










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkHidrobiológica
versión impresa ISSN 0188-8897
Hidrobiológica vol.11 no.1 Ciudad de México jun. 2001
Cambio estacional de clorofila a en la Bahía de La Paz, B. C. S., México
Aída Martínez-López,1 Rafael Cervantes-Duarte,1 Amada Reyes-Salinas1 y José Eduardo Valdez-Holguín1, 2
1 Centro Interdisciplinario de Ciencias Marinas, I. P. N. (CICIMAR-IPN). Av. Instituto Politécnico Nacional s/n. A. P. 592, La Paz, B. C. S. México. Becarios de la COFAA y EDD del I.P.N.
2 Depto. de Investigaciones Científicas y Tecnológicas, Universidad de Sonora (DICTUS). Rosales y Niños Héroes s/n, Hermosillo, Sonora, México.
Recibido: 26 de octubre de 2000
Aceptado: 18 de septiembre de 2000
Resumen
En el periodo abril de 1993 a marzo de 1995 se analizaron datos diarios de intensidad y dirección del viento, datos mensuales de temperatura hasta 25 metros, profundidad del disco de Secchi y la concentración de clorofila a, en superficie, 10 y 25 metros, en 5 estaciones ubicadas en la Bahía de La Paz. Los vientos presentaron una componente norte durante los meses de octubre a marzo y vientos dominantes del sur de abril a septiembre. Las temperaturas más altas ocurrieron durante julio-octubre entre 28 y 32 °C y las menores se registraron de enero a marzo entre 20 y 22 °C. Las mayores transparencias se observaron durante los meses de julio a noviembre, con valores entre 14 y 24 m; y las menores ocurrieron generalmente de enero a junio con profundidades entre 6 y 15 m. La clorofila a presentó un comportamiento estacional inverso a la temperatura y a la transparencia del agua. Las menores concentraciones de clorofila a integrada (< 10 mg m-2) ocurrieron en los meses más cálidos probablemente como resultado de la fuerte estratificación de la columna de agua, que no permite el transporte de material de la capa profunda a la capa superficial. Los mayores valores de clorofila a integrada (142.8 mg m-2) se presentaron en la época fría asociados con los procesos de mezcla de la columna de agua y probablemente con la disponibilidad de nutrientes en la zona eufótica.
Palabras clave: Clorofila a, fitoplancton, transparencia, cambio estacional, Bahía de La Paz, México.
Abstract
From April 1993 to March 1995 hourly wind data, intensity and direction, were analyzed. Monthly sampling of temperature, Secchi disc depth and chlorophyll a were carried out on 5 stations transect across the Bahía de La Paz at surface, 10 and 25 meters depth. Southward winds were dominant during October to March and northward the rest of the year. The highest temperatures were registered from July to October with temperatures between 28 and 32 °C, and the lowest from January to March, with values in the range of 20 and 22 °C. Maximum Secchi depths (14-24 m) was observed during July through November and minimum during January to June (6-15 m). Chlorophyll a concentration showed an inverse relationship with temperature and water transparency. The lowest integrated chlorophyll a values (<10 mg m-2) were found at warm season as a result probably of a strong water column stratification and slow vertical transport of dissolved and particulate materials from deep layers. Maximum values (142.8 mg m-2) during cold season were associated with water column mixing processes and nutrient availability in the euphotic zone.
Key words: Chlorophyll a, phytoplankton, transparency, seasonal change, Bahía de La Paz, Mexico.
Introducción
De los ambientes marinos, las zonas costeras son consideradas como áreas de un elevado interés socioeconómico porque son fuente importante de recursos naturales. Además, son utilizadas por los organismos como áreas de refugio, alimentación y desove. Por su alta productividad se pueden aprovechar para la práctica acuicultural y por sus características morfológicas son lugares ideales para las actividades recreativas. Bajo este contexto, estas zonas son potencialmente vulnerables a los cambios naturales y antropogénicos, por lo cual es preciso obtener un conocimiento básico de la ecología de sus diferentes ambientes.
