











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Abies religiosa (HBK) Schl. & Cham. (oyamel) es una conífera nativa de las montañas del Eje Volcánico Transversal y la Sierra Madre del Sur de México (Rzedowski, 1978). Crece dentro de un intervalo altitudinal entre 2400 y 3600 msnm, en los estados de Hidalgo, Puebla, Veracruz, Michoacán, Jalisco, Morelos, Ciudad de México, Estado de México, Guerrero, Oaxaca y Tlaxcala (Sánchez-Velásquez et al., 1991). El bosque de oyamel que se ubica en la frontera del Estado de México y Michoacán, dentro de la Reserva de la Biósfera de la Mariposa Monarca (RBMM) funge como sitio de refugio y estancia invernal de las poblaciones de mariposa monarca Danaus plexippus L. (Anderson y Brower, 1996).
A pesar del esfuerzo y las prácticas de conservación dentro de la RBMM, existe preocupación por el futuro de Abies religiosa en la región, debido no sólo al cambio de uso de suelo (Champo-Jiménez et al., 2012; Vidal et al., 2014), sino porque como consecuencia del cambio climático se espera una reducción del nicho climático apropiado para su desarrollo. La reducción proyectada es de 69 % para la década centrada en 2030, 87 % para 2060 y 97 % para 2090, de manera que para finales de siglo el nicho climático del oyamel ya no existiría dentro de la RBMM (Sáenz-Romero et al., 2012).
Abies religiosa presenta diferenciación morfológica (Castellanos-Acuña et al., 2014) y genética (Ortiz-Bibian et al., 2017; Graciano-Ávila et al., 2019) entre poblaciones a lo largo del gradiente altitudinal, lo cual es un indicador de la adaptación local a nivel de poblaciones (Rehfeldt, 1988). Esto es especialmente importante para las especies de distribución alpina, donde los efectos del cambio climático se intensifican (Lenoir et al., 2008). Las poblaciones arbóreas quedarán desfasadas del clima que les es propicio (Rehfeldt et al., 2012), ya que éste ocurrirá a cada vez mayores altitudes en las zonas montañosas de México (Graciano-Ávila et al., 2019; Sáenz-Romero et al., 2010); por ello, la continuidad de las poblaciones naturales dependerá de la capacidad de adaptación (la que depende a su vez de la estructura y diversidad genética poblacional y plasticidad fenotípica de la especie), de la velocidad y capacidad de migración de la especie, así como de las barreras que impidan la migración de las poblaciones (Greenwood et al., 2014; Lenoir et al., 2008; Palmroth et al., 1999; Pigliucci et al., 2006).
Las coníferas que presentan una historia de vida longeva, como A. religiosa, se verán limitadas, tanto para su dispersión en altitud como disponibilidad de tiempo, para adaptarse a la velocidad de los cambios climáticos, ya que A. religiosa alcanza su madurez sexual entre los 17 y 25 años de edad (Madrigal-Sánchez, 1964; Com. Pers.) 1 , y es probable que posea una limitada capacidad de dispersión de sus semillas, no cuantificada para A. religiosa, pero en promedio 31 m para Abies alba Mill. (Cremer et al., 2012), lo que hace lento el proceso de migración natural.
Se ha sugerido realizar una migración asistida de las poblaciones de oyamel en la zona de estudio; ésto consistiría en recolectar semilla, producir planta en vivero y plantarla a una altitud mayor que la fuente de semillas, lo que significa desplazarse entre 275 (Sáenz-Romero et al., 2012) y 350 m (Ortiz-Bibian et al., 2017) a mayor altitud, lo que permitiría realinear las poblaciones al clima que les es propicio, pero que en el futuro ocurrirá a mayor altitud.
Para llevar a cabo lo anterior se requerirá una producción masiva de planta en vivero, por lo que el presente trabajo tuvo como objetivos: 1) evaluar la viabilidad y capacidad germinativa de las semillas de A. religiosa procedente de 15 poblaciones de oyamel ubicadas a lo largo de un gradiente altitudinal en el Cerro de San Andrés, Michoacán, México; 2) identificar la existencia de diferenciación entre poblaciones en la capacidad germinativa y 3) determinar con mayor precisión la cantidad de semilla que sería necesario recolectar para la producción suficiente en número de plantas en vivero.
