










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkHidrobiológica
versión impresa ISSN 0188-8897
Hidrobiológica vol.22 no.1 Ciudad de México ene./abr. 2012
Artículos
Composición de la comunidad íctica de la Cuenca del Río Santiago, México, durante su desarrollo hidráulico
Fish community composition of The Santiago River Basin, Mexico, during its hydraulic development
María Antonieta Gómez–Balandra,1,2 Edmundo Díaz–Pardo2 y Altagracia Gutiérrez–Hernández2
1 Instituto Mexicano de Tecnología del Agua. Paseo Cuauhnáhuac 8532, Col. Progreso, Jiutepec, Morelos. 62550. México. E–mail: magomez@tlaloc.imta.mx
2 Universidad Autónoma de Querétaro. Facultad de Ciencias Naturales, Doctorado en Recursos Bióticos. Av. de las Ciencias s/no. Col. Juriquilla, Querétaro. 76230. México.
Recibido: 1 de marzo de 2011.
Aceptado: 20 de noviembre de 2011.
RESUMEN
La cuenca del río Santiago en el Noroeste de México, es una región con importante diversidad íctica en Norteamérica. Con el objetivo de identificar cambios temporales y espaciales de la comunidad íctica durante tres etapas de desarrollo hidráulico del río, entre las cuales existieron distintos grados de conectividad, desde su origen hasta su desembocadura, en 1032 registros de bases de datos nacionales e internacionales de 132 localidades y cinco subcuencas se reconocieron 44 especies; 13 endémicas de la región Lerma–Chapala–Santiago, 22 nativas de México, 7 introducidas y 2 traslocadas. Treinta y tres especies correspondieron a la primera etapa, limitada a 1970. En la segunda etapa considerada de 1971 a 2000, fueron 32 especies y en la última (2001 a 2006), sólo 25 taxa. Por su abundancia relativa, 6 especies representaron el 71% de las colectas y 22 tuvieron una amplitud de distribución entre 5 y 36%. Aún con las diferencias en periodos de desarrollo hidráulico, métodos y localidades de colecta, debido a su ausencia temporal y reducción del hábitat en sus intervalos de distribución altitudinal, aparentemente se han perdido ocho especies endémicas y 17 se hallan en riesgo. En conjunto, el deterioro ambiental y la fragmentación han disminuido el número de especies nativas y endémicas y por el contrario incrementado las introducidas, principalmente en los ríos Santiago y Verde. Por sus condiciones de conservación y posible presencia de poblaciones aisladas, se propone un seguimiento continuo en los ríos Huaynamota y Bolaños para preservar áreas representativas de la comunidad íctica de la cuenca.
Palabras clave: Peces, cambios temporales, cuenca Río Santiago, México.
ABSTRACT
The Santiago River basin located in northwest Mexico is one region with important fish diversity in North America. The objective of this paper is to identify temporal and spatial changes in the fish community over three stages of the river hydraulic development, when certain degrees of connectivity existed, from its origin to its sea mouth. 1032 fish records from national and international databases belonging to132 localities in five sub–basins were analyzed. In total, 44 species were recognized, 13 endemic of the Lerma–Chapala–Santiago region, 22 natives to Mexico, 7 introduced and 2 translocated. In the first stage up to 1970, 33 species were gathered, mainly in the upper part of the basin. In the second stage from 1971 to 2000, 32 species were registered, and in the last stage (2001 to 2006), only 25 taxa. According to their relative abundance six species represented 71% of the total collection and 22 had distributional amplitude between 5% and 36%. Even though differences in periods of time for each development stage, catching methods and localities, due to their temporal absence and habitat reduction in their distributional range, eight species are apparently extirpated and 17 species are at risk in the Santiago River. Overall environmental degradation and fragmentation have reduced native and endemic species and an increase in the introduced ones have been observed mainly in the Santiago and Verde rivers. Based on their conservation degree and potential presence of isolated fish populations, a follow–up monitoring program is proposed in the Huaynamota and Bolaños rivers to preserve representative areas of the fish community in the basin.
Key words: Fish, temporal changes, Santiago River Basin, Mexico.
INTRODUCCIÓN
En los últimos 150 años, las presiones sobre los sistemas fluviales se han incrementado en muchas regiones del mundo debido al crecimiento demográfico, cambios en el uso del suelo, descargas residuales y construcción de infraestructura hidráulica (Pegg & McClelland, 2004; Vörösmarty et al., 2010; Garrido et al, 2010). La extracción, almacenamiento, trasvase y derivación masiva de agua de los ríos para distintos usos, han alterado características ambientales de conectividad y hábitat (FAO, 2001; Quist et al., 2005; Anderson et al., 2006), provocando la reducción de las poblaciones de organismos que viven en ellos y cambiando la estructura de las comunidades, llevando incluso a la perdida de las especies sensibles (Schlosser & Agermeier, 1995; WCD, 2000; Falke & Gido, 2006; Drástik et al., 2008). Las comunidades de peces exhiben cambios aguas arriba y abajo de los embalses, que afectan sobre todo a los peces migratorios al interrumpir sus ciclos de vida (McCarthy et al., 2008). En el caso de las centrales hidroeléctricas, las oscilaciones diarias de caudal aguas abajo, afectan el hábitat y la estructura de las comunidades acuáticas (Drástik et al., 2008; Birgitta et al., 2010). Por otro lado, el desarrollo de las pesquerías con especies introducidas, tolerantes y depredadoras en los embalses, ha tenido también severos efectos en la composición ictica (Lyons et al., 1995, 1998; Lyons & Mercado–Silva, 1999; Guzmán–Arroyo et al., 2009).
En general, se considera que el análisis histórico de las comunidades biológicas proporciona criterios para interpretar la presencia o ausencia de las especies, asociándola con los agentes causales que han modificado su abundancia y distribución, como son las alteraciones al régimen de caudal del río, el deterioro de la calidad del agua y la introducción de especies, entre las más importantes (Lyons et al., 1998; Hughes et al., 2005; Abell et al., 2008; Guzmán–Arroyo et al., 2009; Mercado–Silva et al., 2006, 2009; Penczak et al., 2009).
En la cuenca del Río Santiago existen aproximadamente 189 presas de almacenamiento, derivadoras y de generación de energía eléctrica (Chiapa & Marengo, 2008; Conagua–IMTA, 2009). La mayoría se ubican en el río Verde (131), mientras que en el área de estudio sobre el río Santiago se ubican 11 actualmente en operación (Fig. 1). En la Tabla 1, se señala el año de construcción y la ubicación altitudinal de cada una para reconocer los períodos en los que el río mantuvo cierta conectividad. Su distribución en un perfil longitudinal se señala en la Figura 2.

El desarrollo hidráulico de la cuenca del río Santiago ha sido muy intenso y prolongado, ya que se inició a mediados del siglo XIX (Chiapa & Marengo, 2008; Conagua–IMTA, 2009) y aún continúa. Entre las diversas presas instaladas, sobresalen las construidas en los últimos 50 años por la altura de su cortina y su tamaño, así como por los efectos de la generación hidroeléctrica y su ubicación en cascada. Actualmente se ha incrementado la presión por el agua y la construcción de presas para abastecimiento público, principalmente en el río Verde.
