











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
En las montañas del noreste de México existen especies que son endémicas de la región y debido a las condiciones climáticas particulares se mantienen en ecosistemas con alto grado de fragmentación (Treviño-Garza, 2017). Para su conservación y recuperación es necesario contar con información referente a los lugares que reúnen las condiciones ideales para su desarrollo, con la finalidad de diseñar políticas públicas o programas de manejo que mitiguen su deterioro.
Los ecosistemas forestales poseen una compleja estructura y son el hábitat para muchas especies de flora y fauna (Gao et al., 2015). La variabilidad de la vegetación es atribuida a ciertos requerimientos y condiciones ecológicas que determinan la distribución y biodiversidad de cada especie. Algunas especies se desarrollan en rangos geográficos restringidos y hábitats específicos y son propensas a estar en peligro bajo condiciones ambientales cambiantes (Yilmaz et al., 2017). Esas condiciones están influenciadas principalmente por factores climáticos, antropogénicos y topográficos (Pardo et al., 2018).
Las características topográficas como el gradiente altitudinal, la exposición y el relieve superficial son los factores más importantes que determinan las condiciones para el crecimiento y la distribución de las especies (Wang et al., 2015). En este sentido, evaluar el estado actual de la distribución a nivel de especie es de gran interés, debido a los impactos directos e indirectos que sufren los ecosistemas como son el crecimiento demográfico, las plagas, el pastoreo excesivo, la degradación de los bosques naturales por extracción selectiva y la destrucción de estas por el cambio de uso de suelo (Challenger et al., 2009; Zari, 2014). Utilizando técnicas de análisis espacial es posible modelar la distribución potencial de una especie y obtener información de referencia para monitorear la vegetación en el espacio y tiempo, así como para identificar áreas prioritarias en el contexto de la planificación para la conservación (Sáenz-Romero et al., 2010; Henle et al., 2013).
Estos modelos se centran en especies o comunidades con necesidades de restauración, conservación, con distribución restringida y en áreas de alta riqueza de especies frágiles o endémicas (Suárez-Mota et al., 2018; Manzanilla-Quiñones et al., 2019). Por ejemplo, Sáenz-Romero y colaboradores (2012) determinaron la distribución potencial de Abies religiosa (Kunth) Schltdl. & Cham., considerando su importancia como refugio de la mariposa monarca y las afectaciones ante distintos escenarios de cambio climático, como un elemento para apoyar la planificación de la conservación de los sitios de hibernación de la mariposa en México.
Existen diferentes herramientas que apoyan esta planificación. Los modelos de distribución de especies (MDE) se basan en algoritmos poderosos y capaces de predecir la presencia de una especie en función de variables ambientales (Elith y Leathwick, 2009), pueden proporcionar la distribución potencial de las especies y representarlas en mapas a diferentes escalas, basadas en correlaciones entre los registros conocidos de presencia y sus condiciones ambientales (Phillips et al., 2006).
Se han desarrollado diferentes técnicas basadas en algoritmos para modelar la distribución de especies (Kozak et al., 2008; Kass et al., 2018; Singh et al., 2018). El más ampliamente aceptado y utilizado es el de máxima entropía (Maxent) (Phillips et al., 2006; Radosavljevic y Anderson, 2014). Este algoritmo calcula la probabilidad de áreas idóneas, sujeta a un conjunto de restricciones basadas en nuestro conocimiento de las condiciones ambientales en los sitios de presencia de una especie (Elith et al., 2011; Gomes et al., 2018).
En esta investigación se aplicaron MDE para el oyamel (Abies vejarii Martínez), árbol endémico del noreste de México sobreviviente de la última máxima glaciación, catalogado como Casi Amenazado con población decreciente (SEMARNAT, 2010; Farjon, 2013). Los objetivos fueron: i) buscar áreas potenciales para predecir la distribución geográfica potencial del oyamel, utilizando el algoritmo de máxima entropía en la Sierra Madre Oriental (SMOR), ii) identificar las variables ambientales topográficas asociadas a su distribución actual y iii) evaluar los impactos antropogénicos actuales en la distribución de A. vejarii.