MATERIALES Y MÉTODOS
Recolecta de semillas
En la última semana de noviembre de 2010 se recolectaron conos maduros de 11 árboles al azar de cada una de 15 poblaciones de A. religiosa en el Cerro de San Andrés, Michoacán, México, en un área cercana a la Reserva de la Biósfera de la Mariposa Monarca (RBMM), en un gradiente altitudinal de 2850 a 3550 msnm; cada sitio se ubicó a 50 m de diferencia altitudinal con respecto al sitio de recolecta siguiente. La ubicación geográfica se muestra en la Figura 1 y las coordenadas y caracterización ecológica en el Cuadro 1. Los árboles recolectados se ubicaron a una distancia mínima de 30 m entre cada uno, con la finalidad de disminuir las posibilidades de coancestría. A cada sitio de recolección se le denominó procedencia y al conjunto de árboles muestreados y representados en el experimento por semillas y plántulas se le denominó población.
Caracterización ecológica
Se realizó en sitios circulares de 1000 m2, un sitio por cada nivel de altitud; en ellos se registró el diámetro a la altura del pecho (DAP, con forcípula) y la altura con clinómetro (Marca Suunto, Vantaa, Finland) de las especies arbóreas presentes. A partir del DAP se estimó el área basal. Los árboles se agruparon en tres categorías: A. religiosa, Pinus sp.y hojosas (todos los árboles diferentes a las coníferas).
Manejo de la semilla
Los conos fueron secados durante 19 d en un invernadero de la Unidad San Juanito Itzícuaro del Instituto de Investigaciones Agropecuarias y Forestales de la Universidad Michoacana de San Nicolás de Hidalgo (UMSNH), en Morelia, Michoacán. Una vez secos los conos, las semillas se extrajeron manualmente, se mezcló la semilla por procedencia y se utilizó una cantidad equitativa de semillas de cada individuo. Debido a que las semillas de esta especie, una vez extraídas, presentan microgotas de resina en su exterior, la separación de semillas vanas y vacías resulta sumamente difícil, ya que las semillas se adhieren fácilmente entre sí, lo que dificulta tal separación mediante técnicas comunes, como la exposición a una corriente de viento o una columna de aire en un dispositivo de laboratorio; por ello, no se eliminaron las semillas vanas en esta etapa.

Pruebas de viabilidad y germinación
De cada una de las 15 poblaciones se tomaron 200 semillas al azar a las que se realizó una prueba de viabilidad mediante la prueba de tetrazolio (cloruro de 2,3,5-trifeniltetrazolio) (Sigma-Aldrich®) siguiendo la metodología descrita por la ISTA (2006). La muestra se dividió en cuatro repeticiones de 50 semillas cada una. A cada semilla se le efectuó un pequeño corte en el extremo contrario a donde emerge la radícula para facilitar el permeado del reactivo y se sumergieron en una solución de cloruro de 2, 3, 5-trifenil tetrazolio (0.8 mg 100 mL-1 de agua destilada) en la obscuridad. Después de 24 h, las semillas se sacaron de la solución y se realizó un corte transversal en cada una de ellas; si el embrión se teñía de color púrpura indicaba que el embrión era viable. El total de semillas examinadas se ajustó excluyendo las semillas que, al realizar el corte, se encontraban vacías.
Para el ensayo de germinación se utilizaron 400 semillas de cada población. Las 400 semillas se dividieron en ocho grupos de 50 semillas; cada grupo se colocó en una caja de Petri con papel filtro y se incubaron por un periodo de 28 d en una cámara de crecimiento (Lumistell®, Modelo ICP- 18, Celaya, Guanajuato, México), con ciclos de 12 h luz/12h obscuridad, a una temperatura de 25 °C. Las semillas se humedecieron cada 3 d con agua destilada con Captán® a una concentración de 0.8 g 100 mL-1. El criterio usado para determinar la germinación fue la emergencia de la radícula; los datos de germinación se tomaron cada tercer día.