En el océano, el fitoplancton representa el mayor porcentaje de la materia particulada y su abundancia está controlada principalmente por la luz, la temperatura, los nutrientes y el pastoreo, en cambio en las zonas costeras la dinámica hace mucho más complejo su análisis.
Los antecedentes de la hidrología de la Bahía de La Paz muestran una dinámica compleja con influencia de diferentes tipos de agua (Murillo-Jiménez, 1987; Reyes-Salinas, 1999). Por arriba de 100 m y a lo largo del año se encuentra agua cálida y salina (14-29 °C y > 35 ups) con características del tipo de agua del Golfo de California (Villaseñor-Casales, 1979; Jiménez-Illescas et al., 1994; Jiménez-Illescas, 1996; Zaytsev et al., 1998; Reyes-Salinas, 1999). Así mismo se ha mencionado la posible advección de agua superficial en verano, desde la costa oriental del golfo hacia el interior de la bahía (Signoret y Santoyo, 1980). Por otro lado Jiménez-Illescas (1996) ha reportado la presencia de giros ciclónicos en el invierno y principios de la primavera. La distribución vertical de temperatura indica el establecimiento de un marcado gradiente durante mayo-octubre y una homogeneización termal de la columna de agua el resto del año, lo cual permite una disponibilidad de nutrientes en la capa fótica como lo sugieren algunos estudios en donde se encontraron concentraciones mayores en invierno y principios de primavera y concentraciones menores en verano (García-Pámanes, 1981; Lavaniegos y López-Cortés, 1997; Reyes-Salinas, 1999). Respecto a la biomasa fitoplanctónica en la Bahía de La Paz las investigaciones son escasas. Estas no establecen claramente cuando se presentan los valores máximos y mínimos de la biomasa durante un ciclo anual, debido a una limitada cobertura temporal en esos estudios (Lavaniegos y López-Cortés, 1997; Reyes Salinas, 1999). Por lo tanto el objetivo del presente trabajo es describir los patrones temporales y espaciales de clorofila a en relación con los cambios de temperatura y transparencia del agua en la Bahía de La Paz.
Materiales y métodos
La Bahía de La Paz está localizada en la costa occidental del Golfo de California, entre los 24°06' y 24°47' de latitud norte y 110°18' y 110°45' de longitud oeste (Fig. 1). Tiene forma de óvalo, su eje mayor (81 km) está orientado de noroeste a sureste y su eje menor mide aproximadamente 33 km. La zona norte de la bahía es la más profunda, llega hasta 400 m. En la parte media la profundidad varía entre 180 y 270 m y en la parte sur es menor a 50 m (Cruz-Orozco et al., 1996). La temperatura mínima superficial en la bahía es de 20 °C en invierno-primavera y la máxima de 31 °C en verano (Espinoza y Rodríguez, 1987). La masa continental que delimita a la bahía presenta un clima muy seco y semicálido (Cruz-Ayala, 1996). La precipitación promedio anual es menor a 200 mm, siendo septiembre el mes más lluvioso (<100 mm). Los vientos predominantes del noroeste durante invierno tienen velocidades medias de 2 a 3 m s-1, en ocasiones alcanzan intensidades medias de 4 m s-1, las cuales son denominadas localmente "Collas". En verano, los vientos tienen una componente sur, con intensidades medias de 2 a 3 m s-1 (Robles Gil-Mestre, 1998). La temperatura promedio ambiental mínima es de 8 °C en invierno y la máxima de 37 °C en verano (Cruz-Ayala, 1996).

Con el propósito de describir de manera general la variabilidad del patrón estacional de vientos y de la biomasa fitoplanctónica, se obtuvieron registros horarios de viento (dirección y velocidad) para el periodo de enero de 1993 a marzo de 1995, del Observatorio Meteorológico de la Comisión Nacional del Agua. La estación está ubicada a una altura de 18.50 metros sobre el nivel del mar (Fig. 1). A los datos horarios del viento se les aplicó un promedio móvil de 24, para filtrar los efectos diurnos.