Materiales y Métodos
Área de estudio
El estudio se realizó en los bosques templados de la SMOR, provincia fisiográfica que representa a la cadena de montañas más elevada del noreste de México (Chávez-Cabello, 2016) y se sitúa entre las coordenadas 19 º30' y 25 º30' latitud norte y los 97 º30' y 102 º30' longitud oeste (Fig. 1). El intervalo altitudinal de esta cadena montañosa varía de 2000 a 3721 m, su temperatura medio anual oscila entre 7.8 y 25.1 °C, y la precipitación media anual es de 800 a 1200 mm (Vidal-Zepeda, 1990a; 1990b). En ella se pueden encontrar ecosistemas fragmentados que se consideran relictos de los géneros de distribución boreal que son de gran importancia ecológica (Abies Mill., Picea A. Dietr. y Pseudotsuga Carrière) (Treviño-Garza, 2017; González-Cubas et al., 2018).
Obtención de datos
Para formar la base de datos de presencia de A. vejarii a la largo de la SMOR, se usaron tres tipos de registro: 1) muestreo dirigido en campo en el año 2017, se obtuvieron diez rodales naturales con sus respectivas variables dasométricas y de ubicación, 2) ocho muestras botánicas alojadas en los herbarios de la Facultad de Ciencias Forestales de la Universidad Autónoma de Nuevo León (FCF-UANL) y del Instituto de Ecología Aplicada de la Universidad Autónoma de Tamaulipas (UAT), y 3) diez registros mediante revisiones bibliográficas y consulta de la base de datos Global Biodiversity Information Facility (GBIF, 2019).
Depuración de datos
Los datos fueron depurados al dejar un solo registro a una distancia mínima entre sitios de 1 km2. Además, se descartaron aquellos con ubicación imprecisa, con duplicación en la base de datos y algunos puntos donde el registro se observó en áreas sin vegetación natural. Este procedimiento ayudó a evitar el efecto de la multicolinearidad y a maximizar la contribución de las variables (Peterson y Nakazawa, 2008). Se obtuvo una base con 28 registros.
Variables ambientales y topográficas
De los datos ambientales se utilizaron 15 de las 19 variables bioclimáticas de WorldClim versión 2.0 a una resolución de 30 segundos de arco (1 km2) para el período 1970-2000 (Fick y Hijmans, 2017), excluyendo cuatro capas de menor correlación (coeficiente de correlación Pearson ≥0.85) y que combinan información de precipitación y temperatura para evitar anomalías espaciales (Escobar et al., 2014). Las variables topográficas se obtuvieron a partir del Continuo de Elevaciones Mexicano (CEM 3.0) (INEGI, 2018) con coordenadas geográficas de latitud y longitud con una resolución de 1 km2 para que coincida con las fuentes de WorldClim. Se generaron las variables de exposición (°) y pendiente (%). En total se obtuvieron 18 variables de diferentes bases de datos.
Modelado de distribución potencial
Previo a la modelación de la distribución potencial de una especie se recomienda realizar algunos análisis de autocorrelación espacial para reducir errores en el modelo (Cruz-Cárdenas et al., 2014). En este caso se aplicó una prueba de aleatoriedad a los registros espaciales de la especie mediante la función G (Distance to the Nearest Event); esta función mide la distribución de las distancias desde un evento arbitrario a su evento más cercano (Bivand et al., 2013). El valor de la función G se obtiene por la siguiente fórmula:
Donde λ representa la cantidad media de eventos por unidad de área (o intensidad) y r es la distancia n para un evento elegido al azar. Una vez determinada la aleatoriedad de los datos se procedió a ejecutar el algoritmo de Maxent utilizando los parámetros por defecto (lineales, cuadráticas, de producto, de umbral y de bisagra), con un formato de salida logística, con las funciones recomendadas de convergencia (0.00001), el número máximo de iteraciones (500) y el número máximo de puntos (10,000) (Phillips et al., 2006). Maxent generó predicciones de probabilidad de presencia en A. vejarii en formato ráster de cuadrículas con valor de rango 0-1. Por defecto, el algoritmo supone que valores bajos indican escenarios no aptos para el hábitat de la especie, mientras que valores superiores a 0.5 indican que las condiciones son adecuadas (Phillips y Dudík, 2008).
