











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
Las semillas suelen representar el éxito reproductivo de las angiospermas y permiten dispersar e incorporar nuevos genotipos a las poblaciones (Harper, 1977; Bewley, 2003; Márquez-Guzmán et al., 2013). Se sabe que una gran cantidad de semillas pueden ser depredadas por considerarse un recurso valioso para una gran diversidad de animales como roedores y hormigas (Janzen, 1971; Dirzo y Domínguez, 1986; Crawley, 2000; Daws, 2005). Por otro lado, la probabilidad que tengan las semillas de escapar de la granivoría para poder germinar y establecerse como plántulas depende de varios factores (Andersen, 1989; Daws, 2005; Holland y Molina-Freaner, 2012; Baskin y Baskin, 2014). Entre estos factores se encuentra el tiempo que las semillas puedan permanecer viables (Vázquez-Yanes y Orozco-Segovia, 1996; Bewley, 1997; Márquez-Guzmán et al., 2013), así como la disponibilidad de sitios con luz, y la temperatura y la humedad adecuadas para que las semillas germinen y las plántulas sobrevivan (Holmes y Smith, 1975; Gutterman, 2000; Ruedas et al., 2000; Baskin y Baskin, 2014; Aragón-Gastélum et al., 2018).
Lo anterior contribuye a que las semillas y las plántulas representen las etapas más vulnerables del ciclo de vida de las plantas (Harper, 1977). Por ejemplo, en poblaciones vegetales de ambientes áridos los individuos adultos pueden tolerar escenarios con temperaturas extremas y ausencia de lluvias; sin embargo, esto puede ser muy estresante para las plántulas y provocar que una gran proporción de semillas no llegue a la etapa juvenil (Baskin y Baskin, 1971; Jordan y Nobel, 1981; Godínez-Álvarez et al., 2003). Algunos reportes indican que en respuesta a largos periodos de sequía pueden ocurrir eventos reproductivos fallidos con baja o nula producción de semillas para un año particular (Epling et al., 1960; Baskin y Baskin, 1980).
La capacidad de formar un banco de semillas que contenga una muestra de los genotipos que fueron exitosos uno o varios años atrás en ambientes impredecibles les ha permitido a varias especies contrarrestar la falta de reclutamiento en un periodo desfavorable (Schafer y Chilcote, 1969; Baker, 1989; Simpson et al., 1989; Thompson, 1993; Jiménez-Sierra et al., 2022). La longevidad ecológica, que es la capacidad de las semillas de permanecer viables en condiciones naturales, le permite a las semillas permanecer viables en el suelo y de esta forma aumentar las probabilidades de reclutamiento en diferentes momentos de la dinámica de las poblaciones (Venable, 1989; Vázquez-Yanes y Orozco-Segovia, 1996; Bowers, 2000; Márquez-Guzmán et al., 2013; Baskin y Baskin, 2014). De acuerdo con el tiempo de persistencia de las semillas después de su dispersión, Thompson (1993) reconoció tres tipos de bancos de semillas: i) transitorio, con semillas que pueden permanecer viables máximo un año; ii) persistente a corto plazo, si la viabilidad de las semillas supera un año, pero no excede cinco años, y iii) persistente a largo plazo, formado por semillas muy longevas con potencial de germinar después de cinco años. Las semillas que tienen una longevidad ecológica alta podrían ser capaces de formar bancos de semillas persistentes de corto y largo plazo.
En el caso de la familia Cactaceae, ciertas características morfológicas y fisiológicas de las semillas (p. ej. un tamaño pequeño y redondeado, fotoblastismo positivo y longevidad ecológica; Rojas-Aréchiga y Batis, 2001; Barrios et al., 2020), junto con los resultados de experimentos que evalúan por cuánto tiempo las semillas pueden permanecer viables en condiciones de laboratorio y de campo, han permitido determinar la existencia de un banco de semillas para algunas especies (Fearn, 1977, 1981; de Viana, 1999; Bowers, 2000, 2005; Montiel y Montaña, 2003; Flores-Martínez et al., 2008; Pascacio-Villafán y Ortega Ortiz, 2009; Cheib y Garcia, 2012; Álvarez-Espino et al., 2014; Aranda-Pineda, 2017; Ordoñez-Salanueva et al., 2017). Además, varios estudios han comprobado la existencia de bancos de semillas en el suelo al tomar muestras del mismo y analizarlas (Bowers 2000, 2005; Ordóñez-Salanueva et al., 2017; Guzmán-Vázquez et al., 2021; Varela et al., 2021).
