











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introduction
Dahlia Cav. (Asteraceae, tribe Coreopsideae) is a genus endemic to the Americas that has been appreciated for its ornamental, nutritional, and medicinal properties since the pre-Colombian era (Sørensen, 1970; Carrasco-Ortiz et al., 2019; Reyes-Agüero, 2020). Currently, there are almost 61,000 registered Dahlia ornamental cultivars; this count includes names from the early 1800s to the present day, for which 14 different horticultural groups are distinguished, mainly based on parental wild species, plant sizes, and head structure (TRHS, 2021). Although the horticultural and botanical study of Dahlia has a history going back 230 years, knowledge about its natural diversity has experienced a noticeable boost in the last five decades, during which 60% of the currently recognized species have been described (Sørensen, 1969, 1980, 1987; Hansen and Hjerting, 1996; Saar, 1999, 2002; Saar and Sørensen, 2000, 2006; Saar et al., 2002, 2003; Hansen and Sørensen, 2003; Castro-Castro et al., 2012, 2015; Reyes-Santiago et al., 2018, 2019; Villaseñor and Redonda-Martínez, 2018).
Two centers of species richness for Dahlia in Mexico were identified by Carrasco-Ortiz et al. (2019), where new species are likely to be found. The first of these areas comprises 1089 km2 between northeast Querétaro and eastern Guanajuato, on the transition between the Sierra Madre Oriental and the Chihuahuan Desert biogeographic provinces. Here, D. coccinea Cav., D. cuspidata Saar, P.D. Sørensen & Hjert., D. dissecta S. Watson, D. linearis Sherff, D. merckii Lehm., D. mollis P.D. Sørensen, D. moorei Sherff, D. neglecta Saar, and D. scapigeroides Sherff have been recorded. The second area comprises 2178 km2 between Oaxaca and Puebla, on the limits of the Sierra Madre del Sur and the Balsas Basin, where D. apiculata (Sherff) P.D. Sørensen, D. australis (Sherff) P.D. Sørensen, D. campanulata Saar, P.D. Sørensen & Hjert., D. coccinea, D. imperialis Roezl ex Ortgies, D. macdougalli Sherff, D. merckii, D. pteropoda Sherff, D. sorensenii H.V. Hansen & Hjert., and D. tenuicaulis P.D. Sørensen inhabit.
In areas of high species richness for Dahlia it is still possible to find new species; for example, two taxa have recently been described in Oaxaca: Dahlia calzadanaVillaseñor & Redonda-Mart. (Villaseñor and Redonda-Martínez, 2018) and D. mixtecana J. Reyes, Islas & Art. Castro (Reyes-Santiago et al., 2019). Based on our recent fieldwork, revision of herbarium material, and study of Dahlia living collections from the Universidad Nacional Autónoma de México Botanical Garden (JB-UNAM, 2023), we found additional specimens from the Mixteca region of Oaxaca dissimilar to any described species in the genus.
Therefore, the objective of the present contribution consists in the description of this new species, accompanied by detailed morphological and ecological comparisons with related taxa, a distribution map, an illustration, and a key to the identification of Dahlia in the Cañada and Mixteca biocultural regions. In addition, based on comparative analysis and review of recent collections, we present an expanded morphological, ecological, and geographical distribution of D. calzadana. Finally, we use a species accumulation curve analysis to gauge the state of our knowledge of the genus Dahlia.
Materials and Methods
Taxonomic treatment
This study comprises analyses of Dahlia in the field, botanical garden, and herbaria. The morphological description and illustration of the species used traditional taxonomic methods (Simpson, 2006). Botanical terminology follows Sørensen (1969), Saar (1999), Castro-Castro et al. (2012; 2015), and Reyes-Santiago et al. (2018; 2019). The identification key and distribution map were based on herbarium specimens in the following herbaria: ANSM of the Universidad Autónoma Agraria Antonio Narro, CHAPA of the Colegio de Postgraduados, CIIDIR of the Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional Unidad Durango, CIMI of the Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional Michoacán, CREG of the Instituto Tecnológico Agropecuario de Jalisco, ENCB of the Escuela Nacional de Ciencias Biológicas-Instituto Politécnico Nacional, FEZA of the Facultad de Estudios Superiores Zaragoza- Universidad Nacional Autónoma de México, GUADA of the Universidad Autónoma de Guadalajara, IBUG of the Instituto de Botánica-Universidad de Guadalajara, IEB of the Centro Regional del Bajío-Instituto de Ecología, A.C.-Michoacán, MEXU of the Universidad Nacional Autónoma de México, QMEX of the Universidad Autónoma de Querétaro, SLPM of the Universidad Autónoma de San Luis Potosí, XAL of the Instituto de Ecología A.C.-Veracruz, and ZEA of the Departamento de Ecología y Recursos Naturales-Universidad de Guadalajara (acronyms according to Thiers, 2024), and adjustments to the results of Carrasco-Ortiz et al. (2019), Reyes-Santiago et al. (2018; 2019).