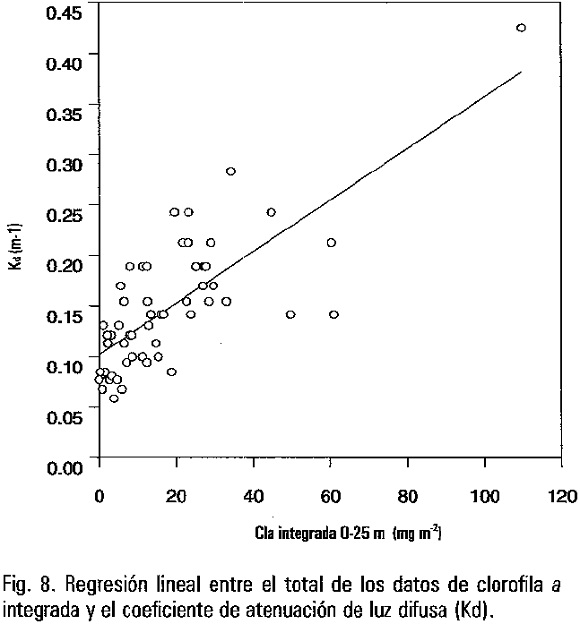
En cinco estaciones establecidas en la Bahía de La Paz en sentido sureste-noroeste se tomaron muestras mensualmente de abril de 1993 a marzo de 1995 (Fig. 1). En cada estación se determinó la temperatura, la profundidad de desaparición del disco de Secchi (PS) y se tomaron muestras a profundidades de 1, 10 y 25 m para determinar la concentración de clorofila a. La temperatura se midió con un termómetro de cubeta con una resolución de ±0.1 °C. A partir de las PS se calculó el coeficiente de atenuación vertical de luz difusa (Kd = 1.7/PS), y la profundidad de la zona eufótica (PZE) (Kirk, 1994). Para la determinación de clorofila a, las muestras colectadas se colocaron en una hielera, en donde se mantuvieron aproximadamente a 4 °C hasta que se concluyó el muestreo y fueron transportadas al laboratorio. Posteriormente se filtró un volumen de agua conocido, generalmente entre 1 y 1.5 litros a través de filtros GF/F de 25 mm de diámetro. La filtración se realizó al vacío a una presión aproximada de un tercio de atmósfera. Los filtros fueron congelados a -30 °C hasta su extracción en acetona al 90% por 24 horas (Venrick y Hayward, 1984) y posteriormente analizados espectrofotométricamente. Los cálculos para la determinación de clorofila a se hicieron de acuerdo a las ecuaciones de Jeffrey y Humphrey (1975). Con el propósito de estimar el efecto de la contribución del fitoplancton a la atenuación de luz, se relacionó la concentración de clorofila a integrada de 0 a 25 m y el coeficiente de atenuación de luz difusa.
Resultados
Se observó que los vientos presentaron una fuerte componente norte sur, mientras que los vientos este oeste fueron muy débiles con una variación mínima (Fig. 2A, B). Los vientos del norte fueron predominantes durante los meses de noviembre a marzo, con intensidades promedio de 2 m s-1 en 1993 e incrementándose hasta 4 m s-1 en 1995. Se presentaron vientos dominantes del sur de abril a octubre, con intensidades promedio de 2 m s-1 durante 1993 y de 4.5 m s-1 en 1994. Los datos en conjunto mostraron una fuerte variabilidad diurna, no mostrada debido a que los datos se filtraron para eliminar la variabilidad menor a 24 horas.

En la figura 3A se presenta la variabilidad de la temperatura en la estación 4 como representativa del patrón anual de las diferentes localidades, las cuales presentaron diferencias en un mismo mes menores a 2 °C. Las temperaturas más altas ocurrieron durante julio-octubre entre 28 y 32 °C, con diferencias entre superficie y 25 metros hasta de 4 °C. Las menores temperaturas generalmente se registraron de enero a marzo entre 20 y 22 °C. En este mismo periodo la temperatura desde la superficie hasta 25 metros fue muy similar. Estas condiciones indican los periodos de estratificación y mezcla de la columna de agua.

Las profundidades de desaparición del disco de Secchi entre las diferentes estaciones en un mismo muestreo fueron heterogéneas. Se presenta la variación estacional promedio y desviación estándar de las 5 estaciones (Fig. 3B). Las mayores transparencias se observaron durante los meses de julio a noviembre, con valores 14 y 24 m; y las menores ocurrieron generalmente de enero a junio con profundidades entre 6 y 15 m.