Se determinó el peso de la semilla; para ello se tomaron al azar 400 semillas de cada una de las 15 poblaciones, se agruparon en ocho repeticiones de 50 semillas y se pesaron en una balanza electrónica con precisión de 0.001g (Ohaus SP402, Parsippany, NJ, USA), a fin de facilitar la estimación de la cantidad de semilla necesaria para producción de planta en vivero, tras considerar la capacidad germinativa.
Análisis estadístico
Para determinar diferencias estadísticas entre poblaciones en viabilidad, germinación y peso de semilla se realizó un análisis de varianza mediante el procedimiento GLM de SAS (SAS Institute, 2004). donde: µ es la media general, B. el efecto del i-ésimo bloque, Pj el efecto de la j-ésima procedencia y εijk es el error. Las procedencias se consideraron como un factor de efectos fijos, en cuanto a que la intención fue comparar los valores promedio de las poblaciones y determinar diferencias estadísticas entre ellas.
Yijk = µ + Bi + Pj + εijk
Cuadro 1 Ubicación geográfica de los sitios de recolecta del germoplasma, densidad de A. religiosa y área basal de ésta, de pinos, hojosas y total, por hectárea

Para determinar el patrón altitudinal de variación de viabilidad y capacidad germinativa se realizó un análisis de regresión (lineal y cuadrático) de los valores medios de las poblaciones (Yi) con respecto a su altitud (Xi), mediante el procedimiento REG de SAS (SAS Institute, 2004) con los siguientes modelos estadísticos: donde: Yi es valor medio de la i-ésima procedencia, β0 es el intercepto, β1 . β2 son los parámetros de regresión, Xi es la altitud de la i-ésima procedencia y εi el error aleatorio.
Yi = β0 + β1Xi + εi
Yi = β0 + β1Xi + β X 2 + εi
Se realizó una estimación de la cantidad de semilla (kg) necesaria para obtener 10,000 plántulas en vivero como promedio de poblaciones agrupadas en zonas altitudinales de acuerdo con la zonificación basada en características morfológicas (Castellanos-Acuña et al., 2014); se utilizó la fórmula reportada por la FAO (1991): donde: . es el número de kg a colectar para producir aproximadamente 10,000 plantas en vivero; S es el número promedio de semillas por kg en las poblaciones de una misma zona altitudinal; G es el porcentaje promedio de germinación en las poblaciones de una misma zona altitudinal, expresado como proporción; F es un factor de corrección (0.85), que considera un 15 % de mortalidad de planta en vivero, con base en experiencia previa.
Y = 10000 / (S G F)
RESULTADOS Y DISCUSIÓN
Viabilidad y germinación de la semilla
En el análisis de varianza de la viabilidad de la semilla se encontraron diferencias significativas entre poblaciones (P ≤ 0.0001). En la regresión entre la viabilidad promedio por población y la altitud de origen de la población, se observó que las poblaciones de la parte central de la distribución altitudinal (de 3100 a 3400 msnm) tienen un mayor número de semillas viables (hasta 57 %) que las de los extremos altitudinales superior e inferior (Figura 2); este patrón es estadísticamente significativo bajo el modelo de regresión cuadrático (R² = 0.510, P ≤ 0.01).
Los resultados de germinación fueron similares a los de viabilidad en cuanto a la tendencia de los datos entre procedencias. Se encontraron diferencias significativas entre poblaciones en el porcentaje de germinación (P ≤ 0.0001). El promedio general de germinación fue bajo (13%). Debe considerarse que este valor muy probablemente está subestimado, ya que, por las dificultades explicadas en metodología, no se eliminaron las semillas vanas antes de hacer la prueba de germinación.