Algunos experimentos realizados bajo condiciones naturales han demostrado que ciertas especies pueden mantener su viabilidad por un tiempo prolongado. Por ejemplo, para Ferocactus wislizeni (Engelm.) Britton & Rose se encontraron plántulas de semillas que habían sido colocadas en el sustrato del sitio de estudio por lo menos tres años atrás; mientras que para el cactus globoso Mammillaria grahamii Engelm. se reportó un banco de semillas persistente a largo plazo, con semillas de hasta seis años que sobrevivieron y germinaron en condiciones naturales (Bowers, 2005). Un estudio que obtuvo resultados conjuntos de laboratorio y de campo reportó que las semillas de Echinocereus enneacanthus Engelm. germinaron un 62% después de estar almacenadas en laboratorio durante ocho años y posteriormente enterradas 12 meses en el sitio de estudio (Aranda-Pineda, 2017).
Por otra parte, se ha explorado bajo condiciones de laboratorio el efecto de la edad de las semillas en su capacidad germinativa; por ejemplo, las semillas de Ferocactus herrerae J.G. Ortega permanecieron viables en 50% después de 15 años de haber sido colectadas (Fearn, 1977) y las semillas de Ferocactus peninsulae (Engelm. ex Weber) Britton & Rose presentaron una germinación por arriba de 65% después de cuatro años de almacenamiento en condiciones de laboratorio (Rojas-Aréchiga y García-Morales, 2022).
En el trabajo de revisión realizado por Rojas-Aréchiga y Batis (2001), en el que se preguntan si las semillas de las cactáceas forman bancos en el suelo, se reportan más de 30 especies que cumplen con al menos una de las características morfológicas o fisiológicas que las semillas requieren para formar un banco, destacando el fotoblastismo positivo. Por lo tanto, se resalta la importancia de determinar la existencia de alguna de estas características que permiten reconocer el potencial de una especie para formar un reservorio de semillas, aún si no cuenta con experimentos en condiciones naturales (Fearn, 1977, 1981; Bowers, 2000; Rojas-Aréchiga y Batis, 2001; Lindow-López et al., 2018). El trabajo de Rojas-Aréchiga y Batis (2001) no incluye el género Lophophora J.M. Coult., pero estudios más recientes (Rojas-Aréchiga et al., 2013) demuestran que una de sus especies, Lophophora diffusa (Croizat) Bravo, de acuerdo con la categorización de Barthlott y Hunt (2000) presenta semillas medianas (Fig. 1). Las semillas medianas tienen una longitud entre 1.2 y 1.9 mm y un requerimiento de luz para germinar (Trujillo-Hernández, 2002; Rojas-Aréchiga et al., 2013). Por otro lado, Trujillo-Hernández (2002) refiere una proporción de semillas con cubierta impermeable que no deja pasar el agua hacia el embrión, lo que puede aumentar el tiempo de permanencia después de su dispersión. A pesar de esta información, aún faltan estudios que determinen el tiempo que las semillas pueden permanecer viables, tanto en condiciones naturales como en laboratorio.

Figura 1: Morfología de la semilla de Lophophora diffusa (Croizat) Bravo, en el microscopio estereoscópico. Fotografía: Mariana Rojas-Aréchiga.
Bajo el supuesto de que Lophophora diffusa presenta algunas características morfológicas y fisiológicas que le pueden permitir formar un banco de semillas, el objetivo de este estudio fue evaluar el efecto de la edad de las semillas en la germinación y la supervivencia de plántulas en condiciones de laboratorio. De esa forma se puede conocer si la respuesta de germinación sugiere la capacidad de formar un banco de semillas, lo que permitiría entender mejor la dinámica de establecimiento en su hábitat.