En general, se observó que la temperatura superficial y el coeficiente de atenuación vertical de luz difusa (Kd) se asociaron directamente, Kd = 0.36 -0.009 T°C. El análisis por grupos de estaciones demostró que la máxima correlación se obtiene al agrupar las estaciones 3, 4 y 5 (r2=0.57; p<0.05) (Fig. 4). La correlación disminuye hasta en 50% cuando se incluyen las estaciones 1 y 2.

La distribución de la concentración de clorofila a en sus tres profundidades fue similar, las mayores concentraciones se registraron en los niveles de 10 y 25 m, principalmente en las estaciones 3, 4 y 5 (Fig. 5). En las estaciones 1 y 2 se observó una mayor variabilidad en la distribución de clorofila a. Las máximas concentraciones (10.9 mg m-3) se presentaron durante abril de 1994, en la estación 5. Los valores disminuyeron de julio a noviembre, y las mínimas concentraciones fluctuaron desde indetectables hasta menores a 1 mg m-3 (excepto en la estación 1 en el mes de septiembre de 1993).

La distribución de clorofila a en sus diferentes niveles (Fig. 6). En general, las concentraciones durante los meses de diciembre a abril fueron mayores a 10 mg m-2, y durante junio a noviembre menores a este valor. Las concentraciones más altas se estimaron en los meses de diciembre a abril de 1994, con máximos de 142.8 mg m-2.

El comportamiento general de las cinco estaciones mostró una relación inversa entre la temperatura y la clorofila a superficial, es decir, cuando la temperatura se incrementó la clorofila a disminuyó (Fig. 7). De manera particular se observaron algunas discrepancias en los meses en que la temperatura del agua se incrementó rápidamente, asociados con aumentos en la concentración de clorofila a.
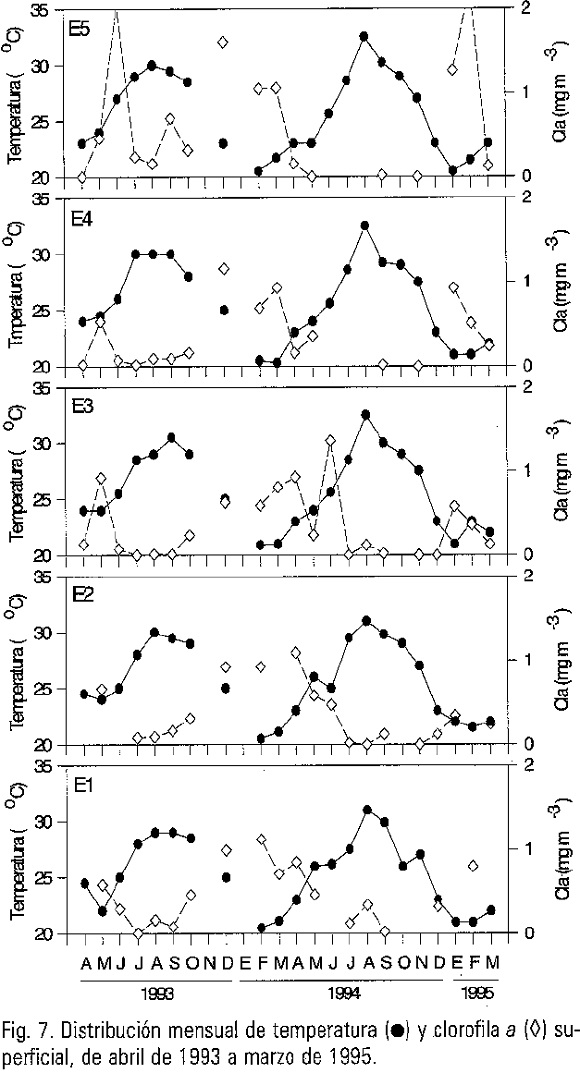
El análisis de correlación entre el total de los datos de clorofila a integrada y el coeficiente de atenuación de luz difusa fue de r2=0.57; (p<0.05) (Fig. 8). En forma particular, para cada estación de muestreo la r2 varió de 0.56 a 0.78 (p<0.05), con excepción de la estación 5, r2=0.14; (p<0.05).