Los efectos de la fragmentación, pérdida del hábitat e introducción de especies exóticas, han sido poco estudiados en esta cuenca, aunque en sistemas similares se ha reconocido que los efectos acumulativos modifican las redes tróficas (Gutiérrez–Hernández, 2003; Contreras–Balderas et al., 2008; Mercado–Silva et al., 2009), incrementando, el riesgo de pérdida o extinción de especies de peces (López–López & Díaz–Pardo,1991; Díaz–Pardo et al., 1993; Contreras–Balderas, 2000; De la Vega–Salazar et al., 2003; Birgitta et al., 2010).
En términos generales, la información disponible está referida como listados o inventarios de especies presentes en distintos ríos, incluyendo, en algunos casos, sus atributos principales (Guzmán–Arroyo & Lyons, 2003; Mercado–Silva & Lyons, 2004; Miller et al., 2005). Por lo tanto, se carece de una recopilación y análisis de los posibles cambios históricos cualitativos y cuantitativos que han ocurrido en la cuenca del río Santiago. Por esta razón, en este trabajo se analiza la variación espacial y temporal de la composición de la comunidad íctica de la cuenca durante tres etapas de su desarrollo hidráulico, en las que existieron distintos grados de conectividad entre los tributarios y la corriente principal, hasta su desembocadura.
MATERIALES Y MÉTODOS
La cuenca del río Santiago se localiza en el centro–occidente de México, enmarcada por las coordenadas 23° 24' 36" y 20° 18' 03" N, –101° 16' 48" y –105° 28' 12" O. Abarca porciones de seis entidades federativas: Durango, Zacatecas, Aguascalientes, Guanajuato, Jalisco y Nayarit (Fig. 3).
La delimitación del área de estudio en la corriente principal, se consideró a partir del origen del río Santiago al noreste del lago de Chapala (1530 msnm) y hasta 10 km aguas abajo de la presa San Rafael (64 msnm), con un recorrido aproximado de 440 km (señalado entre líneas en la Fig. 3).
El río Santiago recibe sus principales afluentes por la margen derecha y corresponden a escurrimientos provenientes de altitudes que rebasan los 2500 msnm, divididos en las subcuencas de los ríos Verde, Juchipila, Bolaños y Huaynamota. Las características de las cinco subcuencas se presentan en la Tabla 2, en la que se han incluido el número de registros de peces y las localidades correspondientes, obtenidos de las bases de datos consultadas.
Se consultaron los registros de las bases de datos de la Colección Nacional de Peces Dulceacuícolas Mexicanos de la Escuela Nacional de Ciencias Biológicas del ENCB–IPN (Díaz–Pardo, 2003a, 2003b); de la Colección Nacional de Peces del Instituto de Biología de la IBUNAM (Espinosa–Pérez, 2001) y de la Colección Ictiológica de la Universidad Autónoma de Nuevo León (Lozano–Vilano, 2002). Los datos de las colecciones señaladas, así como de instituciones de otros países, se integraron, bajo criterios de calidad, al Sistema Nacional de Información sobre Biodiversidad (SNIB) de la Comisión para el Conocimiento y Uso de la Biodiversidad (CONABIO, 2008). También se analizaron los registros de la Universidad Autónoma de Querétaro (Díaz–Pardo, 2009), y la información proporcionada por las Universidades de Guadalajara y Wisconsin para esta cuenca.
También se revisaron los trabajos sobre la comunidad íctica en la región de Guzmán–Arroyo y Lyons (2003), Mercado–Silva y Lyons (2004), Miller et al. (2005); Además de los datos contenidos en los estudios ambientales llevados a cabo en las presas Agua–milpa, El Cajón y La Yesca por la Comisión Federal de Electricidad; los resultados del proyecto Arcediano realizado por la Comisión Estatal de Agua de Jalisco y de la presa Zapotillo, en el río Verde, hecho por la Comisión Nacional del Agua. Cuando existía sólo el dato de presencia, se cotejó con colectores y registros disponibles la presencia de las especies en las distintas subcuencas.
Los registros consultados correspondieron a datos de especie, sus datos taxonómicos, de la colección a la que pertenece, del colector, de la localidad, del cuerpo de agua, la fecha, el número de ejemplares y las coordenadas geográficas. Dado que en los registros no se incluyen los métodos de recolecta dicha información no formó parte del análisis de este trabajo.
La ubicación de las localidades de registro conforme a las etapas de desarrollo hidráulico (1ª hasta 1970, 2ª 1971–2000 y 3ª2001–2006) aparece en la Figura 4, donde se observa que algunas regiones, como los ríos Bolaños, Huaynamota y la parte media del Santiago, tienen escasos registros. La primera etapa correspondió a estudios de inventario y descriptivos de especies por lo que las localidades se ubican en casi toda la cuenca, para la segunda y tercera etapa las recolectas se realizaron principalmente para estudios de presas y aprovechamientos en tramos específicos.
Para integrar el listado sistemático de peces se adoptaron los criterios de Miller et al. (2005), excepto para el nombre genérico Menidia, ya que de acuerdo con lo propuesto por diversos autores (Barbour, 1973a, 1973b; Barbour & Chernoff, 1984; Paulo–Maya, 2000) y dado que no existe un consenso generalizado para su aceptación por los ictiólogos mexicanos (Miller et al., 2009), se siguió usando el nombre de Chirostoma.
Para el análisis de la distribución espacial y temporal de las especies, los registros se agruparon por subcuencas y se relacionaron con el desarrollo hidráulico, considerando que tres etapas de desarrollo, la primera: de 1951 a 1970, que incluyó la puesta en marcha de la presa Santa Rosa en 1964, que limitó la conectividad aguas abajo en el río Santiago y con los tributarios Verde y Juchipila; la segunda etapa, de 1971 a 2000 que incluyó la construcción y operación de las presas Aguamilpa y San Rafael en 1994, que limitaron la conectividad en la parte baja del río Santiago con el río Huaynamota; y la tercera etapa de 2001 a 2006, cuando se incorporó al sistema de embalses en cascada la presa El Cajón, entre Santa Rosa y Aguamilpa, y además se inició, aguas arriba de la presa El Cajón, la construcción del proyecto La Yesca, la cual llenará su embalse en el año 2012 y aislará al río Bolaños.
Para el análisis de la presencia de taxa de peces en las distintas etapas del desarrollo hidráulico, se consideraron los límites temporales de cada una de éstas. Conforme al origen y distribución de las especies, se obtuvo la proporción de especies nativas de México, endémicas de la región Lerma–Chapala–Santiago, introducidas accidentalmente y para las pesquerías, así como las traslocadas de otras áreas de distribución natural (Miller et al., 2005) por etapas de desarrollo hidráulico.
Para el análisis espacial se señaló la presencia de los taxa en las distintas cuencas y los límites de su gradiente altitudinal de distribución obtenido de las bases de datos. Este gradiente se revisó conforme la distribución señalada para cada especie por Guzmán–Arroyo y Lyons (2003), Mercado–Silva y Lyons (2004), Miller et al. (2005). Considerando la ausencia temporal y la reducción del intervalo de distribución altitudinal, se señalaron las especies posiblemente extinguidas y bajo riesgo incluidas en la NOM–059–SEMARNAT (2010), así como en la lista de especies en riesgo reportadas por la Sociedad Americana de Pesquerías (Jelks et al., 2008).
Para el análisis de la abundancia se tomó el número de ejemplares capturados de cada registro por localidad y se comparó con el total de organismos de todo el período (1951–2006) y por etapa de desarrollo hidráulico. Es importante señalar que la segunda etapa reunió la mayor abundancia de peces, no por su mayor duración en el tiempo, sino seguramente por las intensas recolectas realizadas por el Instituto de Biología de la UNAM, en los estudios de la presa Aguamilpa. La amplitud de distribución de cada una de las especies, se calculó con base en su presencia por localidades, y en relación a la proporción del total de las mismas (Díaz–Pardo et al., 1993), considerando tanto el periodo completo (1951–2006), como sus 3 etapas de desarrollo hidráulico.