Conservation status
The extent of occurrence (EOO) and area of occupancy (AOO) were estimated using the GeoCAT tool (Bachman et al., 2011), and a preliminary conservation status was assessed following the IUCN (2022) guidelines of categories and criteria.
Species prediction richness
We followed the procedures and suggestions described by Soberón and Llorente (1993) and Jiménez-Valverde and Hortal (2003) to infer a species richness prediction. The analysis included 41 species (Reyes-Santiago et al., 2018, 2019; Villaseñor and Redonda-Martínez, 2018; Carrasco-Ortiz et al., 2019), 2297 herbarium records obtained by procedures suggested by Carrasco-Ortiz et al. (2019), and each year between 1791 and 2021 as a measure of the sampling effort. Parametric (Clench) and non-parametric (ACE, ICE, Bootstrap, Chao and Jackknife) coefficients were calculated using STATISTICA v. 7 (StatSoft Inc., 2004) and EstimateS v. 9.1 (Colwell, 2019).
Results
Taxonomic treatment
Dahlia gypsicola J. Reyes, Ortiz-Brunel & Art. Castro, sp. nov. Figs. 1, 2.
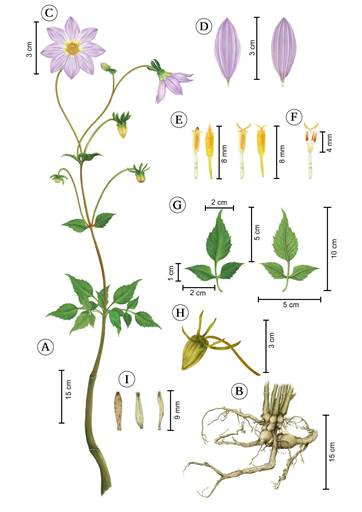
Figure 1: Dahlia gypsicola J. Reyes, Ortiz-Brunel & Art. Castro. A. complete branch; B. rhizome and tuberous roots; C. frontal and lateral view of a head and involucre; D. dorsal and ventral view of ray florets; E. disc floret variations; F. androecium and gynoecium details; G. adaxial and abaxial views of leaf; H. fruiting head; I. dorsal and ventral views of cypselae. Illustrated by Ariadna Arenas and Brenda A. Guerrero Rivera.

Figure 2: Dahlia gypsicola J. Reyes, Ortiz-Brunel & Art. Castro. A. habit and habitat; B. rootstock and thickened roots; C-D. simple and trifoliate leaves; E-G. capitulum from various angles. Photographs by Arturo Castro-Castro.
TYPE: MEXICO. Oaxaca, municipality San Sebastián Tecomaxtlahuaca, Laguna Encantada, 2.2 km al N de Santiago Juxtlahuaca, 1660 m, 17°21'54''N, 98°01'11.1''W, 12.X.2020, A. Castro-Castro et al. 4690 (holotype: CIIDIR!, isotypes: IBUG!, IEB!, MEXU!, OAX!, QMEX!, SERO!, SLPM!, US!, XAL!).
Dahlia gypsicola is similar to D. calzadana but differs by its herbaceous habit, 0.5-0.95 m tall (vs. subshrub, 0.4-1.5 m tall); semi-succulent leaves and stems (vs. succulent); simple leaves 9.5-15 × 2-5.5 cm, obtuse, serrate and glabrous margins, petiole 2.5-7 cm long (vs. (2.2-)9-15 × 3-9.5 cm, often slightly hastate, crenate-serrate and ciliate margins, petioles (4-)6.1-9.5 cm long); trisected leaves 10-22 × 3.5-5.5 cm, petiole 4-6 cm long, basal segments 2-3 × 1-2 cm, terminal segment 4.8-5.5 × 2-2.5 cm, petiolule 1-2 cm long (vs. (11-)16-23 × 11-17 cm, petiole 7-11 cm long, basal segments 5-9.5 × 3-5.3 cm, terminal segment 6.5-14.8 × 4-7.4 cm, petiolule 1-4 cm long); synflorescence (1-)3-9 capitula (vs. 1-3); capitulum 2-4 cm wide across the extended rays (vs. 4.5-7.5 cm); disk florets 36-46, funnel-shaped (vs. 40-67, tubular).