Materiales y Métodos
Zona de estudio
Las semillas utilizadas para este proyecto se colectaron en el municipio Peñamiller en Querétaro, México. Esta localidad forma parte de la región semiárida del estado de Querétaro, presenta un clima semicálido-semiseco con lluvias en verano, precipitación media anual de 456 mm y una temperatura media anual de 21.7 °C (CONAGUA, 2020). La vegetación de la zona de estudio es de tipo matorral xerófilo micrófilo, y cuenta con varias especies representativas como Fouquieria splendens Engelm. (ocotillo), Larrea tridentata (Sessé & Moc. ex DC.) Coville (gobernadora), Bursera fagaroides (Kunth) Engl. (xixote) y cactáceas como Echinocereus pentalophus (DC.) Lem. (alicoche falso), Mammillaria parkinsonii Ehrenb. (biznaga de aréola dorada), Strombocactus disciformis (DC.) Britton & Rose (biznaga trompo) y la especie de estudio: L. diffusa (peyote queretano).
Lophophora diffusa (Fig. 2) tiene una distribución restringida y se reconoce como endémica para la región Semidesierto Queretano-Hidalguense (Anderson, 1969; Díaz-Segura et al., 2012; Hernández-Magaña et al., 2012). Los individuos de esta especie producen pocas semillas, por lo general menos de 40 por fruto (Trujillo-Hernández, 2002; Díaz-Segura et al., 2017; Briseño-Sánchez et al., 2020) y a pesar de tener un periodo de floración extenso también producen pocos frutos por individuo al año (1.62±0.13 (x̄±e.e) Briseño-Sánchez, 2016) y presentan un bajo éxito reproductivo, pues de 42% de las plantas en una población que son reproductivas solamente 28% forma frutos (Briseño-Sánchez et al., 2020). Se ha reportado que sus poblaciones han disminuido 50% (Díaz-Segura et al., 2012). Esta especie se encuentra en la Lista Roja de la UICN bajo la categoría de Vulnerable (VU, UICN; Gómez-Hinostrosa et al., 2017) y ha sido catalogada como una especie en peligro de extinción (P) en la NOM-059-SEMARNAT-2010, en su actualización de 2019 (SEMARNAT, 2010).

Colecta de semillas, germinación de semillas y supervivencia de plántulas
Se realizaron visitas continuas a la localidad de estudio después de la época de floración de L. diffusa (julio-octubre) para colectar frutos de 20 individuos diferentes en 2014 y 2018. Debido a la escasa producción de frutos por individuo y al bajo número de semillas por fruto que presenta esta especie, las semillas no se obtuvieron de una sola cosecha y fue necesario agrupar semillas de varios meses de colecta para tener una muestra con buena cantidad de semillas para cada año.
Las semillas se extrajeron de los frutos, se almacenaron a temperatura ambiente (20 ºC±2) en bolsas de papel y se mantuvieron en esas condiciones en el Instituto de Ecología, de la Universidad Nacional Autónoma de México (UNAM), Ciudad de México, México, hasta el momento de comenzar el experimento de germinación. Este consistió en establecer como factor la edad de las semillas, con semillas colectadas el año que se realizó el experimento (2018) y semillas de cuatro años (colectadas en 2014).
Las semillas se sembraron en cajas Petri con agar bacteriológico (BD Bioxon) al 1% y se incubaron en una cámara ambiental (Lab-Line Instruments, Ciudad de México, México), a 25 ºC y fotoperiodo de 12 h (Rojas-Aréchiga et al., 2008). Se colocó un total de 100 semillas, cinco semillas por caja Petri, teniendo diez repeticiones por tratamiento y 50 semillas por grupo de edad (semillas de 2018 y semillas de 2014). El experimento se monitoreó durante un mes en 2018, y se registró la germinación de las semillas y la supervivencia de las plántulas en 15 tiempos de observación cada tercer día. Las semillas se consideraron germinadas al observar la emergencia de la radícula (Bewley y Black, 1994).