Discusión
En el Golfo de California, durante invierno los vientos provienen del noroeste y en verano del sureste (Roden, 1964). Localmente en el área de La Paz se registran vientos intensos del noroeste, en ocasiones con una duración promedio de 2-3 días principalmente a finales del invierno (Robles Gil-Mestre, 1998). Durante estos periodos se ha reportado en la bahía una capa de mezcla de aproximadamente 20 metros de profundidad (Villaseñor-Casales, 1979; Lavaniegos y López-Cortés, 1997). Sin embargo, el efecto de mezcla por viento en esta época debe ser evaluado, ya que ni la intensidad (promedio de 4 m s-1) ni la persistencia (2-3 días) pueden generar una mezcla tan profunda, posiblemente esta mezcla de la columna de agua durante noviembre a marzo sea también el resultado de movimientos convectivos. Durante el periodo de abril a octubre, aún con vientos más intensos, éstos no tienen la suficiente energía para producir una capa de mezcla similar a la del periodo de noviembre a marzo debido a la intensa estratificación (Δt=4°C en 25 m de profundidad).
La variación estacional de la temperatura en las profundidades de 0, 10 y 25 m en las diferentes estaciones fue similar, esto señala que existe una homogeneidad espacial en esta zona de la bahía. Se observaron temperaturas promedio altas en la capa superficial (0-25 m) de julio a noviembre y bajas de enero a mayo, con cambios pronunciados de una temporada cálida a una fría. La temporada cálida (28.4±1.9°C) se extiende de junio a noviembre y la fría de diciembre a mayo (22.5±4.4°C). Las temporadas encontradas tienen cierta semejanza con las propuestas por Robles-Pacheco y Marinone (1987) para el Golfo de California, en donde, el verano se extiende de junio a octubre, el invierno de diciembre a abril, y noviembre y mayo son periodos de transición. Las diferencias entre los valores de superficie y 25 m muestran los periodos de estratificación y mezcla de la columna de agua. Este calentamiento que origina una estratificación en la temporada cálida es debido a una intensa irradiación solar. Las condiciones anteriormente descritas confieren dentro del contexto del ciclo anual un carácter oligotrófico a la bahía, el cual se puede acentuar por la intrusión de Agua Superficial Ecuatorial (Torres-Orozco, 1993; Lavín et al., 1997; Reyes-Salinas, 1999).
Los valores promedio de la transparencia del agua presentaron un cambio estacional, las mayores profundidades de desaparición de disco de Secchi se registraron cuando la temperatura del agua fue más alta y viceversa. La estratificación de la columna de agua en la temporada cálida, disminuye la cantidad de material particulado en la capa superior de la termoclina por la disminución en la producción biológica y el flujo vertical. Por otro lado, las características oligotróficas del Agua Superficial Ecuatorial que invaden la bahía tienen una menor cantidad de material suspendido. Los vientos del sureste durante esta época, con una velocidad media de 3 m s-1, son incapaces de producir un efecto sobre los procesos biológicos (Therriault y Platt, 1981). La disminución de la temperatura asociada al régimen de frentes fríos y vientos del noroeste, producen una intensa mezcla vertical (por convección o por el esfuerzo del viento) y un aumento de nutrientes en la capa superficial. Como consecuencia se incrementa el material particulado, con una disminución en la transparencia del agua.