Perennial herbs, 0.5-0.95 m tall, from rootstock and thickened roots 5-15 × 2-7 cm; stems semi-succulent, erect or decumbent, slightly cespitose, solid, woody, 3-5 mm diameter, with conspicuous lenticels, cylindrical, purple to greyish green towards nodes, glabrous, internodes 3-9 cm long, solid, glabrous, greyish green; leaves simple, trisected, or both in a single individual, semi-succulent, lanceolate in general outline, adaxial surface glabrate, simple hairs concentrated towards the base of primary and secondary veins and in petiole margins, abaxial surface glabrous, stipels absent; simple leaves 9.5-15 × 2-5.5 cm, cuspidate, obtuse, oblique, margins serrate, glabrous, 3-7 teeth per side, mucronulate, sinus frequently with a short tuft, petioles 2.5-7 cm long, solid, canaliculate; trisected leaves 10-22 × 3.5-5.5 cm, petiole 4-6 cm long, solid, canaliculate, basal segments 2-3 × 1-2 cm, opposite, sessile, ovate-lanceolate, cuneate, cuspidate, terminal segment 4.8-5.5 × 2-2.5 cm, ovate-lanceolate, cuneate, cuspidate, margins serrate, 2-4 teeth per side, mucronulate, sinus frequently with a short tuft, petiolule 1-2 cm long; synflorescence corymbiform; capitula (1-)3-9; peduncles 4-10 cm long, solid, glabrescent, green to purplish, peduncle leaves 1-2 per node, simple, entire, 0.8-3 × 0.2-1 cm, linear, lanceolate to deltoid, glabrous; capitulum 2-4 cm wide across extended rays; involucre 1-1.5 × 0.6-1 cm, campanulate; phyllaries biseriate; outer series 5(-7), reflexed to adpressed in anthesis, narrowly spatulate 1.3-1.5 × 0.2-0.3 cm, glabrous, apex acute or rounded, adaxial surface green with 3-5 purple resiniferous ducts, abaxial surface green, concolorous, tuberculate; inner series 8, yellow-greenish when fresh, brown-translucent when dry, glabrous, lanceolate, 12-13 × 4.5-7 mm, acute to rounded, with 17-20 light brown to purple resiniferous ducts; receptacle convex to plane, 4-6.5 mm diameter, glabrous; paleae 1.4-1.8 × 6-8 mm, pale yellow, translucent, bright when dehydrated, resembling inner phyllaries; ray florets 8, uniseriate, neuter, limb violet with whitish bases, ovate, 2.5-3 × 1-1.2 cm, 3-5 marked ribs outside, glabrous, apices obtuse, entire, tube 3-4 × 0.6-0.9 mm, glabrescent; disk florets 36-46, corollas yellow, funnel-shaped, 5.5-8 mm long; corolla tube 1.5-2 × 0.4-0.5 mm, glabrescent but with trichomes concentrated towards the base of enlarged portion of corolla; throat 1.5-1.8 mm; lobes 5, 0.5-0.7 mm long, reflexed to deflexed, triangular; anthers 3.8-4.5 mm long, yellow, apical appendages triangular-acuminate, base short-sagittate, filaments 0.9-1 mm long, applanate; pollen yellow; style 6.5-7 mm long; style branches 1.5-1.7 mm long, applanate, acute, ciliate-papillose, yellow; cypsela clavate, 8.5-9.5 × 1.5-2 mm, furrowed in dorsal view, with a marked rib along its ventral surface, brown, glabrate, short setulae on the angles, attenuate; carpopodium annular; pappus irregular, short cup-shaped.
Distribution and ecology: the only known population of Dahlia gypsicola is located near Laguna Encantada in Oaxaca. This locality is part of a center of endemism for the gypsicolous flora of Mexico (Ortiz-Brunel et al., 2023), and hosts such gypsophiles as Echeveria subcorymbosa Kimnach & Moran (Crassulaceae), Hechtia gypsophila López-Ferr., Espejo & Hern.-Cárdenas (Bromeliaceae), Pinguicula medusina Zamudio & Studnička and P. pygmaea Rivadavia, E.L. Read & A. Fleischm. (Lentibulariaceae). Dahlia gypsicola grows on shady and humid slopes of gypsum cliffs with an eastern exposure, where they are associated with xerophilous scrub and tropical deciduous forest between 1660 and 1750 m altitude (Fig. 3). Key morphological traits that distinguish D. gypsicola include its semi-succulent leaves and stems and perenniality (Fig. 2). Perennial tissues and storage organs are common plant adaptations associated with growth on edaphically stressful soil substrates such as gypsum (Moore et al., 2014). Common associated genera and species include Agave L., Dasylirion Zucc., Echeandia taxacana Cruden (Asparagaceae), Begonia L. (Begoniaceae), Bursera Jacq. ex L. (Burseraceae), Castilleja Mutis ex L.f. (Plantaginaceae), Dioscorea L. (Dioscoreaceae), Muhlenbergia Schreb. (Poaceae), Oxalis L. (Oxalidaceae), Pinguicula L. (Lentibulariaceae), Selaginella P. Beauv. (Selaginellaceae), and Sisyrinchium L. (Iridaceae).
