para la cebada.
Manejo del cultivo
Inmediatamente después de la siembra se realizó un riego por goteo a capacidad de campo mediante cintillas, las cuales tenían una distancia entre goteros de 20 cm. Posteriormente los riegos se realizaron con una frecuencia de 15 días aproximadamente hasta iniciar la formación de vainas. El suelo no fue fertilizado y la baja densidad de malezas que se presentaron durante el crecimiento del cultivo facilitó el control de éstas en forma manual. Cabe señalar que en el periodo de evaluación no se aplicaron pesticidas, debido a que los daños causados por plagas o enfermedades fueron inapreciables.
Muestreos destructivos y cuantificación de materia seca
En cada muestreo se seleccionaron todas las plantas ubicadas en 0.8 m lineales de los surcos centrales, las cuales se cortaron con tijeras de podar a una altura de 3 cm sobre la superficie del suelo. Las plantas colectadas se lavaron y se colocaron en estufa de aire forzado (Novatech, modelo 0200, Tlaquepaque, México) a 70 °C en el laboratorio de Biotecnología del Departamento de Botánica y Zoología (CUCBA) de la Universidad de Guadalajara (UdG) hasta peso constante. Después de registrar el peso seco, las plantas completas se molieron utilizando un molino (RETSCH GMBH, modelo 5657, Haan, Alemania) del Departamento de Madera Celulosa y Papel del Centro Universitario de Ciencias Exactas e Ingenierías (CUCEI) de la UdG para su posterior análisis de nitrógeno. Además, en cada cosecha se extrajo cuidadosamente el sistema radical con el propósito de contar los nódulos por planta.
Análisis de nitrógeno
El contenido de N (%) en la materia seca de las plantas completas incluyendo la cebada se determinó por medio del método Kjeldahl (Bremner y Keeney, 1965). También se analizó el contenido de N en las semillas de cebada y semillas de las diferentes especies de Lupinus utilizadas en la siembra. Cabe señalar que el contenido de N en las semillas se utilizó para estimar posteriormente el porcentaje de nitrógeno obtenido o derivado de la FBN, ya que durante la germinación y establecimiento de plántula constituye la principal fuente inicial de N.
Estimación del % Ndda y cuantificación de N fijado
Se calculó el contenido de N por planta a partir de los datos de la materia seca (g planta-1) y la correspondiente concentración de N (%) mediante la siguientes fórmulas propuestas por Martín et al. (2007) y Radic y McAdam (2012).
Contenido de N (g planta-1) = (materia seca (g. planta-1) × % N total en la planta)/100
Posteriormente se estimó la proporción o porcentaje de N derivado de la atmósfera (% Ndda) en las especies de Lupinus estudiadas utilizando la siguiente fórmula.
Donde:
Contenido N fix= |
Nitrógeno en la planta fijadora (Lupinus) |
Contenido N control= |
Nitrógeno en la planta control (cebada) |
Contenido N semilla fix= |
Nitrógeno en semillas de planta fijadora (Lupinus) |
Contenido N semilla control= |
Nitrógeno en semillas de planta control (cebada) |
Finalmente el nitrógeno fijado se estimó de la siguiente manera
Análisis estadísticos
Las variables número de nódulos, contenido de Ms (kg ha-1), contenido de N (%), porcentaje de nitrógeno derivado de la atmósfera y cantidad de N fijado (kg ha-1) fueron sometidos a un análisis de varianza de acuerdo con el diseño experimental utilizado y con una comparación de promedios mediante la prueba Tukey (0.05), utilizando el programa estadístico Statgraphics Centurión XVII (Statgraphics, 2014).
Resultados
Las variables analizadas mostraron diferencias significativas por efecto de las especies y fecha de muestreo (p<0.05); sin embargo, la interacción especies × fecha de muestreo no fue significativa. En el Cuadro 1 se presenta el crecimiento de las tres especies de Lupinus y planta control (cebada) en términos de acumulación promedio de Ms y concentración de N, así como el contenido de N que presentaban las semillas en el momento de la siembra. Antes de los 93 Dds se observó una lenta acumulación de Ms en todas las especies con exepción de L. exaltatus. Posteriormente la cantidad de Ms se incrementó conforme las plantas alcanzaban la madurez; sin embargo, estos incrementos no fueron de la misma magnitud en todas las especies. En la Figura 2 se muestran las temperaturas promedio que se registraron en Zapopan, Jalisco, durante el desarrollo del experimento. En cada muestreo realizado la producción promedio de Ms tiende a ser más alta en L. exaltatus, mientras que la producción más baja de Ms se registró en L. rotundiflorus (Cuadro 1). Después de realizar el último muestreo, L. exaltatus acumuló 8274 kg ha-1, mientras que L. mexicanus y L. rotundiflorus acumularon 6404 y 2950 kg ha-1, respectivamente.