Se usó 75% de los registros (presencia) para modelar la distribución potencial de A. vejarii (Phillips et al., 2006). El 25% restante fue utilizado para la validación del modelo mediante dos procedimientos: (1) por su desempeño y ajuste estadístico en las pruebas del área bajo la curva (AUC, por sus siglas en inglés); esta predicción de significancia se realizó mediante 10 pruebas de omisión generadas con una confiabilidad de 99%, que se obtiene de la técnica ROC parcial (Receiver Operating Characteristic) como un análisis de precisión adicional considerando ausencia y presencia (Anderson et al., 2003; Peterson et al., 2008), con el uso de la plataforma NicheToolBox de la CONABIO (2019); (2) mediante salidas a campo para realizar la evaluación de la distribución de A. vejarii, registrando la presencia o ausencia en el área del modelo.
Se consideró como criterio los valores superiores a 0.6 que indican la probabilidad de la presencia de la especie, misma que aumenta al acercarse al valor de 1. Considerando lo anterior se generaron tres capas utilizando los rangos de valores obtenidos: de 0.6 a 0.7 para indicar una probabilidad baja de presencia de la especie, de 0.7 a 0.8 para indicar una probabilidad media y de 0.8 a 1 para una alta. Para la presentación final se sumaron las tres capas, se generó una sola con las calidades de hábitat.
Factores ambientales y topográficos
Para identificar las variables ambientales y topográficas de mayor influencia para cada una de las localidades de presencia, se empleó un análisis estadístico multivariado de grupos (análisis clúster, CA por sus siglas en inglés) y de componentes principales (PCA por sus siglas en inglés). El CA es una técnica para encontrar características similares de los elementos analizados utilizando un método de agrupamiento de Ward con una distanciación euclidiana (Ward, 1963; Maechler et al., 2019). El PCA es una herramienta de ordenación para reducir la dimensionalidad y determinar la importancia de cada una de las principales variables (Tabachnick y Fidell, 2007; Magyar et al., 2013).
La importancia de cada eje de PCA se verificó comparando sus respectivos valores observados en relación con una distribución. Todos los análisis se realizaron con el software estadístico R project (R Core Team, 2019), junto con los paquetes de spatstat (Baddeley y Turner, 2005), maptools (Bivand y Lewin-Koh, 2013) y lattice (Sarkar, 2008).
Índice de impacto antropogénico
Con base en el levantamiento de información en campo, se determinó el impacto humano considerando los diferentes niveles de perturbación observados en los ecosistemas. Para obtener una medida cuantitativa, se calculó un índice que oscila entre 1 y 6 para cada sitio de muestreo con los siguientes criterios: 1, sin perturbación; 2, con actividades de pastoreo; 3, con aprovechamiento forestal; 4, presencia de daño al arbolado; 5, evidencia de incendios y 6, extracción ilegal. Este factor de intensidad (1, 2, 3, 4, 5 o 6) se correlacionó con la distancia de ubicación a un asentamiento humano más cercano.
La distancia se derivó a partir de un análisis espacial determinada por las coordenadas UTM (en inglés Universal Transverse Mercator) de los sitios de muestreo y la ubicación de un asentamiento humano más próximo, donde se efectuó el cálculo de una distancia (D) lineal entre dos puntos por la siguiente fórmula (Sarría, 2006):
Donde x 2-x 1 es la diferencia entre las coordenadas x de los puntos y y 2-y 1 es la diferencia entre las coordenadas y de los puntos.
Finalmente, para evaluar la importancia relativa de los factores climáticos, topográficos y antropogénicos como determinantes de la variación geográfica de las poblaciones de A. vejarii, se empleó un modelo lineal. Las variables de predicción incluyeron la idoneidad climática obtenida a partir del ejercicio de modelado de nicho, el impacto humano y las variables topográficas. Todos los modelos lineales se obtuvieron mediante el software estadístico R Project (R Core Team, 2019).