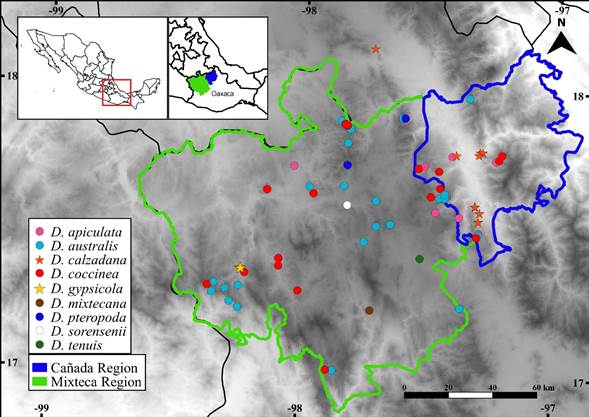
Figure 3: Distribution of Dahlia sect. Dahlia in the Cañada and Mixteca regions of Oaxaca, Mexico. Biocultural regionalization based on Carrillo González and Ramírez Alcántara (2013) and Ordoñez (2000).
Phenology: the new species flowers from late September to November and bears fruits from November to December. The plants can be found in a vegetative state throughout the year, although only the slightly woody bases of the branches are present, which are shorter than plants in flower or fruit and lack leaves.
Etymology: the name refers to the gypsum soil substrate upon which the species grows, an uncommon habitat for Dahlia.
Conservation assessment: Dahlia gypsicola is known from four collections near each other (Fig. 3). The EOO is 0.67 km2, and the size of the AOO is 4 km2, based on cells of 2 × 2 km. According to IUCN (2022) criteria, if the EOO is less than AOO, it should be changed to make them equal and ensure consistency with the definition of AOO, which is an area defined within the EOO. Thus, following the criteria (extent of occurrence estimated to be less than 100 km2 and known to exist at only a single location), a preliminary category of Critically Endangered is proposed (CR B1a).
Additional specimens examined: MEXICO. Oaxaca, municipality San Sebastián Tecomaxtlahuaca, Laguna Encantada, 2.2 km al N de Santiago Juxtlahuaca, 1660 m, 17°21'54''N, 98°01'11.1''W, 12.X.2020, A. Castro-Castro et al. 4690a (CIIDIR, IBUG); Laguna Encantada, laderas de piedra caliza y yeso detrás de la laguna, 1640 m, 17°21'56''N, 98°01'13''W, 24.IX.2019, J. P. Ortiz-Brunel et al. 663 (IBUG); Laguna Encantada, 4 km al norte de Santiago Juxtlahuaca, 1703 m, 17°22'01''N, 98°01'26''W, 25.VI.2009, J. Reyes-Santiago and C. Brachet 6890 (MEXU); Laguna Encantada, 4 km al norte de Santiago Juxtlahuaca, 1733 m, 17°22'6.5''N, 98°01'20.4''W, 1.XI.2011, J. Reyes-Santiago et al. 7352 (MEXU).
Taxonomic notes: Dahlia gypsicola has semi-succulent stems, with solid internodes and petioles, opposite leaf segments, and reflexed outer phyllaries. These characters allow us to place this species in Dahlia sect. Dahlia. Based on the identification key to species of Oaxaca state (Reyes-Santiago et al., 2019), D. gypsicola appears to be related to D. calzadana; however, D. gypsicola differs in habit, succulence of stems and leaves, and leaf structure. There are also differences in capitula number, length of the peduncles, capitulum diameter, involucre dimension and shape, external and internal phyllaries size, ray florets size, disk florets quantity, size, and shape, and cypsela size, shape, and ornamentation. The differences described above are summarized in Table 1 and in the identification key provided below.