En la Bahía de La Paz se han sugerido varios procesos de fertilización de la zona eufótica. Jiménez-Illescas (1996) y De Silva-Dávila y Palomares-García (1998) reportan la posible existencia de giros ciclónicos en la parte central de la bahía durante invierno y principios de primavera, los cuales pudieran concentrar las comunidades planctónicas. Signoret y Santoyo (1980) mencionan que los florecimientos del fitoplancton que encontraron en otoño-invierno están en correspondencia con los pulsos de intensa fertilización en el golfo, como resultado de las sugerencias que se regeneran en la costa oriental. La distancia entre ambas costas (200 km) indica que este transporte es poco probable. Asimismo, Gárate-Lizárraga y Martínez-López (1997) proponen que los procesos físicos cerca de las islas Cerralvo y San José crean condiciones óptimas para el florecimiento del fitoplancton, principalmente para el desarrollo de mareas rojas incluyendo las del protozoario Mesodinium robrum (Lohmann), como fue el caso en abril de 1994, en donde las máximas concentraciones de clorofila (hasta 10.9 mg m-3) se presentaron cercanas al canal de San José. En este trabajo se sugiere un fuerte acoplamiento entre los regímenes climáticos (frío-cálido), vientos, la estructura vertical de la columna de agua y la abundancia de fitoplancton. Durante periodos cálidos, una fuerte estratificación influenciada por una fuerte irradiación solar limita el flujo de nutrientes a la zona eufótica disminuyendo la producción fitoplanctónica. En la época fría, la incidencia de frentes fríos y vientos, incrementan el flujo de nutrientes hacia la zona eufótica y promueven el aumento de la biomasa de fitoplancton.
Esta hipótesis está acorde al patrón general de la variación estacional observada en la biomasa fitoplanctónica (clorofila a) para las zonas subtropicales, en donde el fitoplancton nunca se encuentra limitado por su luz sino por nutrientes (Longhurst, 1995). La tasa de producción primaria se incrementa a través del invierno y alcanza su máximo en primavera antes de que los nutrientes sean limitantes. Mientras que en verano, la estratificación de la columna de agua favorece la formación de un máximo subsuperficial o profundo de clorofila (Longhurst, 1995). En este trabajo, durante la temporada fría (periodo de mezcla) la clorofila a (por volumen o integrada) fue mayor o similar a reportes previos (Lavaniegos y López-Cortés, 1997; Reyes-Salinas, 1999). Por otro lado, para el periodo de estratificación (estación cálida), los valores de clorofila fueron menores. La serie de datos utilizada no permitió complementar el esquema del patrón para zonas subtropicales debido a que solo se registró la mitad de la zona eufótica. Sin embargo, Reyes-Salinas (1999), encuentra por medio de perfiles de fluorescencia natural máximos subsuperficiales, con lo cual es posible caracterizar el patrón encontrado con el de zonas subtropicales. También es similar al reportado para la parte central del Golfo de California, en donde se presentan pronunciados cambios estacionales en las condiciones hidrográficas y climáticas asociados al cambio en la abundancia de plancton (Thunell et al., 1994; Santamaría del Ángel et al., 1994; Thunell et al., 1996). López-Verdín (1999) encontró que durante verano existe la formación de un máximo profundo de clorofila a, con concentraciones superficiales menores a 0.3 mg m-3, en la parte central del golfo causado por una fuerte estratificación, mientras que en invierno, la mezcla de la columna de agua promueve la formación de un máximo subsuperficial con contenidos promedio de 2 mg m-3.
Teóricamente existe una relación entre el incremento en la concentración de partículas y la PS (Megard y Berman, 1989). La turbidez se puede incrementar principalmente por la resuspensión y transporte de partículas, el aumento de la biomasa del plancton, la actividad de pastoreo y la agregación de bacterias alrededor de la materia orgánica. Con la excepción de la estación 1 y 2 la transparencia del agua puede ser parcialmente explicada por la concentración de clorofila a integrada de 0 a 25 m. La estación 1 se localiza en la zona somera de la bahía y se considera que pueda estar sujeta a los efectos de la resuspensión de sedimentos por las corrientes de marea y viento. Asimismo, la estación 2 está situada en un canal más angosto que comunica el golfo con la bahía. En esa zona se han registrado velocidades de corriente de entre 0.3 a 0.6 m s-1 (Jiménez-Illescas, 1996) las cuales pueden generar turbulencia y resuspender el material del fondo. Con base en la relación calculada entre temperatura superficial y Kd (r2=0.59; p<0.05), es posible predecir la profundidad de la zona eufótica, con información de temperatura superficial estimada con sensores remotos. Con la finalidad de hacer estimaciones de la productividad total de la columna de agua, estos sensores remotos también nos proporcionan estimaciones de clorofila a superficial (1 profundidad óptica = 1/Kd; Kirk, 1994). Sin embargo, entre otras propiedades se tiene que caracterizar la forma del perfil de biomasa de fitoplancton. En la bahía es evidente que durante la temporada cálida existe la formación de un máximo profundo de clorofila a. A partir de la información de Kd, la PZE durante esta época del año es en promedio de 50 m, por lo que en este trabajo dejamos sin registrar la zona productiva y probablemente el máximo haya estado dentro de esta región. Sin embargo, durante invierno la mayor parte de la zona productiva (30 m de profundidad) fue muestreada y la forma del perfil mostró que el máximo se desarrolla comúnmente a 25 m de profundidad. Esta información es básica para la aplicación de los datos de sensores remotos en el medio marino, ya que con mediciones de temperatura mediante el sensor AVHRR (A Very Resolution Radiometer) se puede estimar el Kd, y con la relación Kd y clorofila predecir la biomasa fitoplanctónica.