Resultados
Los resultados de la prueba de aleatoriedad aplicada a los registros para A. vejarii indican que los registros muestran patrones de una agregación regular, ya que los valores observados de la línea continua se distribuyen hacia arriba y fuera de las bandas de confianza, porque la probabilidad de encontrar un registro es menor (G(r)=0.55) a distancias de aproximadamente 50 km (<0.45).
Modelo de distribución potencial
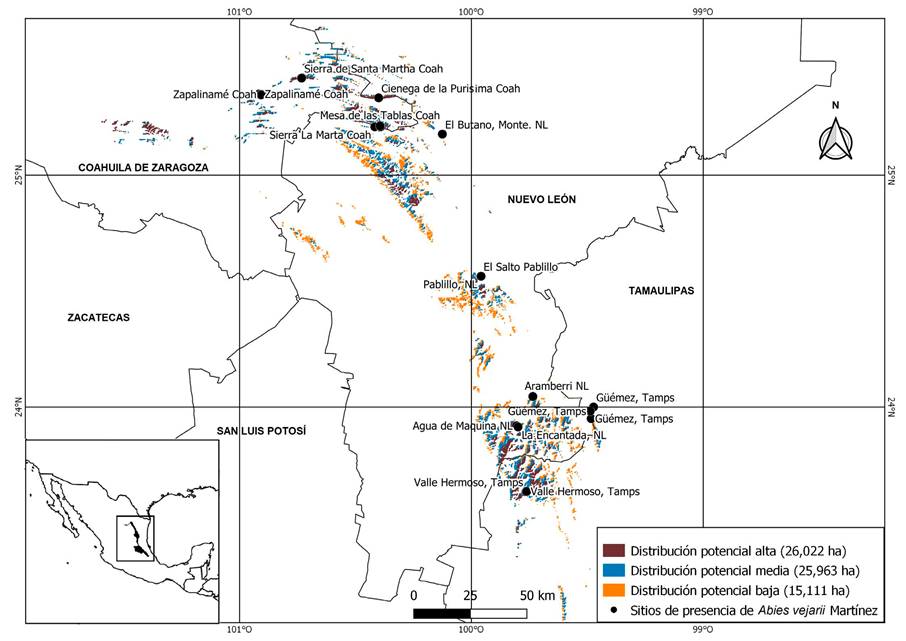
Se ubicaron 28 poblaciones de A. vejarii distribuidas en tres estados de México, todas en la Sierra Madre Oriental. Diecisiete se encuentran en el estado de Coahuila, distribuidos en la parte alta del área natural protegida de la Sierra de Zapalinamé y a lo largo de la Sierra de Arteaga. En el estado de Nuevo León fueron encontradas seis poblaciones, todas en el sur del estado. Para el estado de Tamaulipas, solamente cuatro áreas fueron ubicadas en los bosques templados del municipio Miquihuana.
La validación del modelado potencial de la especie, mediante las 10 pruebas binomiales de omisión usadas en la prueba de ROC fue significativa (p<0.01). Se observó un buen rendimiento en el desarrollo y evaluación del modelo mediante los valores de AUC=0.998, ya que los valores de AUC fueron superiores a 0.9 y de acuerdo con Swets (1988), el modelo desarrollado se puede clasificar como altamente preciso. Cuando se aplicó la prueba de significancia (frecuencia AUCratio>1) al ROC parcial para el modelo, se presentó un valor superior a 1 (1.997). Por lo tanto, se concluye que el modelo es mejor que lo aleatorio.
El mapa que muestra la distribución potencial estimada resultante del ejercicio de modelado y las presencias actuales para A. vejarii se muestra en la Figura 2. Usando una escala de tres colores, el marrón indica la presencia potencial más alta de la especie, mientras que el naranja es la más baja. El área con mayor potencial se ubica en la Sierra de Arteaga, Coahuila, seguida por el municipio General Zaragoza, Nuevo León; mientras que los niveles más bajos se observaron en la zona centro de la SMOR.
Contribución de variables ambientales y topográficas
Con base en CA, los 28 sitios de distribución se separaron en seis grupos principales. Las divisiones se reconocen de acuerdo con su altitud. La primera y la cuarta división fueron sitios distribuidos entre 3000 y 3200 m s.n.m. La segunda y tercera división conforman un bosque con altitudes de 2600-3000 m s.n.m., y la quinta y sexta división, con nueve sitios, se distribuyen en altitudes menores a 2600 m (Fig. 3).