Table 1: A comparison of morphology, ecology, phenology, and geographical distribution between Dahlia calzadana Villaseñor & Redonda-Mart. and D. gypsicola J. Reyes, Ortiz-Brunel & Art. Castro.
| Character | D. calzadana Villaseñor & Redonda-Mart. | D. gypsicola J. Reyes, Ortiz-Brunel & Art. Castro |
|---|---|---|
| Habit | subshrub | herb |
| Plant height (m) | 0.4-1.5 | 0.5-0.95 |
| Stem texture; colour | succulent; greyish green | semi-succulent; purple to greyish green |
| Leaf texture | succulent | semi-succulent |
| Simple leaves length (cm) | (2.2-)9-15 × 3-9.5 | 9.5-15 × 2-5.5 |
| Simple leaves margins | crenate-serrate and ciliate | serrate and glabrous |
| Simple leaves petioles length (cm) | (4-)6.1-9.5 | 2.5-7 |
| Trisect leaves length (cm) | (11-)16-23 × 11-17 | 10-22 × 3.5-5.5 |
| Trisect leaves petioles length (cm) | 7-11 | 4-6 |
| Trisect leaves basal segments length (cm) | 5-9.5 × 3-5.3 | 2-3 × 1-2 |
| Trisect leaves terminal segment length (cm) | 6.5-14.8 × 4-7.4 | 4.8-5.5 × 2-2.5 |
| Trisect leaves terminal segment petiole length (cm) | 1-4 | 1-2 |
| Synflorescence (number of capitula) | 1-3 | (1-)3-9 |
| Peduncle length (cm) | (3-)10-22 | 4-10 |
| Capitulum diameter across extended ray limbs (cm) | 4.5-7.5 | 2-4 |
| Involucre size (cm) | 1-2.4 × 1.5-2.5(-3.2) | 1-1.5 × 0.6-1 |
| Involucre shape | campanulate to hemispheric | campanulate |
| Outer phyllaries (number) | 5-6 | 5(-7) |
| Outer phyllaries size (cm) | 0.6-1.6 × 0.1-0.4 | 1.3-1.5 × 0.2-0.3 |
| Inner phyllaries size (mm) | 9-15 × 3.2-5.7 | 12-13 × 4.5-7 |
| Ray limb size (cm) | 2.3-3.5 × 1.2-2 | 2.5-3 × 1-1.2 |
| Ray limb colour | whitish to soft pink | violet with whitish bases |
| Disk florets (number) | 40-67 | 36-46 |
| Disc corolla shape | tubular | funnel-shaped |
| Disc corolla lenth (mm) | 6.5-10.2 | 5.5-8 |
| Disc corolla tube length (mm) | 2.2-2.6 | 1.5-2 |
| Disc corolla lobes | erect to deflexed | reflexed to deflexed |
| Anther cylinder length (mm) | 5.5-6 | 3.8-4.5 |
| Cypsela length (mm) | 9.8-10.5 | 8.5-9.5 |
| Cypsela shape; indumentum | ellipsoid; puberulent to sparsely pilose | clavate; glabrate |
| Pappus | absent | irregular short cup-shaped |
| Habitat | transition between tropical deciduous, tropical subdeciduous and Quercus forest; on limestone outcrops | transition between rosetophile scrub and tropical deciduous forest; on gypsum outcrops |
| Biogeographic province | Balsas Basin | Sierra Madre del Sur |
| Phenology (flowering and fruiting) | May to November | September to December |
| Distribution | Cañada region in Oaxaca and Puebla states | Mixteca region in Oaxaca state |
| Altitude range (m) | 1100-1380 | 1660-1750 |
Both species are found in Oaxaca, but Dahlia calzadana also occurs in the southern portion of Puebla state. Dahlia calzadana and D. gypsicola are allopatric and do not overlap in their ecological preferences or phenology. Dahlia gypsicola grows in the Sierra Madre del Sur biogeographical province, and possibly in the upper Balsas Basin, on gypsum outcrops between 1660-1750 m altitude in the transition between xerophilous scrub and tropical deciduous forest. It flowers from September to November. Meanwhile, D. calzadana inhabits the Balsas Basin biogeographical province, in the upper basin of the Papaloapan river or Pacific Basin, on limestone outcrops at 1100-1380 m of altitude in the ecotones of tropical deciduous forest, subdeciduous tropical forest, and Quercus forest. Dahlia calzadana flowers from May to November. In addition, based on a biocultural regionalization (Carrillo González and Ramírez Alcántara, 2013; Ordoñez, 2000), D. gypsicola grows in the western Mixteca region and D. calzadana is found in the Cañada region (Fig. 3).
Analysis of herbarium and living specimens allowed us to identify morphological, geographical, and ecological variations not documented in the original description of Dahlia calzadana (Villaseñor and Redonda-Martínez, 2018). Thus, an expanded description, new distribution map, clarification of phenology, and herbarium material not previously reported for the species are stated below.
Dahlia calzadanaVillaseñor & Redonda-Mart. Phytotaxa 362(2): 240-242, f. 1-3. 2018. Fig. 4.

Figure 4: Dahlia calzadana Villaseñor & Redonda-Mart. A. habit and habitat; B. succulent stem; C-D. individuals with simple and trifoliate leaves; E. front view of a head. Photographs by María de los Angeles Isla-Luna (A) and Jerónimo Reyes-Santiago (B-E).