Análisis estadísticos
Los resultados de germinación se analizaron a partir de un modelo de “tiempo al evento” con un ajuste de función log-logística (Ritz et al., 2013). Ese modelo se formula en términos de la distribución F(t)=d/(1+exp(b{log(t)-log(t 50)}), donde F precisa una función de distribución acumulativa, que para cada punto de tiempo t≥0 devuelve la fracción de semillas que ya han germinado; por lo tanto, F(t) indica la fracción de semillas germinadas entre el inicio del experimento y el tiempo t. Este modelo considera tres parámetros: i) el límite superior o la proporción máxima de semillas germinadas durante el experimento (d); ii) el tiempo medio de germinación; es decir, el tiempo necesario para que se alcance 50% de la germinación total (t 50) y iii) la pendiente de la curva de germinación (se obtiene a partir del vector numérico con la respuesta medida), proporcional a la pendiente de F en el tiempo medio de germinación (b).
Las diferencias entre los parámetros estimados para el grupo de semillas de 2018 y el grupo de semillas de 2014 se compararon con pruebas de t. Para analizar el efecto de la edad de las semillas en la supervivencia de plántulas, se obtuvo la proporción de plántulas vivas por grupo de edad considerando la proporción máxima de semillas que germinaron durante el experimento. Las diferencias entre tratamientos se analizaron mediante un modelo lineal generalizado (GLM), con distribución de error binomial y función de enlace logit. Los análisis estadísticos se realizaron por medio de la paquetería drc (Ritz y Streibig, 2005) en el programa R v. 3.5.2 (R Core Team, 2020).
Resultados
Germinación de semillas y supervivencia de plántulas
La germinación de L. diffusa se registró desde el primer tiempo de observación (tres días) para las semillas de 2018, y desde el segundo tiempo de observación (a los seis días) para las semillas de colecta 2014. El grupo de semillas de 2018 mostró un porcentaje máximo de germinación de 62%, que se registró en el tiempo diez de observación (20 días), mientras que el grupo de semillas de 2014 mostró un porcentaje máximo de germinación de 52%, para el tiempo 13 de observación (26 días) (Fig. 3).
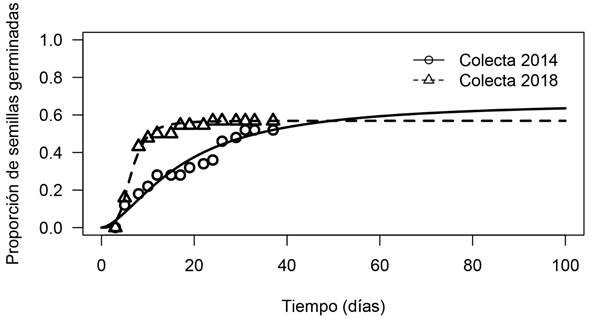
Figura 3: Efecto de la edad de las semillas en la germinación de Lophophora diffusa (Croizat) Bravo. Línea continua (colecta 2014) y línea discontinua (colecta 2018). Se presentan las curvas de germinación del modelo “tiempo al evento” con un ajuste de función log-logística.
Como se muestra en el Cuadro 1, los valores estimados para el modelo log-logístico no señalan diferencias significativas al comparar los valores del parámetro d entre tratamientos (t=0.592, p=0.553); es decir, que la proporción máxima de germinación no difiere significativamente entre el grupo de semillas de 2018 y el grupo de semillas de 2014. Sin embargo, el tiempo necesario para que se alcance 50% de la germinación total fue más del doble para las semillas de 2014 en comparación con las semillas colectadas en 2018. Por lo tanto, se encontraron diferencias significativas al comparar el parámetro b entre tratamientos (t=2.81, p=0.004); este parámetro es proporcional a la pendiente de la curva de germinación en el momento t 50.