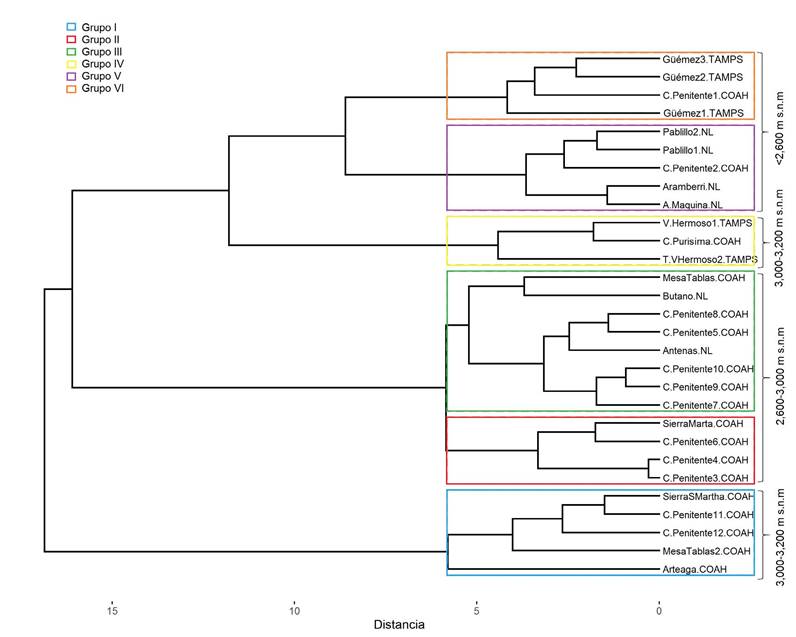
Figura 3: Dendrograma con los principales sitios de distribución asociados con variables ambientales y topográficos.
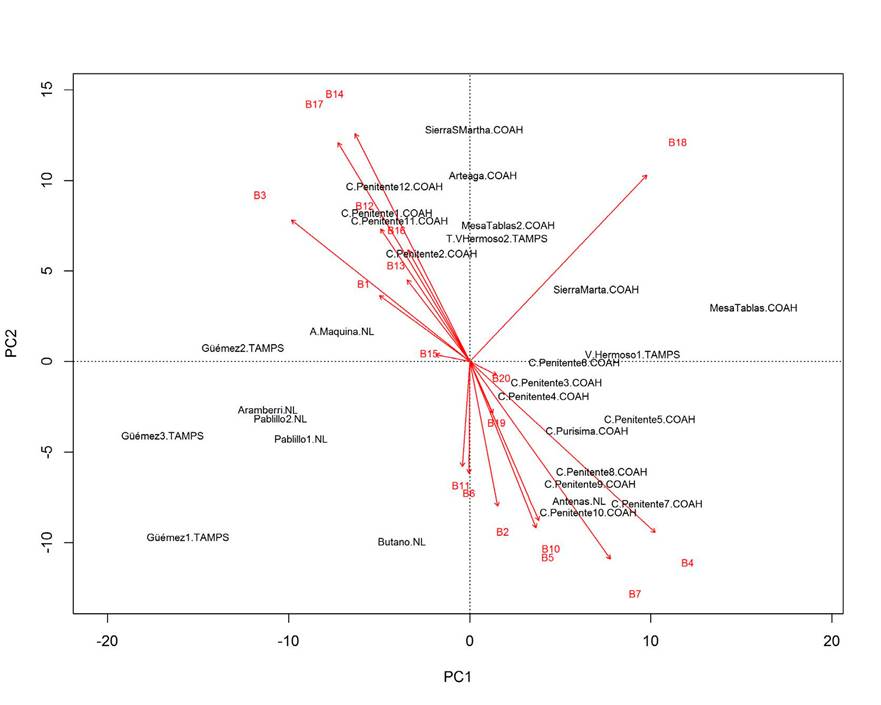
La contribución relativa de las variables ambientales y topográficas mediante el análisis de componentes principales (PCA) para A. vejarii mostró que PC1, PC2 y PC3 explican 80% de la varianza. El rango anual de temperatura (B7), la precipitación del mes más seco (B14), la precipitación del trimestre más seco (B17) y la altitud sobre el nivel del mar (B18) resultaron ser las variables más importantes para explicar la distribución potencial de A. vejarii (Fig. 4).