TYPE: MEXICO. Oaxaca, municipality San Juan Bautista Cuicatlán, 3 km antes del poblado San Juan Coyula, sobre la carretera vecinal de terracería de Quiotepec a San Juan Coyula, 1250 m, 17°55'24.74''N, 96°56'15.6''W, 20.VII.2003, J. I. Calzada 23910 (holotype: MEXU!, isotypes: IEB!, TEX, XAL).
Subshrubs, 0.4-1.5 m tall, from rootstock and thickened roots 5-30 × 2-7 cm; stems succulent, erect, solid, woody, 10-20 mm diameter, striate, cylindrical, hollow, greyish green, glabrous, internodes 5-17 cm long, solid, glabrous, greyish green; leaves simple, trisected, or both in a single individual, succulent, lanceolate to ovate in general outline, adaxial and abaxial surface glabrous to glabrate, short simple hairs towards the bases of the primary and secondary veins, young leaves scarcely pilose below, stipels absent; simple leaves (2.2-)9-15 × 3-9.5 cm, cuspidate, obtuse, oblique, margins often slightly hastate, crenate-serrate and ciliate, 2-8 teeth per side, mucronulate to rounded, sinus frequently with a short tuft, petioles (4-)6.1-9.5 cm long, solid, canaliculate; trisected leaves (11-)16-23 × 11-17 cm, petiole 7-11 cm long, solid, canaliculate, basal segments 5-9.5 × 3-5.3 cm, opposite, sessile, ovate to lanceolate, cuneate, cuspidate, terminal segment 6.5-14.8 × 4-7.4 cm, ovate-lanceolate, cuneate, cuspidate, margins serrate, 2-4 teeth per side, mucronulate, sinus frequently with a short tuft, petiolule 1-4 cm long; synflorescence corymbiform, capitula 1-3; peduncles (3-)10-22 cm long, solid, glabrescent, green to yellow, peduncle leaves 1-2 per node, simple, entire to serrate, 1-2 × 0.5-1 cm, linear, lanceolate to deltoid, glabrous; capitulum 4.5-7.5 cm wide across the extended rays; involucre 1-2.4 × 1.5-2.5(-3.2) cm, campanulate to hemispheric; phyllaries biseriate; outer series 5-6, reflexed in anthesis, narrowly spatulate, 0.6-1.6 × 0.1-0.4 cm, glabrous, acute or rounded apex, adaxial surface green with 3-5 purple resiniferous ducts, abaxial surface green concolorous, smooth; inner series 8, yellow-greenish in fresh, brown-translucent when dry, glabrous, lanceolate, 9-15 × 3.2-5.7 mm, acute to rounded, with 15-18 light brown resiniferous ducts; receptacle convex to plane, 5.5-7.5 mm diameter, glabrous; paleae 1.5-2 × 6-8 mm, pale yellow, translucent, bright when dehydrated, resembling inner phyllaries; ray florets 8(-10), uniseriate, neuter, whitish to soft pink limb, ovate, 2.3-3.5 × 1.2-2 cm, 3-5 marked ribs outside, glabrous, apices obtuse, entire, tube 2.8-4.4 × 0.6-1 mm, glabrescent, glandular trichomes; disk florets 40-67, corollas yellow, tubular, 6.5-10.2 mm long; corolla tube 2.2-2.6 × 0.4-0.6 mm, pilose; throat 1.6-2 mm; lobes 5, 0.4-0.8 mm long, erect to deflexed, triangular; anthers 5.1-5.3 mm long, yellow, apical appendages triangular-acuminate, base short-sagittate, filaments 1-1.2 mm long, applanate; pollen yellow; style 5.5-6 mm long; style branches 3-3.5 mm long, applanate, acute, ciliate-papillose, yellow; cypsela ellipsoid, 9.8-10.5 × 1.5-2.5 mm, furrowed in dorsal view, brown, puberulent to sparsely pilose, scabrous on rounded angles, attenuate; carpopodium annular; pappus absent.
Distribution and ecology: Dahlia calzadana is found in Oaxaca and Puebla (Fig. 3) in the ecotones of tropical deciduous forest, subdeciduous tropical forest, and Quercus forest, on alkaline soils with limestone outcrops at 1100-1380 m altitude. The associated genera and species include Actinocheita F.A. Barkley, Pseudosmodingium Engl. (Anacardiaceae), Bakeridesia subcordata (Hochr.) D.M. Bates (Malvaceae), Beaucarnea gracilis Lem., B. stricta Lem., Dasylirion (Asparagaceae), Cephalocereus tetetzo (F.A.C. Weber ex J.M. Coult.) Diguet, Echinocactus platyacanthus Link & Otto, Mammillaria polyedra Mart. (Cactaceae), Cnidoscolus Pohl, Euphorbia tanquahuete Sessé & Moc. (Euphorbiaceae), Dioon spinulosum Dyer ex Eichler (Zamiaceae), Nahuatlea V.A. Funk (Asteraceae), and Thouinidium Radlk. (Sapindaceae).