Los mapas de la cuenca, ubicación de embalses y localidades de registro, se elaboraron con el sistema de información geográfica Arc–View 3.2, tomando como base la información cartográfica digitalizada del Catálogo de Metadatos Geográficos de la CONABIO publicada el 18 de marzo de 2001 http://www.conabio.gob.mx/informacion/gis/.
RESULTADOS
Se obtuvieron 1032 registros, procedentes de 132 localidades, abarcando el período de 1951 a 2006. De éstos, 995 exhibieron datos completos de los ejemplares capturados, mientras que 87 se refieren sólo a la presencia de los taxa en los ríos. El río Santiago presentó el mayor número de registros (676) y localidades (53), mientras que el río Verde, aunque concentró un importante número de localidades (49), reunió un número de registros significativamente menor (165). Las subcuencas Juchipila, Bolaños y Huaynamota, presentaron el menor número de registros y localidades (Tabla 2).
El listado sistemático histórico de peces obtenido para la cuenca del río Santiago comprendió 13 familias, 30 géneros y 44 especies, 38 se registraron en el río Santiago, 25 en el Verde, 14 en el Juchipila, 10 en el Bolaños y 12 en el Huaynamota. La Tabla 3 incluye la información de las subcuencas donde se ha registrado cada una de las especies así como sus acrónimos e información de su origen y distribución, considerando endémicas de la región Lerma–Chapala–Santiago (n = 13); nativas de México (n = 22), introducidas (n = 7) y traslocadas (n = 2) (Espinosa–Pérez et al., 1993; Miller et al., 2005).
Las familias más diversas fueron: Atherinopsidae (2 géneros y 8 especies), Cyprinidae (6 géneros y 8 especies) y Goodeidae (6 géneros y 8 especies), quienes también tuvieron el mayor porcentaje de especies endémicas de la región Lerma–Chapala–Santiago.
Utilizando los acrónimos de las especies, la Tabla 4 muestra que 17 están presentes durante las tres etapas de desarrollo hidráulico de la cuenca, 5 en las dos primeras etapas y 5 en las dos últimas; 2 en la primera y última etapas; 9 sólo en la primera etapa; 5 solamente en la segunda y una introducida sólo en la tercera etapa.
Un grupo importante fue el conformado por las 9 especies sólo registradas en la primera etapa (Tabla 3), de las cuales 8 son endémicas y 4 pertenecientes al género Chirostoma (C. consocium, C. labarcae, C. lucius, C. sphyraena); Algansea popoche (ciprínido) y tres godeidos: Allotoca dugesii, Alloophorus robustus, Skiffia lermae, así como una nativa (Zoogoneticus quitzeoensis), cuya distribución se limita a la parte alta de la cuenca.
Las 5 especies presentes solo en la segunda etapa del desarrollo hidráulico comprendieron peces marinos que remontan corrientes (Lile gracilis, Lile stolifera, Gobiomorus maculatus y Awaous transandeanus), así como al pez migratorio Agonostomus montícola, que ocupan las porciones baja e intermedia del área de estudio. El pez introducido Oreochromis aureus constituyó la única especie con registro a partir de la tercera etapa de desarrollo hidráulico de la cuenca, proveniente de la parte media–alta de la cuenca.
La distribución de los taxa por su origen y presencia en las subcuencas y distintas etapas de desarrollo hidráulico se muestran en la Tabla 5, en la que se observa que entre la primera etapa de 1951 a 1970 y la tercera de 2001 a 2006, hubo una disminución de las especies nativas y endémicas, al mismo tiempo que se incrementaron las especies introducidas, principalmente en los ríos Santiago y Verde, que cuentan con un mayor número de registros.

Las especies endémicas, nativas, e introducidas con registros en las tres etapas de desarrollo hidráulico fueron las siguientes: 4 endémicas: Chirostoma arge, Algansea monticola, Yuriria alta y Chapalichthys encaustus; 10 nativas: Chirostoma jordani. C. beani, Dorosoma smithi, Algansea tincella, Gobie–sox fluviatilis, Goodea atripinnis, Xenotoca variata, Ictalurus dugesii, Poecilia butleri y Poeciliopsis infans, y 3 introducidas: Micropterus salmoides, Oreonchromis mossambicus y Cyprinus carpio.
Conforme a su distribución altitudinal las especies se ubicaron como se muestra en la Tabla 6, donde se observa que 12 especies se distribuyen a la parte alta de la cuenca con altitudes mayores de 1500 msnm, cinco de la familia Atherinopsidae (Chirostoma consocium, C. labarcae, C. lucius, C. sphyraena y C. Chapalae); tres de Cyrpinidae: Algansea popoche, Aztecula sallaei e Hybopsis calientis y 4 godeidos: Alloophorus robustus, Chapalichthys encaustus, Xenotoca variata y Skiffia lermae.

Cuatro especies están restringidas en la parte baja de la cuenca (con altitud hasta los 1200 msnm) (Chirostoma arge, Allotoca dugesii, Xenotoca melanosoma y Zoogoneticus quitzeoensis). En los 900 msnm se tienen registros de A. monticola y Gobiesox fluviatilis, mientras que a los 600 msnm están registrados Chirostoma jordani, Oreochromis aureus, O. niloticus, Cyprinus carpio y Carassius auratus. Entre los 300 y los 1500 msnm, se registraron a las especies Algansea tincella, Yuriria alta, Goodea atripinnis e Ictalurus dugesii. En la parte baja de la cuenca las especies distribuidas a altitudes menores de 300 msnm fueron Dorosoma smithi, Lile gracilis, L. stolifera. Agonostomus montícola y Poeciliopsis latidens.
Sólo las siguientes 8 especies se distribuyeron a lo largo de todo el intervalo altitudinal: Atherinella crystallina, Scartomyzon austrinus, Lepomis macrochirus, Micropterus salmoides, Cichlasoma beani, Oreochromis mossambicus, Poecilia butleri y Poeciliopsis infans.
En cuanto a la abundancia relativa, el número total de individuos registrados fue de 27,525; y sólo 6 de los 44 taxa rebasaron el 5% de la abundancia, sumando entre ellos 71.5% del total de individuos registrados. Entre estos seis está incluida la especie introducida Lepomis macrochirus (Fig. 5). Dos de tales taxa Poeciliopsis latidens y Atherinella crystallina ya no se registraron en la tercera etapa de desarrollo hidráulico, ambas especies nativas de México (Fig. 5).
La abundancia de las especies de peces, con relación a las etapas del desarrollo hidráulico de la cuenca indicó que en la segunda etapa se registró el mayor número de ejemplares (19,383), seguido de la primera (con 6144 ejemplares) y la tercera (con 1998 ejemplares). Los taxa más abundantes en la primera etapa fueron los siguientes en orden descendente: Goodea atripinnis, Poeciliopsis infans, Xenotoca variata, P. latidens, Chirostoma jordani y Algansea tincella. En la segunda, P. latidens, Poecilia butleri, Lepomis macrochirus, Atherinella crystallina, Cichlasoma beani y Oreochromis mossambicus, mientras que en la tercera etapa predominaron: Algansea monticola, O. mossambicus, P. infans, Micropterus salmoides, O. aureus y Scartomyzon austrinus. Es importante mencionar que la única especie que mantuvo abundancias importantes, a lo largo de las tres etapas de desarrollo hidráulico, fue Poecilia butleri(POBU) y Poeciliopsis infans (POIN) (Tabla 7).