Impacto antropogénico
Se determinó la relación de la distancia de los sitios de muestreo a un asentamiento humano más cercano a ellos, utilizando el índice de impacto antropogénico se observó una asociación negativa y significativa entre ambas variables (Spearman rs=-0.89, p=0.007, n=9, Fig. 5), es decir a mayor distancia menor impacto.
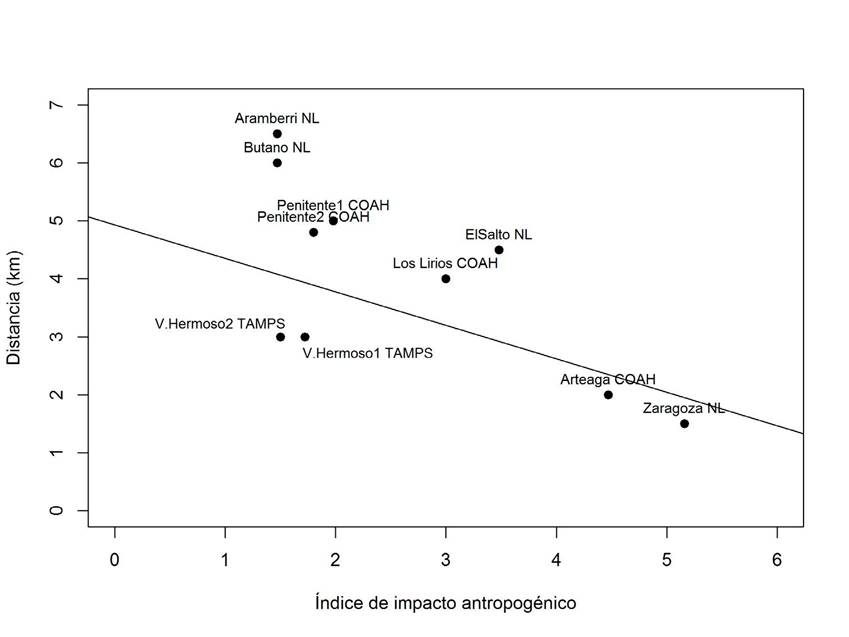
Figura 5: Índice de impacto antropogénico para cada población de Abies vejarii Martínez con respecto a la distancia de ubicación a un asentamiento humano próximo.
Los resultados del modelo lineal (Cuadro 1) mostraron que el efecto de la probabilidad de presencia obtenida del ejercicio de modelado (idoneidad climática) sobre las poblaciones naturales de la especie fue significativo y tuvo un efecto positivo general. En cuanto a los factores topográficos, la altitud mostró un efecto negativo significativo, mientras que la pendiente fue estadísticamente significativa.
Cuadro 1: Resultados del modelo lineal para estimar los efectos de la idoneidad climática y variables topográficas (altitud pendiente y exposición) en poblaciones naturales de Abies vejarii Martínez. p-valor: Probabilidad de significancia menor a 0.05
| Estimador | Error estándar | Valor de t | Significancia | p-valor | |
| Intercepto | -6.9567183 | 1.9237258 | -3.616 | 0.001 | 0.00029 |
| Idoneidad climática | -0.0344788 | 0.0046211 | -7.461 | 0.001 | 8.58e-14 |
| Exposición | 0.0026778 | 0.0011122 | 2.408 | 0.01 | 0.016051 |
| Altitud | 0.0072333 | 0.0006348 | 11.394 | 0.001 | <2e-16 |
| Pendiente | 0.1110882 | 0.0233547 | 4.757 | 0.001 | 1.97e-06 |
Discusión
El modelo de distribución potencial obtenida muestra con precisión la idoneidad climática de A. vejarii, ya que los resultados son congruentes con un estudio previo de modelo de nicho llevado a cabo exclusivamente para el género Abies en México (Martínez-Méndez et al., 2016). El modelo resultante muestra una superficie total adecuada para la especie de 67,096 ha (Fig. 2), representando 42.7% con respecto a la superficie total del bosque de oyamel reportado en México (FAO, 2015). Su área potencial adecuada es mucho más grande que el área conocida. Específicamente, la especie podría potencialmente habitar a lo largo de la SMOR donde actualmente no se ha registrado.
