




![Erratum to Structural analysis of a relay ramp in the Querétaro graben, central Mexico: Implications for relay ramp development [Rev. Mex. Cienc. Geol., 28 (2011), 275-289]](/img/es/next.gif)





Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versión On-line ISSN 2007-2902versión impresa ISSN 1026-8774
Rev. mex. cienc. geol vol.30 no.3 Ciudad de México dic. 2013
Taxonomy and notes on the paleobiology of the late Pleistocene (Rancholabrean) antilocaprids (Mammalia, Artiodactyla, Antilocapridae) from the state of Hidalgo, central Mexico
Taxonomía y notas sobre la paleobiología de antilocápridos (Mammalia, Artiodactyla, Antilocapridae) del Pleistoceno tardío del Estado de Hidalgo, centro de México
Victor M. Bravo-Cuevas1*, Eduardo Jiménez-Hidalgo2, Miguel A. Cabral-Perdomo1, and Jaime Priego-Vargas3
1 Museo de Paleontología, Area Académica de Biología, Universidad Autónoma del Estado de Hidalgo, Ciudad del Conocimiento, Carretera Pachuca-Tulancingo km 4.5, CP 42184, Pachuca, Hidalgo, México. *vbravo@uaeh.edu.mx.
2 Laboratorio de Paleobiología, Instituto de Recursos, Campus Puerto Escondido, Universidad del Mar, Carretera Puerto Escondido-Oaxaca km 2.5, CP 71980, Oaxaca, México.
3 Maestría en Ciencias en Biodiversidad y Conservación, Area Académica de Biología, Universidad Autónoma del Estado de Hidalgo, Ciudad del Conocimiento, Carretera Pachuca-Tulancingo km 4.5, CP 42184, Pachuca, Hidalgo, México.
Manuscript received: July 9, 2013
Corrected manuscript received: October 20, 2013
Manuscript accepted: October 22, 2013
ABSTRACT
A set of cranial and dental material of Antilocapridae was recovered from Quaternary deposits that outcrop in southeastern Hidalgo, central Mexico. The fossil bearing unit consists of clay, silt, sand, and scarce conglomeratic lenses in a fluvial environment; the specimens are associated with Bison indicating a Rancholabrean Land Mammal Age. The comparative study of the material with selected specimens of North American antilocaprids, allowed its identification as the small-sized species Capromeryx minor and the medium-sized Stockoceros conklingi. The sample referable to C. minor shows a combination of cranial and dental characters typical of this species, including a large posterior tine that is curved upward and forward, posterior horn core with a deep sulcus on its posterior side that extends from the base to the tip, and well-developed heel on M3. The isotopic analysis of teeth referable to this species from Hidalgo indicates a mixed feeding dietary behavior. A partial skull and associated horn core fragments were assigned to Stockoceros conklingi, considering its larger size than C. minor and shorter than the extant pronghorn Antilocapra americana, as well as for having symmetrical and diverging horn cores, and more outwardly flaring horn cores. The recognition of these antilocaprids in southeastern Hidalgo represents one of the southernmost occurrences within temperate North America. The tiny Capromeryx minor is known in North American localities of lower altitude (0-100 m a.s.l.) compared to those of the medium-sized Stockoceros conklingi (1000-2000 m a.s.l.). Hence, body size differences between both species and altitude differences between some of their localities suggests minimal direct competition between these antilocaprids.
Key words: Antilocapridae, Capromeryx, Stockoceros, late Pleistocene, Hidalgo, central Mexico.