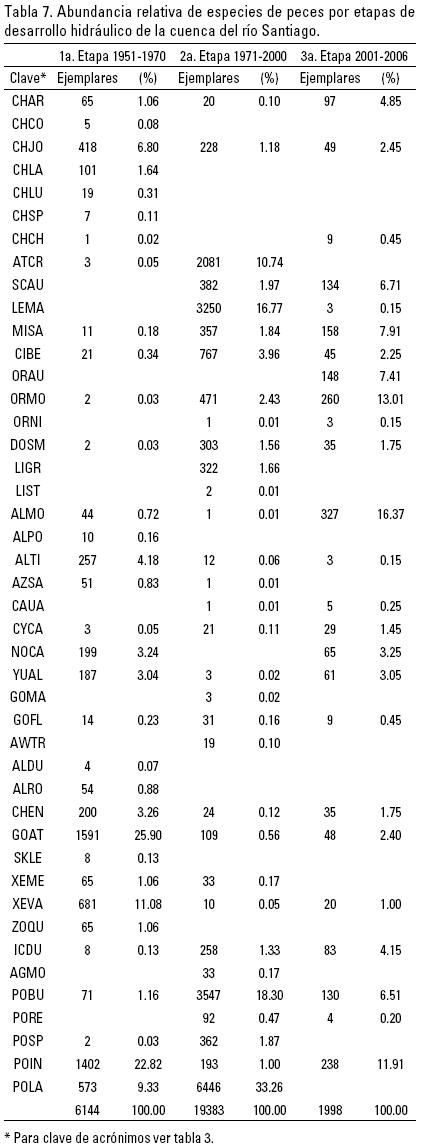
Por el contrario los siguientes taxa han disminuido su abundancia como es el caso de Goodea atripinnis que estuvo representado en la primera etapa por 1591 especímenes y en la tercera por tan solo 48 (reducción de 23.7 a 2.3% de abundancia relativa). En el mismo orden de cifras, Xenotoca variata tuvo una disminución de especímenes de 681 a 20 y 10.1 a 1.0% en abundancia relativa; Chirostoma jordani de 418 a 49 y pasando de 6.2 a 2.4%; Algansea tincella de 257 a 3 y pasando de 3.8 a 0.1%; Chapalichthys encaustus de 200 a 35 y pasando de 3 a 1.7%. Un caso especial fue Hybopsis calientis que redujo el número de ejemplares (de 199 a 65), pero incrementó su abundancia relativa de 2.9 a 3.2 debido al efecto conjunto de la composición y abundancia de los componentes de la comunidad en la 3a. etapa (Tabla 7).
En cuanto al análisis de la amplitud temporal de las especies (histórica), la mayor correspondió a Goodea atripinnis quien ha ocupado 47 localidades (35.6%) del total de 132 en el transcurso del desarrollo hidráulico del río, seguida de Poecilliopsis infans (40 y 30.3%) y Algansea tincella (23 y 17.4%) (Fig. 6). De éste grupo de taxa, Scartomyzon austrinus sólo se ha registrado en las dos últimas etapas de desarrollo hidráulico (Fig. 6).
Del análisis de la Tabla 8 se deduce que aun cuando el número de localidades muestreadas fue diferente entre las tres etapas del desarrollo hidráulico del río Santiago (68, 34 y 30 respectivamente), el número de especies registradas por etapa tuvo poca variación, resultando en 34, 32 y 25 respectivamente.
DISCUSIÓN
La riqueza específica histórica de 44 taxa de peces para la cuenca del río Santiago coincide con lo reportado por Espinosa–Pérez etal. (1993). Las diferencias en el número de taxa de las Familias Atherinopsidae, Cyprinidae y Goodeidae, citadas por Guzmán–Arroyo y Lyons (2003) y por Jelks et al. (2008) se deben tanto a inclusiones de taxa regionales del sistema Lerma–Chapala–Santiago como a cambios taxonómicos del sistema de clasificación.
Por otra parte, la presencia de un 30% (13 de 44) de especies endémicas en los registros confirma la importancia de la radiación adaptativa de especies en este sistema Lerma–Chapala–Santiago, donde se ha señalado hasta un total del 66% de especies endémicas en otros trabajos (Díaz–Pardo et al., 1993; Lyons et al., 1998; Soto–Galera et al., 1998; Miller et al., 2005; Guzmán–Arroyo & Lyons, 2003; Mercado–Silva et al., 2006; Domínguez–Domínguez et al., 2008; Jelks et al., 2008; Contreras–Balderas et al., 2008). En la cuenca se reconocieron 9 especies introducidas, de las cuales 7 se deben considerar invasoras, ya que se han expandido en un gran número de cuencas del país (Contreras–Balderas et al., 2009; March & Martínez, 2008), y dos del género Poecilia (P. reticulata y P. sphenops) (Gesundheit & Macías–García, 2007) son traslocadas.
Atherinopsidae, Cyprinidae y Goodeidae, fueron las tres familias más representativas en la cuenca del río Santiago, tanto por su riqueza específica como por el número de especies endémicas. Sin embargo, 12 de ellas (5 en el caso de los aterinópsidos y goodeidos, y 2 para los ciprínidos) no tienen registros en los últimos 10 años. Cabe señalar que la riqueza que le han conferido estas familias a la cuenca se considera actualmente empobrecida por el deterioro ambiental (Gesundheit & Macías–García, 2007; Abell et al., 2008; Domínguez–Domínguez et al., 2008).
La mayor concentración de registros y localidades en los ríos Santiago (su parte cercana al lago de Chapala) y Verde (parte alta), se asocia a la accesibilidad que tuvieron durante su desarrollo regional diversos ictiólogos, quienes reconocieron la importancia de esta como una zona de alta diversidad y especiación (Miller et al., 2005), así como a la presión de uso que ha habido sobre la cuenca y sus recursos hídricos.
La distribución natural de las especies de peces en el río Santiago señala con claridad las características faunísticas de la parte inicial de este río, ya que históricamente estuvo habitada por especies ahora restringidas al lago de Chapala, como son los componentes del género Chirostoma: C. chapalae, C. consocium, C. labarcae, C. lucius y C. sphyraena, además de los goodeidos Chapalichthys encaustus y Zoogoneticus quitzeoensis y el ciprínido Algansea popoche. Tamayo (1996) señala que la salida del lago de Chapala es estrecha y con baja pendiente en los primeros kilómetros, por lo que el río Santiago fluye a baja velocidad en su primer tramo, dándole características lénticas al mismo, que es el hábitat sobre el cual muestran preferencia dichos peces. Posteriormente, tanto El Salto de Juanacatlán que se ha señalado como una barrera física de distribución (Miller et al., 2005), como las presas El Salto y Poncitlán, que se construyeron en 1895 y 1897 respectivamente, para derivar agua del río Santiago, junto con la degradación ambiental que se hizo referencia, determinaron que, con la excepción de Chirostoma chapalae y Chapalichthys encaustus, el resto de las especies sólo se registraron en la primera etapa de desarrollo hidráulico de la cuenca.