Sin embargo, estudios de modelos de proyección a futuro (2050, 2080, 2100) para especies de alta montaña concluyen que el cambio climático representa una amenaza innegable para su existencia, además del cambio en el uso del suelo que representa la causa más grave para su extinción (Keenan, 2016; Marchi et al., 2016). Las especies sensibles a la temperatura pueden responder a un clima cálido moviéndose a lugares más fríos en latitudes o altitudes más altas (Tang et al., 2017). Ante estos escenarios los bosques del oyamel (A. vejarii) podrían ser las más vulnerable, debido a las pocas oportunidades de migración ascendente.
Estudiar las variables ambientales, topográficas y antropogénicas en la distribución de las especies es de vital importancia, ya que permite entender las implicaciones en el desarrollo espacio-temporal de sus poblaciones a una escala local, proporcionando información sobre el efecto potencial del calentamiento global en la idoneidad climática para su desarrollo y determinar posibles áreas de refugio para el oyamel y con ello mitigar el impacto climático a futuro.
El análisis CA mostró una clasificación clara con respecto a la altitud de distribución de las poblaciones naturales de A. vejarii. Esto explica la contribución del sistema montañoso complejo del noreste de México, donde existe un fuerte gradiente altitudinal (Castillón et al., 2015). Además, otros factores como el cambio de uso de suelo también pueden influir en la distribución de las especies, mediante la transformación y pérdida de superficie en las coberturas vegetales, generando cambios en la composición y estructura de los ecosistemas.
La distribución de la especie a través de gradientes altitudinales estuvo estrechamente relacionada con variables ambientales más secas (temperatura y precipitación). Tales relaciones indican la sensibilidad de los bosques templados al estrés por sequía en el norte de México (Cabral-Alemán et al., 2017; Pompa-García et al., 2017a, b). Las condiciones más cálidas podrían conducir a una reducción en el crecimiento radial de algunas coníferas; además pueden afectar negativamente la regeneración de especies de alta montaña y la estructura y composición futura de los ecosistemas relictos (Thijs et al., 2014).
Se encontró un significativo impacto negativo por actividades antropogénicas sobre las poblaciones actuales de A. vejarii. La destrucción del hábitat ha reducido drásticamente la cubierta vegetal de algunas localidades sujetas al pastoreo (Jiménez et al., 2005). Investigaciones recientes recomiendan múltiples alternativas de manejo de los ecosistemas forestales que contienen especies relictos, como son la creación de programas de reforestación a escala local, áreas protegidas prioritarias, monitoreo del tamaño de la población y la estructura demográfica, implementación de la propagación de la especie fuera de su rango natural (como rodales artificiales en parcelas experimentales), y la combinación de información distributiva, genética y biológica se puede utilizar para su conservación (Aguilar-Soto et al., 2015; Tang et al., 2017; Wu et al., 2018). Por lo tanto, el aporte final de esta investigación es proporcionar información referente a superficies reales disponibles para la planificación y la expansión de A. vejarii con fines de conservación y restauración.
Conclusiones
Este estudio destaca que la idoneidad climática y la presión humana representan los principales factores que determinan la distribución potencial de A. vejarii en el noreste de México. La fragmentación de su hábitat y lo restringido de su distribución actual puede indicar una capacidad limitada para adaptarse a las condiciones futuras. Por lo tanto, la futura conservación de esta especie requiere de manera urgente la implementación de programas locales a corto plazo que incluya la protección de las poblaciones actuales frente a nuevas perturbaciones antropogénicas. En particular, las medidas de conservación dirigidas a la reincorporación natural de la especie mediante programas de reforestación deberían implementarse a lo largo de la Sierra Madre Oriental, con el objetivo de facilitar la regeneración natural de este árbol amenazado.