Cuadro 1: Parámetros estimados del modelo log-logístico d/(1+exp(b{log(t) - log (t 50)}), el error estándar para cada parámetro se muestra entre paréntesis. El límite superior (parámetro d), señala la proporción de semillas de Lophophora diffusa (Croizat) Bravo que germinaron durante el experimento. El parámetro t 50 denota el momento (en días), para el que había germinado 50% de la proporción total de semillas germinadas en el experimento. El parámetro b denota la pendiente de la curva de germinación en el momento t 50. Se muestran los resultados para las semillas de 2014 y para 2018. Se marca con negritas los valores que mostraron diferencias significativas al compararlos mediante la prueba de t.
| Tratamiento | d (e.e) Límite superior | t 50 (e.e) 50% germinación (días) | b (e.e) Pendiente |
| Semillas de 2014 | 0.672(0.395) | 17.301(12.4) | -1.612(0.377) |
| Semillas de 2018 | 0.568(0.506) | 6.364(3.127) | -3.897(4.607) |
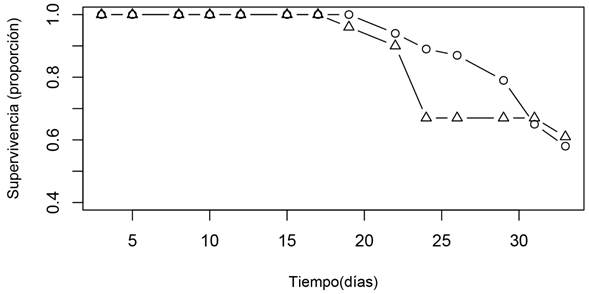
Los resultados de supervivencia de las plántulas señalan que las primeras muertes ocurrieron en el tiempo ocho de observación para las semillas de 2018 y en el tiempo nueve para las semillas de 2014. Las plántulas del grupo de semillas de 2018 tuvieron una supervivencia de 90% en los primeros nueve tiempos de observación; sin embargo, este porcentaje se redujo a 67% para el tiempo de observación diez. Por su parte, las plántulas de semillas de 2014 mostraron una supervivencia de 94 y 89% en los mismos tiempos de observación (Fig. 4). Al final del registro de supervivencia, las plántulas de semillas de menor edad (colecta de 2018) presentaron un mayor porcentaje de supervivencia: 61±5.92 (%±e.e.), en comparación con las plántulas de semillas de cuatro años (colecta 2014), con un porcentaje de supervivencia de 58±4.81 (%±e.e.), pero no hubo diferencias significativas entre edades de las semillas (n=20, χ²=0.043, g.l=1, p=0.83).
Discusión
El reconocer las características tanto morfológicas como fisiológicas de una especie en particular e inferir su potencial para formar un banco de semillas ayuda a entender su dinámica de reclutamiento y la estructura de sus poblaciones (Mandujano et al., 1998; Godínez-Álvarez et al., 2003; Eriksson y Ehrlén, 2008; Ordoñez-Salanueva et al., 2017; Aragón-Gastélum et al., 2018; Rojas-Aréchiga y García-Morales, 2022). El tiempo de inicio de germinación y el porcentaje de germinación en L. diffusa fueron muy similares independientemente de la edad de las semillas. La emergencia de la radícula se comenzó a registrar desde los primeros tiempos de observación. Al final del experimento se presentó un porcentaje de germinación por arriba de 50% para las semillas de los dos grupos de edad, lo que demuestra que las semillas de la especie de estudio no pierden viabilidad en poco tiempo a diferencia de otras especies, donde la edad de las semillas afecta su viabilidad, como es el caso del género Frailea Britton & Rose, cuyas semillas pierden viabilidad muy rápido (Fearn, 1981), o en algunas especies del género Echinopsis Zucc., para las que se reporta una disminución en la proporción de la germinación después de 12 meses de estar enterradas en el suelo (Lindow-López et al., 2018), y por ello la posibilidad de formar un banco de tipo persistente se ha descartado.