En el otro extremo del área de estudio se han registrado taxa correspondientes a dos componentes biogeográficos, las especies marinas eurihalinas (Castro–Aguirre et al., 1999) que son tolerantes a la disminución de la salinidad, como son Llile gracilis, L. stolifera, Dorosoma smithi, Gobiomorus maculatus y Awaous transandeanus, y las llamadas especies secundarias (Myers, 1940), cuyo origen es dulceacuícola, pero que son tolerantes al aumento de la salinidad, representadas por Poecilia butleri, P. la–tidens y Cichlasoma beani. En conjunto ambos componentes son representativos de la parte baja de la cuenca del río Santiago, misma que se vio modificada en 1994 (segunda etapa del desarrollo hidráulico) con la construcción de las presas Aguamilpa y San Rafael, porque se asume que las cortinas y embalses de dichas presas constituyeron un límite para la distribución de estas especies aguas arriba.
Recientemente, también se ha señalado que el pez catádromo Agonostomus monticola y el anádromo Awaous transandeanus, han resultado afectados por la cortina del embalse Aguamilpa (Guzmán–Arroyo et al., 2009). Ambas especies se registraron en los ríos Huaynamota y Santiago entre 1991 y 1992, pero desde 1997 sólo han sido capturados en sitios aguas abajo de la presa San Rafael, lo que podría significar que distintas poblaciones quedaron aisladas. Se sabe que en otros sistemas, A. monticola establece poblaciones entre los embalses y es capaz de completar su ciclo reproductivo en la parte alta de las cuencas (Díaz–Pardo, 2008), aunque por presentar una densidad baja, permanece el riesgo potencial de su extinción local debido a la disminución de su diversidad genética (Anderson et al., 2006; Rojas & Rodríguez, 2008).
De las especies nativas que cuentan con registros en las tres etapas de desarrollo hidráulico, como Chirostoma jordani, Cichlasoma beani, Dorosoma smithi, Algansea tincella, Gobiesox fluviatilis, Goodea atripinnis, Xenotoca variata, Ictalurus dugesii, Poecilia butleri y Poeciliopsis infans, se estima que, a pesar de la fragmentación del hábitat, pueden existir poblaciones aisladas de estas especcies en la entrada de los embalses del río Santiago y en sus tributarios. De la misma manera, otras especies endémicas que pueden presentar poblaciones aisladas son: C. arge, Y. alta y C. encaustus.
Las especies introducidas para el desarrollo de pesquerías y actividades deportivas en los embalses como Oreochromis mossambicus, Cyprinus carpio y Micropterus salmoides, se han establecido y mantenido en la cuenca desde la primera etapa. Por otra parte, Lepomis macrochirus, O. niloticus, Carassius auratus y Poecilia reticulata (reconocida como traslocada), aparecen a partir de la segunda etapa de desarrollo y O. aureus cuando inicia la tercera etapa. En el caso de L. macrochirus, que se introdujo como forrajero de M. salmoides, aunque ha sido capturada en un número limitado de localidades, es abundante debido a su gran capacidad de adaptación, que la coloca como una especie posiblemente invasora (Guzmán–Arroyo & Lyons, 2003; Olden & Poff, 2005; Contreras–Balderas et al., 2008). En cambio P. sphenops, también traslocada, y con intervalo altitudinal amplio (<100 a 900 msnm), ya no fue registrada en la tercera etapa de desarrollo hidráulico de la cuenca. Seguramente las tilapias (Orechromis spp.) deberían haber ocupado un lugar privilegiado en cuanto a su abundancia, pero esto no se vio reflejado en los resultados del presente trabajo, porque los registros de los cuales derivó el análisis, fueron de tipo biológico y no pesquero.
Con base en lo antes mencionado, se considera que la localización de las presas, su magnitud y los períodos de construcción, jugaron, en su momento y juegan hasta ahora, un papel importante en la movilidad y dispersión de las especies de peces, aunado a otras presiones ecológicas como han sido el deterioro y reducción del hábitat y la introducción de especies, entre las más importantes (Lyons et al., 1998; Díaz–Pardo, 2003b; Gutiérrez–Hernández, 2003; Mercado–Silva & Lyons, 2004; Miller et al., 2005; Jelks et al., 2008; Domínguez–Domínguez et al., 2008; Guzmán–Arroyo & Lyons, 2003 y Guzmán–Arroyo et al., 2009).
El conocimiento y la intensidad de muestreo en ciertas sub–cuencas como la del Río Verde por diferentes investigadores (entre ellos, Lyons, Mercado–Silva y Domínguez–Domínguez), puede ser una de las razones de los registros únicos de cinco especies (Aztecula sallaei, Allotoca dugesii, Skiffia lermae, Xenotoca melanosoma y X. variata, la última registrada recientemente). Por otro lado, como estos taxa son característicos de la subprovincia biogeográfica del Medio Lerma (Díaz–Pardo et al., 1993) su presencia en el río Verde puede ser indicativa de conexiones hidrológicas prehistóricas entre ambas áreas y no necesariamente por su dispersión a través del sistema Lerma–Chapala–Santiago. En relación a estas especies, es importante mencionar que debido a las condiciones de fragmentación y deterioro del río Verde, sus poblaciones pueden haber sido afectadas, con la consecuente reducción de la capacidad reproductiva de las poblaciones pequeñas (Domínguez–Domínguez et al., 2008). Por otro lado, Alloophorus robustus, Skiffia lermae, Hybopsis calientis, Ictalurus dugesii y Scartomyzon austrinus (que también habitan en el Medio Lerma) han sido mencionadas en riesgo de extinción en las cuencas de la Mesa Central (Lyons et al., 1998; Soto–Galera et al., 1998; Mercado–Silva et al., 2006).
De las 9 especies que sólo se registraron durante la primera etapa del desarrollo hidráulico (Tabla 4), puede intuirse que las 8 endémicas, aparentemente han sido extintas de la cuenca del río Santiago (Chriostoma consocium, C. labarcae, C. lucius, C. sphyraena, Algansea popoche, Allotoca dugessii, Alloophorus robustus y Skiffia lermae). Se detectaron además, otros taxa nativos que mostraron una clara disminución en su amplitud de distribución, como es el caso de Goodea atripinnis (que de estar presente 35 localidades en la primera etapa, pasó a 7 en la tercera), Algansea tincella (de 20 a 2), Hybopsis calientis (de 13 a 2), C. jordani(de 13 a 3), Yuriria alta (de 13 a 6), y Poecilliopsis infans (de 19 a 12 localidades) (Tabla 8).
La condición inversa también fue detectada, es decir especies que han presentado un incremento en el número de localidades que ocupan a lo largo del tiempo como es el caso de Oreochromis aureus (que aumento de 0 en la primera etapa a 10 localidades en la tercera) al igual que Micropterus salmoides (que paso de 1 a 6), O. mossambicus (que paso de 1 a 5) y Algansea monticola (que paso de 5 a 9 sitios). Los tres primeros taxa corresponden a peces introducidos, que seguramente por sus preferencias a los ambientes lénticos, se han visto favorecidas por la construcción de presas (Tabla 8).
El número de especies en riesgo para la cuenca del río Santiago, se ha incrementado de 9 a 10 especies, en las versiones de la Norma 059 en el periodo, de 2001 a 2010 NOM–059–SEMARNAT(2010), lo que representa el 22.7% de la riqueza obtenida en este trabajo. Las especies enlistadas en la categoría de "amenazadas" son: Chirostoma labarcae, Algansea popoche, Skiffia lermae y Zoogoneticus quitzeoensis, a las que se debe agregar Allotoca dugesii, que pasó a la categoría de "en peligro de extinción". Esas cinco especies sólo tienen registros en la primera etapa de desarrollo hidráulico. También forman parte de la lista: Gobiesox fluviatilis, Ictalurus dugesii (amenazadas) y P. butleri (bajo protección especial), que están presentes en todas las etapas, así como Poecilia latidens (amenazada) y Xenotoca melanosoma (en peligro de extinción), únicamente registradas a partir de la segunda etapa.