Agradecimientos
Agradecemos a la M. C. Diane Gendron y a su grupo de estudiantes (CICIMAR-IPN) por su valiosa ayuda para la obtención de las muestras y datos de campo de este estudio. Este trabajo fue desarrollado dentro del programa de investigación "PRODUCCIÓN BIOLÓGICA DE LA PENÍNSULA DE BAJA CALIFORNIA", el cual fue auspiciado por el CONACyT y por la Dirección de Estudios de Posgrado e Investigación como parte del proyecto "ECOLOGÍA DEL PLANCTON" (IPN-DEPI 931420).
Literatura citada
Cruz-Ayala, M. B., 1996. Variación espacio-temporal de la ficoflora y su abundancia relativa en la Bahía de La Paz, B. C. S., México. Tesis de Maestría. CICIMAR-IPN. 100 p. [ Links ]
Cruz-Orozco, R., C. Martínez-Noriega y A. Mendoza-Maravillas, 1996. Batimetría y sedimentos de la Bahía de La Paz, B. C. S., Oceánides 11(1): 21-27. [ Links ]
De Silva-Dávila, R. y J. R. Palomares-García, 1998. Unusual larval growth production of Nyctiphanes simplex in Bahía de La Paz, Baja California Sur, México. Journal of Crustacean Biology 18(3): 9 p. [ Links ]
Espinoza, J. y H. Rodríguez, 1987. Seasonal phenology and reciprocal transplantation of Sargassum sinicola Setchell et Gardner in the Southern Gulf of California. Journal Experimental Marine Biology and Ecology 110: 183-195. [ Links ]
Gárate-Lizárraga, I. y A. Martínez-López, 1997. Red tides of the protozoan Mesodinium rubrum (Lohmann) Kahk in Bahía de La Paz. Boletín No. 53, CICIMAR-IPN. [ Links ]
García-Pámanes, J., 1981. El fitoplancton de la porción oriental de la Bahía de La Paz, B. C. S., durante primavera y verano. pp. 201-218. En: Resúmenes y Progama, VII Simposium Latinoamericano sobre Oceanografía Biológica, Acapulco, Guerrero, México. 15-19 de noviembre. [ Links ]
Jiménez-Illescas, A. R., 1996. Análisis de Procesos Barotrópicos y Baroclínicos en la Bahía de La Paz, B. C. S. Tesis de Doctorado. UNAM. 193 pp. [ Links ]
Jiménez-Illescas, A. R., M. Obeso-Nieblas, M. A. Alatorre-Mendieta, S. Troyo-Diéguez y H. García-Escobar, 1994. Hidrología de la Bahía de La Paz. Oceanología 1(2): 115-131. [ Links ]
Jeffrey, S. W. y G. F. Humphrey, 1975. New spectrophotometric equations for determining chlorophylls a, b and c, in higher plants, algae and natural phytoplankton. Biochemie und Physiologie der. Pflanzen 167: 191-194. [ Links ]
Kirk, J. T. O., 1994. Light and photosynthesis in aquatic ecosystems. Segunda Edición, Cambridge University Press, Cambridge, 509 p. [ Links ]
Lavaniegos, B. E. y D. López-Cortés, 1997. Fatty acid composition and community structure of plankton from the San Lorenzo Channel, Gulf of California. Estuarine Coastal and Shelf Science 45: 845-854. [ Links ]
Lavín, M. F., E. Beier y A. Badan, 1997. Estructura hidrográfica y circulación del Golfo de California: Escalas estacional e interanual. pp. 141-171. En: M. F. Lavín (Comp.). Contribuciones a la Oceanografía Física en México, Monografía No. 3, Unión Geofísica Mexicana. [ Links ]
Longhurst, A. R., 1995. Seasonal cycles of pelagic production and consumption. Progress in Oceanography 36: 77-167. [ Links ]