Cuadro 1: Acumulación de materia seca y contenido de N en tres especies del genero Lupinus L. durante el crecimiento. Promedios seguidos por la misma letra dentro de cada columna indican que no son significativamente diferentes (p≤0.05). Dds=Días despues de la sembra.
| Especies/Dds | Materia seca (g planta -1) | Materia seca (kg ha-1) | N en Materia seca (%) | N en semillas (%) |
| L. exaltatus Zucc. | ||||
| 93 | 29.2 b | 3482 b | 3.5 a | 5.97 |
| 108 | 31.8 b | 3816 b | 3.2 a | |
| 122 | 68.9 a | 8274 a | 2.7 b | |
| L. rotundiflorus M.E Jones | ||||
| 93 | 2.50 c | 300 c | 3.3 b | 6.29 |
| 108 | 7.93 b | 952 b | 4.0 a | |
| 122 | 23.37 a | 2950 a | 3.6 b | |
| L. mexicanus Cerv. ex Lag. | ||||
| 93 | 2.8 c | 345 c | 3.3 a | 5.13 |
| 108 | 14.1 b | 1702 b | 2.6 b | |
| 122 | 49.4 a | 6404 a | 2.5 b | |
| Cebada | ||||
| 93 | 5.2 | 1560 | 1.08 | 1.74 |
| 108 | 9.4 | 2820 | 1.25 | |
| 122 | 8.9 | 2670 | 1.25 |

Figura 2: Temperaturas máximas, mínimas y promedio registradas durante el crecimiento de Lupinus exaltatus Zucc., L. rotundiflorus M.E. Jones y L. mexicanus Cerv. ex Lag. Datos obtenidos de la estación meteorológica localizada en la base aérea militar n° 5 de Zapopan, Jalisco, México.
En la especie utilizada como referencia, la concentración de N varió de 1.08 a 1.25%. Sin embargo, en las diferentes especies de Lupinus, la concentración de N fluctuó de 2.5 a 4% dependiendo de la edad de las plantas (Dds). En L. exaltatus y L. mexicanus se observó una tendencia a disminuir la concentración de N conforme se incrementó la edad de las plantas. Con respecto al número promedio de nódulos por planta, también se observó un incremento significativo de éstos conforme las plantas alcanzaban la madurez (Fig. 3). Nuevamente, L. exaltatus se caracterizó por presentar en cada uno de los muestreos realizados el mayor número de nódulos. La cantidad de N fijado y % Ndda mostraron diferencias significativas por efecto de las especies y periodos de muestreo (Dds). En el Cuadro 2 se muestran los valores promedio N fijado y % Ndda por especie y muestreo, los cuales fluctuaron de 0 a 187.0 kg N ha-1 y de 0 a 83.8% respectivaente. A los 93 Dds la cantidad promedio de N fijado en L. exaltatus fue de 87.4 kg ha-1 equivalente a 71% de Ndda, mientras que en L. mexicanus y L. rotundiflorus, la fijación de N fue inapreciable o nula, lo cual puede estar relacionado a una menor nodulación (Fig. 3) y en consecuencia una pobre acumulación de Ms en estas especies en comparación a L. exaltatus (Cuadro 1).
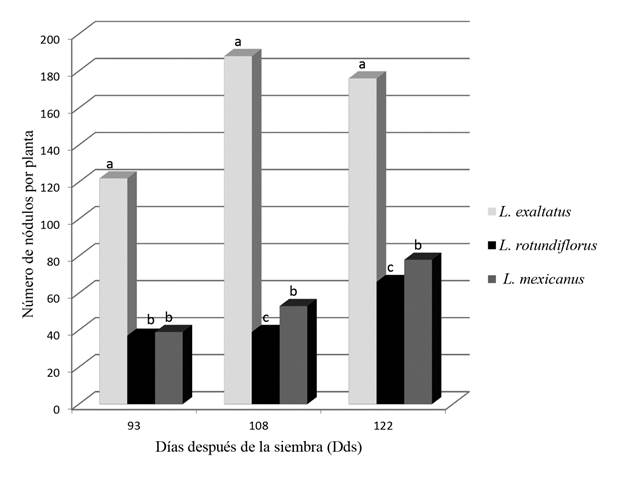
Figura 3: Número de nódulos durante el crecimiento de tres especies silvestres del género Lupinus L. en suelos de Zapopan, Jalisco, México.