La AFS (American Fisheries Society) incluye en su listado a todas las especies antes mencionadas, excepto a P. butleri y agregan con la categoría "en peligro" a Chirostoma lucius y Chirostoma sphyraena y como "vulnerables" a Scartomyzon austrinus, Algansea tincella, Aztecula sallaei, Hybopsis calientis y Chapalichthys encaustus (Jelks et al., 2008). La conjunción de los dos listados denota con claridad que la situación de riesgo es mayor en las familias características del sistema Lerma–Chapala–Santiago, es decir Atherinopsidae, Cyprinidae y Goodeidae.
De los 17 taxa que aparecen en los listados mencionados, cuatro son sensibles (Chirostoma labarcae, Skiffia lermae, Zoogonecticus quitzeoensis e Ictalurus dugesii) y otras seis son medianamente tolerantes (C. arge, Scartomyzon austrinus, Algansea tincella, Hybopsis calientis, Allotoca dugesii y Chapalichthys en–caustus (Díaz–Pardo et al., 1993).
A pesar de las diferencias existentes entre los métodos de recolecta, las localidades incluidas en cada etapa y la concentración de las recolectas sólo en ciertos tramos de la cuenca debido a intereses particulares de estudio , los resultados de abundancia y amplitud de distribución, indican que en términos globales, la presencia de especies nativas y endémicas ha disminuido, y en cambio se ha incrementado el número de especies introducidas; lo anterior es más evidente en las subcuencas Santiago y Verde, que son las que cuentan con más registros.
Hoy, aproximadamente el 50 % de la longitud del río Santiago se encuentra embalsada, incluyendo la presa La Yesca, que actualmente está en construcción y por llenar su embalse en este año (2012),, y lo preocupante es que aún restan áreas potenciales de aprovechamiento público o generación de energía, tanto en la corriente principal, como en los tributarios.
La evaluación de los cambios regionales en la comunidad íctica, asociados al desarrollo de grandes obras de infraestructura hidráulica, no está realizándose actualmente por ninguna institución, ya que los únicos estudios requeridos para autorizar este tipo de obras, se solicitan a través de las manifestaciones de impacto ambiental, que en general incluyen el inventario de especies y el señalamiento de las especies incluidas en la NOM–059–SEMARNAT–2010. El seguimiento de los cambios esperados en la estructura y función de la comunidad por la infraestructura o usos del agua es además insuficiente, porque no se aplica un programa de monitoreo, ni en la construcción, ni en la operación de las obras. Estudios aislados como los de Gutiérrez–Hernández (2003) y Mercado–Silva et al. (2009), han señalado efectos de los embalses y de las especies introducidas sobre la estructura de la red trófica, siendo uno de éstos el traslape trófico aguas arriba y abajo de las presas. Los efectos conjuntos de estas acciones han llevado a la extinción de especies en tramos del río Lerma y así como a extinciones locales (Díaz–Pardo et al., 1993; Mercado–Silva et al., 2006, 2009; De la Vega–Salazar, 2006; Domínguez–Domínguez et al., 2008).
Estudios recientes en la región central de México están explorando la manera en que las especies nativas han sido impactadas por las actividades humanas, para determinar los puntos rojos o focales de diversidad remanente y así guiar las prioridades de conservación (Helmus et al., 2007), por lo que resulta imperativo revisar las variaciones impuestas por los embalses, antes de avanzar, con escasos estudios, en la determinación y autorización del futuro potencial para abastecimiento o generación de energía de las subcuencas menos explotadas.
De esta forma, las cuencas de los ríos Bolaños y Huaynamota, por sus condiciones de menor deterioro y fragmentación, dado que albergan especies endémicas, podrían proponerse como regiones representativas y prioritarias, para la conservación de la ictiofauna dulceacuícola de la cuenca del río Santiago.
AGRADECIMIENTOS
Al personal de la Coordinación de Información y Servicios Externos de la CONABIO por el aporte de datos para la realización de este trabajo. A Norman Mercado Silva y John Lyons. Fernando Álvarez Noguera y Héctor Espinoza Pérez, del Instituto de Biología de la UNAM. A Eduardo Soto Galera de la Escuela Nacional de Ciencias Biológicas del IPN, así como a Agustín Camacho Rodríguez de la Universidad de Guadalajara, por proporcionarnos sus registros. Al personal de la Comisión Federal de Electricidad, por la información sobre las presas. También agradecemos a Marco Antonio Mijangos Carro y a Sergio Rodríguez Torres del Instituto Mexicano de Tecnología del Agua, por su apoyo en el manejo del sistema de información geográfica.
REFERENCIAS
Abell, R., M. L.Thieme, C. Revenga, M. Bryer, M. Kottelat, N. Bogutskaya, B. Coad, N. Mandrak, S. Contreras Balderas, W. Bussing, M. L. J. Stiassny, P. Skelton, G. R. Allen, P. Unmack, A. Naseka, R. Ng, N. Sindorf, J. Robertson, E. Armijo, J. V. Higgins, T. J. Heibel, E. Wikramanayake, D. Olson, H. L. López, R. E. Reis, J. G. Lundberg, M.H. Sabaj Pérez & P. Petry. 2008. Freshwater Ecoregions of the World: a new map of biogeographic units for freshwater biodiversity conservation. BioScience 58 (5): 403–414. [ Links ]
Anderson, E. P., M. C. Freeman & C. M. Pringle. 2006. Ecological consequences of hydropower development in Central America: Impacts of small dams and water diversion on neotropical stream fish assemblages. River Research and Applications 22 (4): 397– 411. [ Links ]
Barbour, C. D. 1973a. The systematics and evolution of the genus Chirostoma, Swainson (Pisces, Atherinidae). Tulane Studies in Zoology and Botany 18 (3): 97–141. [ Links ]
Barbour, C. D. 1973b. A biogeographical history of Chirostoma (Pisces, Atherinidae): a species flock from the Mexican plateau. Copeia (3): 533–556. [ Links ]
Barbour, C. D. & B. Chernoff. 1984. Comparative morphology and morphometrics of the pescados blancos (genus Chirostoma) from Lake Chapala, México. In: Echelle A. A. & I. Kornfield (Eds.). Evolution of fish species flocks. University of Maine Pres. U.S.A. 111–127. [ Links ]
Birgitta, M. R., R. Jansson & C. Nilsson. 2010. Effects of hydropower generation and opportunities for environmental flow management in Swedish riverine. Freshwater Biology 55 (1): 49–67. [ Links ]
Castro–Aguirre, J. L., H. S. Espinosa–Pérez & J. J. Schmtiter–Soto. 1999. Ictiofauna Estuarino–Lagunar y Vicaria de México. Instituto Politécnico Nacional y Editorial Limusa S.A. de C. V. Cd. de México. 711 p. [ Links ]