La germinación promedio por población presenta un pronunciado patrón altitudinal bajo un modelo cuadrático (R² = 0.768, P = 0.0002), donde las poblaciones de la parte central de la distribución altitudinal (de 3100 a 3400 m de altitud) producen semillas con mayor porcentaje de germinación (20 % en promedio, con valores máximos de 27 %) que las poblaciones de los extremos altitudinales superior (3400 a 3550 msnm) e inferior (2850 a 3000 msnm), con un porcentaje muy bajo, de 9 y 5 % de germinación promedio, respectivamente (Figura 2).
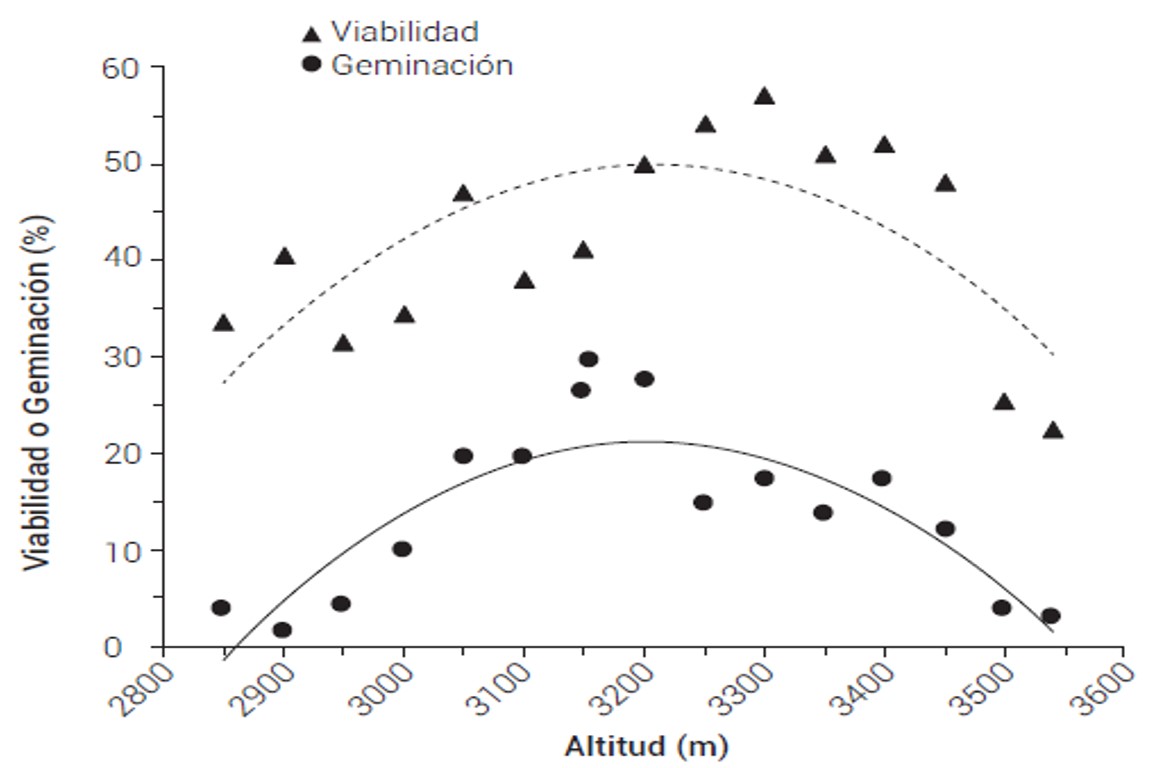
Figura 2 Porcentaje de viabilidad (R2 = 0.510; P = 0.0138) y germinación (R2 = 0.7680; P = 0.0002) promedio de la semilla por población de Abies religiosa en relación con la altitud de origen.
Las curvas de regresión de los porcentajes de viabilidad y de germinación son similares (Figura 2); esto indica que aun cuando la germinación está probablemente subestimada debido a que no se eliminaron las semillas vanas en la prueba, la estimación de la tendencia de la capacidad germinativa parece ser robusta; las procedencias de la parte central de la distribución altitudinal tienen más capacidad germinativa que las de los extremos altitudinales.