Los resultados que se han obtenido sobre estudios de viabilidad en semillas de la familia Cactaceae son muy variados (Rojas-Aréchiga y Vázquez-Yanes, 2000), porque depende de varios factores como las condiciones de colecta y almacenamiento, el contenido de humedad en las semillas y el grado de maduración de las semillas, entre otros (Barrios et al., 2020). En L. diffusa no se encontró una disminución en los porcentajes de germinación en semillas de cuatro años en comparación con semillas de menor edad. El potencial para formar un banco de semillas se ha reconocido en algunas especies de cactáceas como Ferocactus townsendianus Britton & Rose, F. peninsulae y E. enneacanthus a partir de experimentos de germinación en laboratorio (Guillén-Trujillo et al., 2014; Aranda-Pineda, 2017; Rojas-Aréchiga y García-Morales, 2022).
De acuerdo con el trabajo de Rojas-Aréchiga et al. (2013), en el cual se plantea una relación entre el fotoblastismo positivo, el tamaño de la semilla y la filogenia en varias especies de la tribu Cacteae a la cual pertenece L. diffusa, se menciona que tienen fotoblastismo positivo y semillas de tamaño pequeño a mediano. También es el caso de Aztekium ritteri (Boed.) Boed. y Epithelantha micromeris F.A.C. Weber ex Britton & Rose (Maiti et al., 1994), Ferocactus spp. Britton & Rose (Rojas-Aréchiga et al., 1997), Mammillaria spp. Haw. (Benítez-Rodríguez et al., 2004) y Turbinicarpus spp. (Backeb.) Buxb. & Backeb. (Flores et al., 2006). Las especies con semillas pequeñas o medianas, a diferencia de con semillas grandes, tienen una mayor probabilidad de escapar de los granívoros y de permanecer en el suelo, y por ende pueden tener una mayor probabilidad de formar un banco de semillas por más de un año (Thompson y Grime, 1979; Rojas-Aréchiga y Batis, 2001; Rojas-Aréchiga et al., 2013).
El fotoblastismo positivo, el cual se ha asociado a semillas de tamaño pequeño o mediano, les permite a las semillas permanecer viables en el suelo hasta que la luz dispare la germinación, junto con la humedad y una temperatura adecuada (Rojas-Aréchiga y Vázquez-Yanes, 2000; Flores et al., 2011; Rojas-Aréchiga et al., 2013). Esto es muy importante biológicamente porque las semillas pequeñas tienen pocas reservas, de tal manera que al ser fotoblásticas positivas solo pueden germinar si están poco enterradas o en la superficie del suelo donde pueden percibir la luz, germinar y emerger exitosamente (Flores et al., 2011, 2016; Rojas-Aréchiga et al., 2013; Barrios et al., 2020).
Además de la viabilidad de las semillas de hasta cuatro años de edad descrita en el presente trabajo, se ha reportado que L. diffusa cuenta con algunas características necesarias para formar un banco de semillas: semillas redondeadas de tamaño mediano (longitud promedio de 1.41±0.11 mm; Rojas-Aréchiga et al., 2013), ausencia de germinación en oscuridad, y porcentajes de germinación en luz mayores a 50% y en ocasiones mayores a 80% (Trujillo-Hernández, 2002; Rojas-Aréchiga et al., 2013). En cuanto a la velocidad de germinación, la especie de estudio mostró diferencias en la pendiente de la curva de germinación en el tiempo t 50, las semillas de mayor edad mostraron una velocidad de germinación menor, en comparación con las semillas recién colectadas, que alcanzaron el porcentaje medio de germinación desde los primeros días.
El efecto de la edad de las semillas en la velocidad de germinación ha sido poco explorado en cactáceas (Guillén-Trujillo et al., 2014). Sin embargo, para otros grupos de plantas se ha señalado que la edad de las semillas puede afectar la velocidad de germinación (Villiers, 1973). Una posible explicación de esta respuesta es que después de la hidratación las semillas de mayor edad deben pasar por un proceso de reparación de las membranas celulares antes de que puedan germinar (Villers, 1973) y posiblemente este proceso deba ser más prolongado para las semillas de mayor edad.