Chiapa H. L. & H. Marengo M. 2008. Criterios de diseño de presas de enrocamiento con cara de concreto. Memorias del XX Congreso Nacional de Hidráulica. Toluca, México. 164 p. [ Links ]
Comisión Nacional del Agua–Instituto Mexicano de Tecnología del Agua (Conagua–IMTA). 2009. Estudio de actualización de la disponibilidad y balance hidráulico en aguas superficiales de la Región hidrológica No. 12 Río Santiago, utilizando el modelo ArcHydro. Convenio CNA–OCLSP–JAL–CONV–002–2008. Informe Final. 243 p. [ Links ]
Comisión Nacional del Agua (Conagua). 2010. Acuerdo por el que se actualiza la disponibilidad media anual de las aguas superficiales en las cuencas hidrológicas Río San Pedro, Presa Calles, Presa El Niágara, Presa El Cuarenta, Río de Lagos, Presa Ajojucar, Río Grande, Río Encarnación, Río Aguascalientes, Río San Miguel, Río del Valle, Río Verde 1, Río Verde 2, Río Palomas, Presa El Chique, Río Juchipila 1, Río Juchipila 2, Río Santiago 1, Río Santiago 2, Presa Santa Rosa, Río Santiago 3, Río Tepetongo, Río Tlaltenango, Arroyo Lobatos, Río Bolaños 1, Río Bolaños 2, Río San Juan, Río Atengo, Río Jesús María, Río Huaynamota, Río Santiago 4, Río Santiago 5 y Río Santiago 6, mismas que forman parte de la subregión hidrológica Río Santiago de la región hidrológica número 12 Lerma–Santiago, y su ubicación geográfica. Diario Oficial de la Federación. México, D. F. Noviembre 29: 1–128. [ Links ]
CONABIO. 2008. Capital natural de México, vol. I: Conocimiento actual de la biodiversidad. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad México. 621 p. [ Links ]
Contreras–Balderas, S. 1999. Annotated checklist of introduced invasive fishes in Mexico, with examples of some recent introductions. In: Claudi R. & J. Leach (Eds). Non–indigenous freshwater organisms: vectors, biology and impacts. Lewis Publishers. Washington, D.C., pp. 33–54. [ Links ]
Contreras–Balderas, S., G. Ruiz–Campos, J.J. Schmitter–Soto, E. Díaz–Pardo, T. Contreras–McBeath, M. Medina–Soto, L. Zambrano–Gonzalez, A.Varela–Romero, R. Mendoza–Alfaro, C. Ramirez–Martinez, M. A.Leija–Tristan, P. Almada–Villela, D. Hendrickson & J. Lyons. 2008. Freshwater fishes and water status in Mexico: A country–wide appraisal. Aquatic Ecosystem Health and Management 11 (3): 246–256. [ Links ]
De La Vega–Salazar, M. Y., E. Ávila–Luna, & C. Macías–García. 2003. Ecological evaluation of local extinction: the case of two genera of endemic Mexican fish, Zoogoneticus and Skiffia. Biodiversity and Conservation 12 (10): 2043–2056. [ Links ]
De La Vega–Salazar, M. Y. 2006. Estado de conservación de los peces de la familia Goodeidae (Cyprinodontiformes) en la Mesa Central de México. Revista de Biología Tropical54 (1): 163–177. [ Links ]
Díaz–Pardo, E. 2003a. Sistematización de la Colección Nacional de Peces Dulceacuícolas Mexicanos de la Escuela Nacional de Ciencias Biológicas, IPN. Instituto Politécnico Nacional. Bases de datos SNIB–CONABIO Proyecto No. T027. México. [ Links ]
Díaz–Pardo, E. 2003b. Peces en riesgo de la Mesa Central de México. Instituto Politécnico Nacional. Escuela Nacional de Ciencias Biológicas. Bases de datos SNIB–CONABIO Proyecto No. W039. México. [ Links ]
Díaz– Pardo, E. 2008. Caracterización del subsistema biótico en el Sistema de energía renovable Moctezuma. Informe Técnico Final para la Comisión Federal de Electricidad, México. 104 p. [ Links ]
Díaz–Pardo, E. 2009. Sistematización de la Colección de Peces del Centro de México. Universidad Autónoma de Querétaro, Facultad de Ciencias Naturales. Bases de datos SNIB–CONABIO Proyecto No. EC–007, México. [ Links ]
Díaz–Pardo, E., M. A. Godínez Rodríguez, E. López–López & E. Soto–Galera. 1993. Ecología los peces de la cuenca del río Lerma, México. ENCB–IPN México. Anales de la Escuela Nacional de Ciencias Biológicas 39: 103–127. [ Links ]
Domínguez–Domínguez, O., L. Zambrano, L. H. Escalera–Vázquez, R. Pérez–Rodríguez & G. Pérez–Ponce de León. 2008. Cambios en la distribución de Goodeidos (Osteichthyes: Cyprinidontiformes: Goodeidae) en cuencas hidrológicas del centro de México. Revista Mexicana de Biodiversidad 79: 501–512. [ Links ]
Drástik, V., J. Kubecka, M. Tuser, M. Cech, J. Frouzova, O. Jarolim & M. Prechalova. 2008. The effect of hydropower on fish stocks: comparison between cascade and non–cascade reservoirs. Hydrobiologia 609 (1): 25–36. [ Links ]
Espinosa–Pérez, H. 2001. Computarización de la Colección Nacional de Peces del Instituto de Biología UNAM. Proyecto SNIB–CONABIO Proyecto No.T023. México. [ Links ]
Falke, J.A. & K. B. Gido. 2006. Spatial effects of reservoirs on fish assemblages in great plains streams in Kansas, USA. River Research and Applications 22: (1) 55–68. [ Links ]
Espinosa–Pérez, H., M. T. Gaspar–Dillanes & P. Fuentes–Mata. 1993. Listados Faunísticos de México III. Los peces dulceacuícolas Mexicanos. Instituto de Biología, Universidad Nacional Autónoma de México. México. 99 p. [ Links ]
Food and Agriculture Organization (FAO). 2001. Dams, fish and fisheries opportunities, challenges and conflict resolution. In: Marmulla, G. (Ed.). Fisheries technical paper 419. Rome, 166 p. [ Links ]
Garrido–Pérez A., M. L. Cuevas, H. Cotler, D.I. González & R. Tharme. 2010. Evaluación del grado de alteración ecohidrológica de los ríos y corrientes superficiales de México. Investigación Ambiental 2 (1): 25–46. [ Links ]
Gesundheit, P. & C. Macías–García. 2007. Distribución de peces dulceacuícolas introducidos en las cuencas del México central, con especial atención a las localidades dentro del área de distribución de los peces endémicos de la familia Goodeidae. Universidad Nacional Autónoma de México Instituto de Ecología. Informe final SNIB CONABIO Proyecto No. CE014. México D. F. [ Links ]
Gutiérrez–Hernández, A. 2003. Análisis limnológico e ictiofaunístico del embalse Zimapán Querétaro–Hidalgo. Tesis de Doctorado en Ciencias–Recursos bióticos. Facultad de Ciencias Naturales. Universidad Autónoma de Querétaro. México. 139 p. [ Links ]
Guzmán–Arroyo, M. & J. Lyons. 2003. Los peces de las aguas continentales del estado de Jalisco, México. Análisis preliminar. E.Gnosis (I) Art. 12. 1–37, disponible en línea en: http://redalyc.uaemex.mx/pdf/730/73000112.pdf (consultado el 10 septiembre 2010). [ Links ]
Guzmán–Arroyo, M., A. Orbe M., R. Maciel M. & M. López Hernández. 2009. El impacto de la "PH Aguamilpa" en las especies pesqueras, en la Cuenca Baja del Río Santiago, Nayarit. In: Peniche C. S. & M. Guzmán–Arroyo (Comps). Estudios de la Cuenca del Río Santiago. Un enfoque multidisciplinario. Universidad de Guadalajara, pp. 175–188. [ Links ]