Phenology: Dahlia calzadana flowers and fruits from May to November. The plants can be found in a vegetative state throughout the year.
Conservation assessment: We documented 11 records of Dahlia calzadana, from one locality in southern Puebla and four in Oaxaca (Fig. 3). The EOO was 969.8 km2, and the AOO was estimated at 32 km2. We propose a category of endangered (EN B1a) based on the EOO and AOO size results and following the IUCN (2022) criteria (EOO to be less than 5000 km2 and AOO less than 500 km2).
Specimens examined: MEXICO. Oaxaca, municipality San Juan Bautista Cuicatlán, Barranca de las Guacamayas, 1187 m, 17°40'36''N, 96°57'55''W, 29.IX.2002, J. Calónico-Soto et al. 23908 (MEXU); loc. cit., J. Calónico-Soto et al. 23968 (MEXU); 3 km antes del poblado San Juan Coyula, sobre la carretera vecinal de terracería de Quiotepec a San Juan Coyula, 1250 m, 17°55'24.74''N, 96°56'15.6''W, 20.VII.2003, J. I. Calzada 23910 (MEXU); Cueva de la barranca de las Guacamayas, San José El Chilar, 1111 m, 17°40'39''N, 96°57'58''W, 27.X.2001, C.A. Cruz-Espinoza and R. García 283 (MEXU); Barranca de las Guacamayas, San José El Chilar, 1178 m, 17°40'40''N, 96°57'57''W, 29.IX.2001, R. García 135 (MEXU); San Juan Coyula, 20.VI.2006, R. L. García-García et al. 880 (MEXU); Torre eléctrica 292, Dominguillo, 1100 m, 17°38'54''N, 96°56'38''W, 4.X.2002, J. Pérez-Crisanto et al. 448 (MEXU); Loma de Buena Vista, 10 km al E de Quiotepec, 1300 m, 17°55'26''N, 96°56'57.5''W, 5.V.1990, A. Salinas et al. 5421 (MEXU). Municipality Santa María Ixcatlán, Río Seco-Río Santiago, al SSO de Tecomavaca, 1150 m, 17°53'N, 97°05'W, 19.VII.1993, A. Salinas et al. 7214 (MEXU). Municipality Santiago Nacaltepec, a 2 km en línea recta al NE de San Juan Tonaltepec, 1380 m, 17°36'29.1''N, 96°56'56.2''W, 30.X.2004, G. Juárez-García and C. A. Cruz-Espinoza 655 (MEXU). Puebla, municipality Zapotitlán, 2 km al SW de San Antonio Texcala, por la carretera a Huajuapan de León, 1615 m, 18°22'46.48''N, 97°26'15.07''W, 4.VII.1981, F. Chiang et al. 2106a (MEXU).
Key to species of Dahlia occurring in Cañada and Mixteca biocultural regions, Oaxaca-Puebla, Mexico (adapted from Reyes-Santiago et al. 2019)
1a. Ray limbs yellow, orange, orange-red, scarlet, bright red, dark red, or blackish red in living plants ………………………………………………………………………………… 2
1b. Ray limbs whitish-purple, purple or violet-purple, seldom reddish purple or pure whitish in living plants ………...…………………………………………………………… 4
2a. Plants 0.45-3 m tall; leaves 12-35 cm long, simple to trisected and alternate or whorled; ray limbs yellow, orange, orange-red, scarlet, bright red, dark red or blackish red …………………………………………………………………………..… D. coccinea Cav.
2b. Plants 0.25-1.2 m tall; leaves 6-18 cm long, pinnatisect to bipinnatisect and alternate; ray limbs yellow …………………………………………………………..………………….… 3
3a. Plants 0.25-0.6 m tall; stems conspicuously pubescent; leaves pinnatisect; petioles 0.8-4.5 cm long; 1-4 capitula per plant or on each main branch; disc florets 3.5-4.5 mm long …………………………………………………………...… D. tenuis B.L. Rob. & Greenm.