En el caso de L. diffusa, comprender los procesos que promueven la germinación de las semillas implica además tomar en cuenta las características que contribuyen a la formación de un banco de semillas en especies amenazadas (Sánchez-Salas et al., 2006; Barrios et al., 2020; Gurvich et al., 2021) y permite desarrollar planes de conservación tanto in situ como ex situ, al determinar el tiempo que pueden permanecer viables las semillas para su almacenamiento y para la reintroducción de plántulas (Flores et al., 2005; Gurvich et al., 2021). El almacenamiento de semillas y la reintroducción de plántulas de L. diffusa podría ser una medida favorable ya que una gran parte de sus poblaciones ha desaparecido y las poblaciones restantes se encuentran bajo amenaza (Díaz-Segura et al., 2012).
Además, habría que tomar en cuenta la baja supervivencia encontrada para muchas especies vegetales durante las primeras etapas del ciclo de vida (Harper, 1977; Buckley, 1982), y la transición de semilla a plántula ha sido considerada como un punto crítico en la dinámica de las poblaciones de cactáceas (Godínez-Álvarez et al., 2003; Pierson et al., 2013; Martínez-Ramos et al., 2016). Se ha reportado que el tamaño y la supervivencia de las plántulas puede ser una variable asociada con la longevidad de las semillas (González et al., 2009). En este trabajo encontramos que el tiempo de almacenamiento de las semillas no afectó la supervivencia de las plántulas en condiciones de laboratorio para L. diffusa, ya que cerca de la mitad de las semillas que germinaron lograron permanecer como plántulas al menos un mes y no se presentaron diferencias significativas entre los porcentajes de supervivencia de las plántulas por grupos de edad. Sin embargo, se sabe que la supervivencia de plántulas y el establecimiento de nuevos individuos en condiciones naturales podría verse afectado por varios factores, como la cantidad de semillas por fruto, los porcentajes de germinación en campo, la presencia de nodrizas y la depredación de semillas y plántulas (Mandujano et al., 2001; Godínez-Álvarez et al., 2003; Flores-Martínez et al., 2008; Ordoñez-Salanueva et al., 2017; Ortiz-Martínez et al., 2021).
Para esta población de L. diffusa se ha reportado una probabilidad de transición de semilla a plántula de 0.0039; además, las plántulas (i.e., individuos con un diámetro <1.7cm), solo representan 15% del total de la población (Briseño-Sánchez, 2019). El potencial de formar un banco de semillas para las especies que se desarrollan en ambientes impredecibles como las zonas áridas, ha sido considerado como una estrategia para que las poblaciones puedan permanecer y crecer (Harper, 1977; Baskin y Baskin, 2014; Ordoñez-Salanueva et al., 2017). En este caso, la presencia de un banco de semillas persistente a corto plazo en L. diffusa podría aumentar su propagación a partir de la fracción de semillas que permanecen viables, ya que estas semillas tienen el potencial de germinar en distintos momentos bajo condiciones favorables. Además, la presencia de semillas con la capacidad de permanecer viables en el suelo por varios meses podría disminuir los efectos de las perturbaciones y permitir la regeneración de especies amenazadas a través de la incorporación de nuevos individuos en diferentes tiempos (Houle y Phillips, 1988; Keddy et al., 1989; Baskin y Baskin, 2014).
Conclusiones
La edad de las semillas no afectó la germinación y supervivencia de las plántulas en L. difusa. La viabilidad de las semillas en condiciones de laboratorio reportadas en este trabajo, sumada a las características morfológicas y fisiológicas que ya se conocían de la especie, sugiere que L. diffusa tiene características que le permiten formar un banco de semillas persistente a corto plazo, ya que las semillas permanecen viables en condiciones de laboratorio hasta cuatro años. Sin embargo, es necesario conocer si las semillas pueden permanecer viables bajo condiciones naturales por largo tiempo. La presencia de un banco de semillas es importante en especies que se encuentran bajo amenaza, ya que disminuye los efectos de las perturbaciones, y permite su regeneración a través de la incorporación de nuevos individuos en diferentes momentos.