Helmus, M. R., K. Savage, M. W. Diebel, J. T. Maxted & A. R. Ives. 2007. Separating the determinants of phylogenetic community structure. Ecology Letters 10: 917–925. [ Links ]
Hughes, R. M., J. N. Rinne & B. Calamusso. 2005. Introduction to historical changes in large river fish assemblages of the Americas. American Fisheries Symposium 45: 1 –12. [ Links ]
Jelks, H. L., S. J. Walsh, N. M. Burkhead, S. Contreras–Balderas, E. Díaz–Pardo, D. A. Hendrickson, J. Lyons, N. E. Mandrake, F. McCormick, J. S. Nelson, S. P. Platania, B. A. Porter, C. B. Renaud, J. J. Schmitter–Soto, E. B. Taylor, & M. L. Warren. 2008. Conservation status of imperiled North American freshwater and diadromous fishes. Fisheries 33 (8): 372–407. [ Links ]
March, M. & M. Martínez–Jiménez (Eds.). 2008. Especies invasoras de alto impacto a la biodiversidad. Prioridades en México. Instituto Mexicano de Tecnología del Agua y The Nature Conservancy. 74 pp. [ Links ]
López–López, E. & E. Díaz–Pardo. 1991. Cambios distribucionales de los peces del río de La Laja (Cuenca del río Lerma), por efecto de disturbios ecológicos. ENCB–IPN México. Anales de la Escuela Nacional de Ciencias Biológicas 35: 91–116. [ Links ]
Lozano–Vilano, M. L. 2002. Repatriación del material ictiológico de 12 estados de la República Mexicana, depositado en la colección ictiológica de la Universidad de Tulane. Universidad Autónoma de Nuevo León. Facultad de Ciencias Biológicas. Departamento de Zoología de Vertebrados. Proyecto SNIB–CONABIO S056. México. [ Links ]
Lyons, J., S. Navarro, P. A. Cochran, E. Santana–Castellón & M. Guzmán–Arroyo. 1995. Index of biotic integrity based on fish assemblages for the conservation of streams and rivers in West–Central Mexico. Conservation Biology 9 (3): 569–584. [ Links ]
Lyons, J., G. González–Hernández, E. Soto–Galera & M. Guzmán–Arroyo. 1998. Decline of freshwater fishes and fisheries in selected drainages of West–Central Mexico. Fisheries 23 (4): 10–18. [ Links ]
Lyons, J. & N. Mercado–Silva. 1999. Patrones taxonómicos y ecológicos entre comunidades de peces en ríos y arroyos en el oeste de Jalisco, México. Anales del Instituto de Biología UNAM 70 (2): 169–190. [ Links ]
McCarthy T. K., P. Frankiewicz, P. Cullen, M. Blaszkowski, W. O'Connor & D. Doherty. 2008. Long–term effects of hydropower installations and associated river regulation on River Shannon eel populations: mitigation and management. Hydrobiologia 609: 109–124. [ Links ]
Mercado–Silva, N. & J. Lyons. 2004. Listado de Peces Potenciales y Observados en las Cuencas del Río Verde y Río Santiago (Jalisco, México) y consideraciones sobre el impacto de modificaciones a su hábitat. Peces: Presa Arcediano. Preparado para: Fundación Manantlán para la Biodiversidad de Occidente, A.C. 16 p. [ Links ]
Mercado–Silva, N., J. Lyons, E. Díaz–Pardo, A. Gutiérrez–Hernández, C. P. Ornelas–García, C. Pedraza–Lara & M. J. V. Zanden. 2006. Long–term changes in the fish assemblage of the Laja River, Guanajuato, Central Mexico. Aquatic Conservation: Marine and Freshwater Ecosystems 16 (5): 533–546. [ Links ]
Mercado–Silva, N., M. R. Helmus & M. J. V. Zanden. 2009. The effects of impoundment and non–native species on a river food web in Mexico's central plateau. River Research and Applications 25 (9): 1090–1108. [ Links ]
Miller, R. R., W. L. Minckley & S. M. Norris. 2005. Freshwater Fishes of Mexico. The University of Chicago Press. U.S.A. 652 p. [ Links ]
Miller, R. R., W. L. Minckley & S. M. Norris. 2009. Peces Dulceacuícolas de México. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad/El Colegio de la Frontera Sur/Sociedad Ictiológica Mexicana, A.C. /Desert Fishes Council. México. 559 p. [ Links ]
Myers, G. S. 1940. Freshwater fishes and West Indian zoogeography. Annual Report Smithsonian Institute 1937: 339–364. [ Links ]
NOM–059–SEMARNAT–2010 (2010). Norma Oficial Mexicana. Protección Ambiental–Especies nativas de México de flora y fauna silvestres–categorías de riesgo y especificaciones para su inclusión, exclusión o cambio–Lista de especies en riesgo. Diario Oficial de la Federación. 30 de diciembre de 2010. 78 p. [ Links ]
Olden, J. D. & N. L. Poff. 2005. Long–term trends of native and non–native fish faunas in the American Southwest. Animal Biodiversity and Conservation 28 (1): 75–89. [ Links ]
Paulo–Maya, J. 2000. Situación actual sobre el conocimiento de la biología del género Chirostoma Swaison (Pisces: Ahterinopsidae). Ejercicio Predoctoral, Doctorado en Biología, Instituto Politécnico Nacional, México. 174 p. [ Links ]
Pegg, M. A. & M. A. McClelland. 2004. Spatial and temporal patterns in fish communities along the Illinois River. Ecology of Freshwater Fish 13: 125–135. [ Links ]
Penczak, T., A. A. Agostinho, l. C. Gomes & J. D. Latini. 2009. Impacts of a reservoir on fish assemblages of small tributaries of the Corumbá River, Brazil. River Research and Applications 25: 1013–1024. [ Links ]
Quist, M. C., W. A. Hubert & F. J. Rahel. 2005. Fish assemblage structure following impoundment of a Great Plains river. Western North American Naturalist 65: 53–63. [ Links ]
Rojas, M. J. R. & O. Rodríguez S. 2008. Diversidad y abundancia ictio–faunística del río Grande de Térraba, sur de Costa Rica. Revista de Biología Tropical 56 (3): 1429–1447. [ Links ]
Schlosser, I. J. & P. L. Angermeier. 1995. Spatial variation in demographic processes of lotic fishes: conceptual models, empirical evidence, and implications for conservation. In: Nielsen J. L. (Ed.). Proceedings of the Symposium on Evolution and the Aquatic Ecosystem: Defining Unique Units in Population Conservation. Monterey, California, USA, 23–25 May 1994. American Fisheries Society Symposium No. 17, Bethesda, Md. pp. 392–401. [ Links ]
Soto–Galera, E., E. Díaz–Pardo, E. López–López & J. Lyons. 1998. Fish as indicators of environmental quality in the Rio Lerma Basin, Mexico. Aquatic Ecosystem Health and Management 1 (3): 267–276. [ Links ]
Tamayo, J. L. 1996. Geografía Moderna de México. Edit. Trillas, Cd. de México. 400 p. [ Links ]
Vörösmarty, C. J., P. B. McIntyre, M. O. Gessner, D. Dudgeon, A. Prusevich, P. Green, S. Gliden, S. E. Bunn, C.A. Sullivan, C. Residy–Liermann & P. M. Davies. 2010. Global threats to human water security and river biodiversity. Nature 467: 555–561. [ Links ]
The Report of The World Commission on Dams (WCD). 2000. Dams and Development: A New Framework for Decision–Making. UNEP. Earthscan, London, UK. 356 p. [ Links ]