3b. Plants 0.9-1.2 m tall; stems smooth to puberulent; leaves bipinnatisect; petioles (3-)6.5-7 cm long; 3-6 capitula per plant or on each main branch; disc florets 7-10 mm long ………………………………………………….... D. mixtecana Reyes, Islas & Art. Castro
4a. Petioles broadly auriculate-winged at the base, 0.8-1.2 cm wide ………………..… 5
4b. Petioles wingless …………………………………………………………………… 6
5a. Stems plicate; median leaves not markedly scabrous above ….… D. pteropoda Sherff
5b. Stems without distinct furrows; median leaves usually scabrous above ……………………………………………………...… D. sorensenii H.V. Hansen & Hjert.
6a. Stems herbaceous; leaves pinnatisect or bipinnatisect ……………………………………………………….… D. australis (Sherff) P.D. Sørensen
6b. Stems woody; leaves entire or pinnatisect ………………………………………..… 7
7a. Leaves pinnatisect; ray limbs 3.5-5.5 cm long ………..… D. apiculata P.D. Sørensen
7b. Leaves entire or pinnatisect; ray limbs 2.5-3.5 cm long ………………………….… 8
8a. Subshrubs; stems succulent, greyish green; leaves succulent; simple leaves often slightly hastate at base, margins crenate-serrate and ciliate; trisect leaves petiole 7-11 cm long, basal segments 6-9.5 × 3-5.3 cm, terminal segment 9-14.8 × 4-7.4 cm, petiolule 1-4 cm long; synflorescence of 1-3 capitula; capitulum 4.5-7.5 cm diameter across extended ray limbs; disk florets 40-67, corollas tubular ………….... D. calzadana Villaseñor & Redonda-Mart.
8b. Herbs; stems not succulent, purple to greyish green; leaves semi-succulent; simple leaves obtuse at base, margins serrate and glabrous; trisect leaves petiole 4-6 cm long, basal segments 2-3 × 1-2 cm, terminal segment 4.8-5.5 × 2-2.5 cm, petiolule 1-2 cm long; synflorescence of (1-)3-9 capitula; capitulum 2-4 cm diameter across extended ray limbs; disk florets 36-46, corollas funnel-shaped .… D. gypsicola J. Reyes, Ortiz-Brunel & Art. Castro
Species accumulation curve
A quantitative prediction of species richness suggests that knowledge of diversity in Dahlia is close to reaching a plateau. The completeness estimators indicate similar scenarios. Inventory is approaching 95% with a prediction of total species diversity of approximately 43 (Fig. 5). Nearly 62% of the specimens deposited in collections were collected in the last four decades, a period in which 16 species were described.
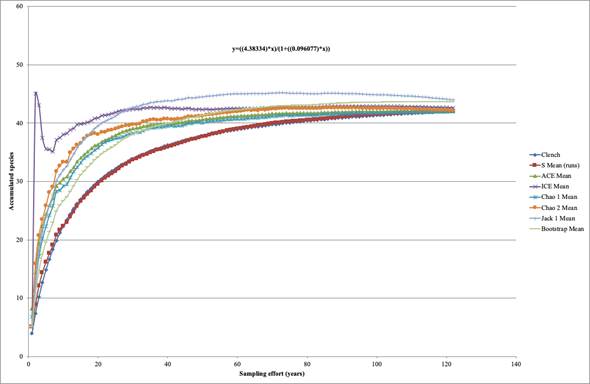
Figure 5: Species accumulation curves for the genus Dahlia Cav. based on 2297 herbarium specimens for 41 species collected between 1791 and 2021 as a measure of sampling effort. Values of total species richness provided by different species accumulation models (legend) converge upon similar total estimates of species richness.
Discussion
Dahlia is one of the most important genera in ornamental horticulture, with a remarkable diversity of cultivars throughout the world (TRHS, 2021). The current diversity of cultivars has been achieved in a relatively short time from hybridization between a few wild species, backcrosses, and subsequent intervarietal crossings (Wegner and Debener, 2008). A more complete knowledge of the natural diversity in Dahlia could enhance plant breeding programs, with important economic and cultural impacts on a global scale, because the possibilities of generating new cultivars are unquantifiable.
With the description of Dahlia gypsicola the number of species for the genus reaches 41. To cover the slight taxonomic shortfall presented here, we recommend further exploration in the centers of Dahlia species richness to increase scientific collections for rare and endangered species. Another priority is areas of high environmental complexity, for example, microenvironments generated by soils with a calcium and gypsum composition that support the presence of microendemic taxa (García-Mendoza et al., 2023). Mexico has extensive gypsum and calcic outcrops and hosts the most diverse gypsophile flora in the world but has been poorly studied (Ortiz-Brunel et al., 2023). Undescribed Dahlia species may exist where gypsophyte centers of endemism (Ortiz-Brunel et al., 2023) overlap with richness areas (Carrasco-Ortiz et al., 2019).