López-Verdín, C., 1999. Variabilidad de la productividad orgánica primaria en el Golfo de California. Tesis Profesional Químico-Biólogo. Universidad de Sonora. 80 p. [ Links ]
Megard, R. O. y T. Berman, 1989. Effects of algae of the Secchi transparency of the Southeastern Mediterranean Sea. Limnology and Oceanography 34(8): 1640-1655. [ Links ]
Murillo-Jiménez, J. M., 1987. Algunas características paleoceanográficas y cuerpos de agua inferidos a partir del registro micropaleontológico (Radiolaria) en la Bahía de La Paz, Baja California Sur, México. Tesis Profesional. UABCS, 70 pp. [ Links ]
Reyes-Salinas, A., 1999. Factores que controlan la productividad primaria en la Bahía de La Paz, B. C. S. Tesis de Maestría. CICIMAR. 122 pp. [ Links ]
Robles-Pacheco, J. M. y S. G. Marinone, 1987. Seasonal and interannual thermohaline variability in the Guaymas Basin in the Gulf of California. Continental Shelf Research 7: 715-733. [ Links ]
Robles Gil-Metre, S., 1998. El clima de la ciudad de La Paz. Tesis de Maestría. UNAM. 233 pp. [ Links ]
Roden, G. I., 1964. Oceanographyc aspects of the Gulf of California. pp. 30-58. En: T. J. H. Van Andel y G. G. Shor (Comps.). Marine Geology of the Gulf of California: A Symposium. AAPG, Memoir 3. [ Links ]
Santamaría del Ángel, E., S. Álvarez-Borrego y F. E. Müller-Karger, 1994. Gulf of California biogeographic regions based on Coastal Zone Color Scanner imagery. Journal of Geophysical Research 99(C4): 7411-7421. [ Links ]
Signoret, M. y H. Santoyo, 1980. Aspectos ecológicos del plancton de la Bahía de La Paz, Baja California Sur. Anales del Instituto de Ciencias del Mar y Limnología, UNAM, 7(2): 217-248. [ Links ]
Terriault, J. C. y T. Platt, 1981. Environmental control of phytoplankton patchiness. Canadian Journal of Fisheries and Aquatic Sciences 38: 638-641. [ Links ]
Thunell, R., C. Pride, P. Ziveri, F. Müller-Karger, C. Sancetta y D. Murray, 1996. Plankton response to physical forcing in the Gulf of California. Journal of Plankton Research 18(11): 2017-2026. [ Links ]
Thunell, R., C. Pride, E. Tappa y F. Müller-Karger, 1994. Biogenic silica fluxes and accumulation rates in the Gulf of California. Geology 22: 303-306. [ Links ]
Torres-Orozco, E., 1993. Análisis volumétrico de las masas de agua del Golfo de California. Tesis de Maestría. CICESE. 80 p. [ Links ]
Venrick, E. y T. Hayward, 1984. Determining chlorophyll on the 1984 CALCOFI surveys. California Cooperative Oceanic Fisheries Investigations Report, 25: 74-79. [ Links ]
Villaseñor-Casales, A., 1979. Distribución vertical de temperatura, salinidad y oxígeno disuelto en la Bahía de La Paz, B. C. S., durante la primavera de 1976, California Coopetarive Oceanic Fisheries Investigations Report 20: 146-149. [ Links ]
Zaytsev, O. V., F. Salinas-González y S. Troyo-Diéguez, 1998. Water dynamics and mixing studies in the coastal zone of the Gulf of California. Proceedings of Canadian Hydrographic Conference, 267-276. [ Links ]