A los 108 Dds, en L. mexicanus y L exaltatus se observó un balance positivo respecto a la FBN, pero con baja cantidad N fijado y baja proporción de Ndda (Cuadro 2). Sin embargo, en L. exaltatus, la cantidad de N fijado y el porcentaje de Ndda fueron significativamente similares a los que se registraron en el muestreo anterior (87.4 kg ha-1 y 71% vs 86.9 kg ha-1 y 70.9% respectivamente). En el último muestreo realizado (122 Dds) se registraron los porcentajes más altos de Ndda y cantidad N fijado, con valores significativamente diferentes (p<0.05) entre las especies estudiadas. Los % de Ndda fueron más altos en L. exaltatus y L. mexicanus que en L. rotundiflorus con 83.8, 76.4 y 64.5%, lo cual corresponde a una cantidad de N fijado de 187, 113.62 y 65.6 kg ha-1, respectivamente.
Cuadro 2: Porcentaje de Nitrógeno derivado de la atmósfera (Ndda) y N fijado en Lupinus exaltatus Zucc., L. mexicanus Cerv. ex Lag. y L. rotundiflorus M.E. Jones sembrados en suelos de Zapopan, Jalisco, México. Promedios seguidos por la misma letra dentro de cada columna indica que no son significativamente diferentes (p≤0.05).
| Especie/Dds | Contenido de N (g planta -1) | Contenido N total (kg ha-1) | Fijación Biológica del Nitrógeno | |
| Ndda (%) | N fijado (kg ha-1) | |||
| L. exaltatus Zucc. | ||||
| 93 | 1.02 | 122.6 | 71.0 b | 87.4 b |
| 108 | 1.01 | 122.1 | 70.9 b | 86.9 b |
| 122 | 1.86 | 223.2 | 83.8 a | 187.0 a |
| L. rotundiflorus M.E. Jones | ||||
| 93 | 0.08 | 9.91 | 0.0 c | 0.0 c |
| 108 | 0.31 | 37.2 | 2.85 b | 2.01 b |
| 122 | 0.84 | 100.8 | 64.54 a | 65.6 b |
| L. mexicanus Cerv. ex Lag. | ||||
| 93 | 0.09 | 10.8 | 0.0 c | 0.0 c |
| 108 | 0.36 | 43.2 | 7.18 b | 2.75 b |
| 122 | 1.23 | 148.2 | 76.4 a | 113.62 a |
| Cebada | ||||
| 93 | 0.04 | 12.80 | ||
| 108 | 0.11 | 35.20 | ||
| 122 | 0.11 | 35.20 | ||
Discusión
Únicamente L. exaltatus registró un rendimiento de Ms acumulada superior o igual al que se ha registrado en otras especies domesticadas o silvestres del género Lupinus (Kitessa, 1992; Romero, 1993; Björnsson, 2007; Prusiński, 2014; Zamora et al., 2017). La mayor acumulación de Ms en L. exaltatus indica probablemente una mejor adaptación a los suelos ácidos, arenosos e infértiles de Zapopan, Jalisco, así como a una adecuada adaptación a las condiciones de temperatura registradas durante el estudio, ya que el agua no fue un factor limitante. Los valores de N encontrados en la materia seca de las tres especies en estudio se encuentran dentro de los rangos que se han reportado en otras especies del género Lupinus, tanto domesticadas como silvestres (Kitessa, 1992; Björnsson, 2007; McNeill y Fillery, 2008; Prusiński, 2014).
La declinación del N en L. exaltatus y L. mexicanus conforme se incrementó la edad de las plantas es un comportamiento que ha sido reportado en varias especies no leguminosas y leguminosas, incluyendo especies cultivadas del género Lupinus como L. luteus, L. albus y L. mutabilis (Romero et al., 1993; Plénet y Lemaire, 1999; Barbazán et al., 2002; Pérez et al., 2004; Prusiński, 2014). Sin embargo, el tiempo en que da inicio el descenso de la concentración de N en función del estado de madurez ha sido variable en diferentes especies estudiadas (Gastal y Lemaire, 2002; Prusiński, 2014).