El peso promedio de semillas por kg también tuvo un patrón similar al de viabilidad y germinación; existieron diferencias significativas entre procedencias (P ≤ 0.0001), con un pronunciado patrón altitudinal, donde las procedencias de los extremos altitudinales tuvieron un menor peso que las de la parte central de la distribución altitudinal (modelo cuadrático, R² = 0.757, P = 0.0008, que excluye dos poblaciones atípicas).
El pronunciado patrón altitudinal encontrado podría deberse a que los árboles de las procedencias de los extremos altitudinales se encuentran expuestos a condiciones ambientales extremas. Las poblaciones de mayor altitud, entre 3500 y 3550 msnm, estarían expuestas a la ocurrencia frecuente de heladas, mientras que las del extremo altitudinal inferior probablemente lo están a estrés por sequía, ya que conforman el “límite xérico” (Mátyás, 2010). Probablemente el estrés ambiental en los límites altitudinales dificulta los procesos de formación de la semilla, tales como la polinización, fertilización y translocación de reservas para el llenado de la misma, como parece ocurrir con Pinus pseudostrobus (López-Toledo et al., 2017). El papel que juegan las condiciones ambientales en la fenología de las plantas ha sido ya reportado (Vitasse et al., 2009).
Si bien estudios de germinación realizados con Pseudotsuga menziesii (Mirb.) Franco en zonas boscosas fragmentadas y con poblaciones aisladas, reportan un indiscutible detrimento en el porcentaje de germinación de las semillas debido a la baja densidad del arbolado (Mápula- Larreta et al., 2008), en este caso la densidad de A. religiosa no parece haber influido de manera decisiva en el patrón altitudinal encontrado. No se encontró una asociación significativa entre la media por población de la viabilidad de las semillas de A. religiosa y las características del rodal, tales como densidad de individuos por hectárea (modelo lineal, R2 = 0.020, P = 0.6125) o área basal (modelo lineal, R2 = 0.007, P = 0.7600); tampoco se encontró asociación entre la germinación y la densidad de individuos (R2 = 0.080, P = 0.3068) o área basal (R2 = 0.093, P = 0.2694); sin embargo, estos datos deben tomarse con cautela, ya que la caracterización de los rodales se realizó tres años después de la recolecta de semillas y se encontraron evidencias de perturbación debido a tala, particularmente en sitios de baja altitud; por ello, la densidad de individuos y su área basal podría estar subestimada.
Los bajos niveles de viabilidad y germinación encontrados son congruentes con los reportados en otros estudios; por ejemplo, Nieto de Pascual-Pola et al. (2003) reportaron 37 % de viabilidad en semillas de A. religiosa de Río Frío, Ixtapaluca, México. En otro estudio realizado hace más de 30 años también se encontró un bajo porcentaje de germinación, de entre 45 y 49 % para esta especie, sin aplicar ningún tratamiento pre-germinativo y con semilla recién recolectada (Patiño-Valera et al., 1983).
Los bajos niveles de viabilidad y germinación también se han reportado para otras especies de Abies. En A. guatemalensis se encontró una germinación promedio de sólo 15% (INAB, 2017); si bien en ese estudio no se realizaron pruebas de viabilidad, necesariamente el porcentaje de semillas viables es mayor o igual al de germinación. Arista (1993) reportó 26 % de germinación en semillas de Abies pinsapo Boiss. en el Parque Natural Sierra de Grazalema, España. Al menos en el caso de las especies de Abies en México, ésto podría estar relacionado con que las poblaciones en general presentan un exceso de homocigotos, lo que indica altos niveles de endogamia, según análisis de isoenzimas (Eguiarte-Fruns y Furnier, 1997; Com. Pers.) 2 .
En cuanto a la velocidad de germinación, a los 7 d de que se inició el ensayo de germinación se registraron las primeras semillas germinadas. Las procedencias alcanzaron la mayor germinación acumulada al día 14; posteriormente, la germinación decreció de manera importante y después del día 21 ya no hubo germinación de semillas. Los datos de velocidad de germinación aquí obtenidos son en gran medida coincidentes con los de Mayen (1987; Com. Pers.) 3 , quien indica que el mayor porcentaje de semillas de Abies religiosa germina a los 12d. De acuerdo con los resultados de este trabajo, entre los días 12 y 14 se encontró la germinación máxima.