La nula fijación de N en las primeras etapas del crecimiento, particularmente en L. rotundiflorus y L. mexicanus es una característica que también fue registrada en Lupinus albus y L. mutabilis (Schulz et al., 1999), así como en otras especies de leguminosas utilizando el mismo método que se aplicó en esta investigación (Sanginga et al., 1996; Unkovich y Pate, 2000). Lo anterior indica que el N total acumulado en los tejidos de L. rotundiflorus y L. mexicanus durante los 93 Dds fue menor al que se registró en la especie de referencia (cebada). Por lo tanto, es posible señalar que el bajo contenido de N total acumulado en estas especies durante las etapas iniciales del crecimiento fue obtenido directamente del suelo y probablemente de la remobilización del nitrógeno almacenado como reserva en los cotiledones de las semillas utilizadas en la siembra. Lo anterior fue tambien reportado por Ayisi et al. (1992) al evaluar la fijación de N atmosférico en otras especies de Lupinus cultivadas. Los porcentajes de Ndda obtenidos en el último muestreo (122 Dds) son similares a los que se registraron en las primeras investigaciones realizadas en especies del género Lupinus. Por ejemplo en Australia, Unkovich et al. (1997) reportaron en L. angustifolius un valor de 77% de Ndda, mientras que en Islandia, Pálmason et al. (1992), mediante técnicas que involucran 15N, mencionaron en L. angustifolius un porcentaje de Ndda de 90% a los 177 Dds, el cual es superior a los valores de % Ndda obtenidos con las especies del presente estudio. Posteriormente en Chile, Barrientos et al. (2002), en L. angustifolius y L. albus a los 160 Dds registraron en promedio 80% de Ndda, similar al que se registró en este estudio con L. exaltatus (83%). Con respecto a las especies silvestres, los porcentajes de Ndda obtenidos en este estudio son inferiores a los reportados por Pálmason et al. (2002) en L. nootkatensis Donn ex Sims, ya que el % Ndda varió durante cinco años de 97.6 a 99.6%. Sin embargo, los valores de N fijado (80-90 kg ha-1) fueron muy similares o inferiores a los obtenidos en esta investigación con L. exaltatus y L. mexicanus en función de la fecha de muestreo. El mismo patrón se registró en L. arboreus Sims, ya que Baker et al. (1986), aunque no cuantificaron el % de Ndda, reportaron una cantidad de N fijado de 80 kg ha-1 anualmente.
Las variaciones observadas entre los resultados obtenidos en este estudio y los resultados de otras investigaciones con respecto al % Ndda y cantidad de nitrógeno fijado pueden ser explicadas en términos de diferencias con respecto a la especie estudiada, fecha de muestreo, condiciones climáticas y edáficas durante la investigación, así como la metodología utilizada para cuantificar la fijación de N atmosférico y especie de referencia utilizada (Herridge, 1982; Duthion et al., 1987; Evans et al., 1989; Larson et al., 1989; Pálmason et al., 1992; Barrientos et al., 2002). Es probable que los altos valores de materia seca y N fijado en L. exaltatus en comparación con L. mexicanus y L. rotundiflorus estén relacionados a una eficiente simbiosis entre las raíces de esta especie y bacterias nativas del suelo fijadoras de N atmosférico, ya que de las tres especies en estudio, L. exaltatus fue la que mostró mayor número de nódulos bacterianos en la raíces desde las primeras etapas del crecimiento.
Los resultados obtenidos en la presente investigación representan las primeras estimaciones sobre la proporción de N en la planta que proviene de la atmósfera y cantidad de N que fijan algunas especies silvestres del género Lupinus en México. Aunque algunos autores consideran que el método de la diferencia no es el más confiable para estudiar la FBN (Unkovich y Pate, 2000), en estudios con diferentes leguminosas donde se han comparado la técnica de la dilución isotópica de 15N y el método de la diferencia de N, no se han encontrado diferencias significativas en la cantidad de N fijado (Khol et al., 1980; Martín et al., 2017). Por cada tonelada de Ms producida en el último muestreo se estimó que las especies en estudio fijaron aproximadamente 20 kg de N ha-1, lo cual coincide con lo reportado por Unkovich et al. (2010), quienes, mediante modelos lineales para relacionar la producción de Ms con la cantidad de nitrógeno fijado en varias especies de leguminosas (forrajeras y de grano), reportaron valores de 18.7 a 24.3 kg N fijado por tonelada de materia seca cosechada. Lo anterior indica la importancia que pueden tener en un futuro las especies estudiadas como fuente de N para otros cultivos después de su incorporación al suelo en etapa de floración como abono verde.
Conclusiones
Las tres especies en estudio mostraron habilidad para acumular materia seca y fijar N atmosférico en las condiciones en las que se desarrolló el experimento. La especie con mayor potencial de adaptación a estas condiciones en términos de mayor número de nódulos por planta, mayor acumulación de Ms y mayor cantidad de N fijado fue L. exaltatus. La técnica utilizada en este estudio es confiable para obtener las primeras estimaciones sobre la cantidad de N que potencialmente pueden obtener de la atmósfera y fijar diferentes especies del género Lupinus.