Clasificación de la calidad de semilla por zona altitudinal para su uso en vivero
Utilizando los límites definidos de zonas altitudinales para A. religiosa en la región estudiada con base en la diferenciación genética entre poblaciones (Ortiz-Bibian et al., 2017) es posible agrupar las poblaciones por zona altitudinal y estimar un promedio del número de kg de semilla necesarios para producir plantas en vivero. Esto permite simplificar el trabajo de planeación del esfuerzo necesario para colectar semilla para abastecer programas de reforestación a gran escala (Alía et al., 2005).
Para producir 10,000 plantas en vivero, si la semilla se recolecta de la zona de altitud intermedia (3000 a 3350 msnm) se requieren aproximadamente 2.0 kg; en cambio, la misma meta de producción en vivero requiere tres veces más semilla de la zona de mayor altitud (6.1 kg, 3350 a 3700 msnm) y casi cinco veces más semilla de la zona de menor altitud (11.1 kg, 2650 a 3000 msnm) (Cuadro 2).
La gran diferencia en semilla necesaria por cada zona altitudinal se debe principalmente a las diferencias en capacidad germinativa de las poblaciones, mas que al promedio del número de semillas por kg, que es muy semejante entre zonas, entre 32,708 y 33,916 semillas kg-1 (Cuadro 2) y relativamente cercana a la señalada en la ficha técnica de la especie (CONAFOR, 2013), de 26,599 semillas por kg y a los valores obtenidos por Nieto de Pascual-Pola et al. (2003), de 23,054 semillas por kg.
Cuadro 2 Cantidad de semilla necesaria por zona altitudinal para producir 10,000 plantas en vivero, en función del número de semillas kg-1, el porcentaje promedio de germinación y una pérdida en vivero estimada en 15 %

†Zonificación altitudinal según Ortiz-Bibian et al. (2017).
El resultado anterior indica las dificultades a las que se enfrentaría un programa de reforestación que contemple como un factor relevante el origen altitudinal de la semilla tras considerar o no el cambio climático. Si se considera únicamente que existe diferenciación genética entre poblaciones se podría reforestar usando semilla de la misma zona altitudinal que el sitio de plantación. Si adicionalmente a lo anterior se consideran los efectos del cambio climático sería necesario recolectar semilla en una zona altitudinal, producir planta en vivero y reforestar la zona altitudinal inmediata superior (Ortiz-Bibian et al., 2017). En ambos casos, la baja germinación de la semilla de la zona altitudinal inferior representará un reto para lograr el abasto de semilla a los viveros.
Limitaciones de este estudio
La cuantificación de semillas vanas y su eliminación de cada lote de semillas por procedencia previo a la germinación, indudablemente, habría sido de gran utilidad para explicar las diferencias en capacidad germinativa a lo largo del gradiente altitudinal. Tal análisis se recomienda para futuros trabajos a pesar de la dificultad que ocasiona la presencia de resina en los conos y semillas de esta especie.
CONCLUSIONES
Las poblaciones que se encuentran en la parte media del gradiente altitudinal de Abies religiosa en la región de la Reserva de la Biósfera de la Mariposa Monarca poseen el promedio más alto de viabilidad y de germinación de la semilla, en comparación con las poblaciones de los extremos altitudinales. Si se recolecta de la altitud intermedia (de 3000 a 3350 msnm) se requieren aproximadamente 2.0 kg de semilla para producir 10,000 plantas en vivero; en cambio, la misma meta requiere de casi cinco veces más semilla de la zona de menor altitud (2650 a 3000 m). Esto indica las dificultades que enfrentaría un programa de reforestación que contemple como un factor relevante el origen altitudinal de la semilla, ya que la baja capacidad germinativa de la semilla, especialmente de la zona altitudinal inferior, representa un reto para lograr el abasto de semilla a los viveros.