RESUMEN
En el presente estudio se describe un conjunto de restos craneales y dentales de Antilocapridae recuperado de depósitos del Cuaternario, que afloran en el sector suroriental del estado de Hidalgo, centro de México. La unidad portadora de los fósiles consiste en un conjunto de arcilla, limo, arena y escasos lentes conglomeráticos depositados en un ambiente fluvial; los ejemplares están asociados a restos pertenecientes al género Bison, fósil estratigráfico del Rancholabreano en la cronología de mamíferos. El estudio comparado de la muestra disponible con ejemplares seleccionados de antilocápridos de Norteamérica reveló la existencia de la especie de tamaño pequeño Capromeryx minor y de la especie de tamaño mediano Stockoceros conklingi. El material asignado a C. minor muestra una combinación de caracteres craneales y dentales propia de esta especie, incluyendo un cuerno posterior grande que se curva ligeramente hacia delante y hacia arriba y con un surco profundo a lo largo del margen posterior de la base hacia la punta, así como la presencia de un talón bien desarrollado sobre el M3. El análisis isotópico de material dental asignado a esta especie indica que tuvo un hábito alimentario mixto. Un cráneo parcial y restos de cuernos asociados se asignaron a Stockoceros conklingi, dado que el material corresponde a una especie más grande que C. minor y menor en tamaño al berrendo actual Antilocapra americana, la cual a su vez se distingue por presentar cuernos simétricos y divergentes que además se dirigen hacia fuera con respecto a la línea media del cráneo. El reconocimiento de estas especies de antilocápridos en el sureste de Hidalgo, representa uno de sus registros más australes para Norteamérica Templada. La especie pequeña C. minor se conoce de localidades en Norteamérica ubicadas en un rango altitudinal de 0-100 m s.n.m., mientras que aquellas donde se conoce S. conklingi están ubicadas en un rango altitudinal de 1000-2000 m s.n.m. Con base en esto, las diferencias en tamaño corporal y de rango altitudinal de las localidades de donde se conocen estos antilocápridos, sugieren que la competencia ecológica entre ellas no fue tan intensa.
Palabras clave: Antilocapridae, Capromeryx, Stockoceros, Pleistoceno Tardío, Hidalgo, centro de México.
INTRODUCTION
Antilocaprids are endemic North American ruminants that appeared in the fossil record during the Early Miocene (Late Hemingfordian). Today the Family Antilocapridae is represented only by Antilocapra americana, the pronghorn antelope, but in the past these artiodactyls constituted a very successful lineage with as many as six genera roaming North America during the Late Miocene (Janis and Manning, 1998; Davis, 2007).
Mexican antilocaprids ranged from the middle Miocene to the late Pleistocene. The oldest reported genus is Merycodus, from the middle Miocene (Barstovian) of southern Mexico (Jiménez-Hidalgo et al., 2002). Pliocene pronghorns are the best known and have been recorded mainly from central Mexico; seven genera have been reported thus far, ranging from Hemphillian 2 to Blancan III NALMA's (Jiménez-Hidalgo et al., 2004; Jiménez-Hidalgo and Carranza-Castañeda 2004; Jiménez-Hidalgo and Carranza Castañeda, 2011; Carranza-Castañeda et al., 2013).
The Pleistocene Mexican record of Antilocapridae includes Stockoceros sp. and Tetrameryx sp. from the Irvingtonian of El Golfo, Sonora (Shaw, 1981). Rancholabrean records include Antilocapra americana from Cuatro Ciénegas, Coahuila, Cueva Jiménez in Chihuahua, Minas in Nuevo León state and Laguna de la Media Luna in San Luis Potosí (Kurtén, 1975; Lundelius, 1980); Capromeryx minor from La Brisca, and Capromeryx sp. and Stockoceros sp. from Terapa, Sonora (Van Devender et al., 1985; Mead et al., 2006); Stockoceros conklingi from San Josecito Cave in Nuevo León, Mina San Antonio and La Presita in San Luis Potosí (Polaco and Butron, 1997; Arroyo-Cabrales and Jhonson, 2008). Capromeryx mexicanus, Stockoceros conklingi, Tetrameryx mooseri and Tetrameryx tacubayensis were recorded from the El Cedazo Local Fauna, Aguascalientes (Mooser, 1958, Mooser and Dalquest, 1975); Capromeryx mexicanus and Stockoceros conklingi from Tequixquiac, State of Mexico (Furlong, 1925; Maldonado-Koerdell, 1955); Stockoceros conklingi and Tetrameryx shuleri from Hueyatlaco, Puebla (Guenther and Bunde, 1973); and Capromeryx mexicanus, Stockoceros conklingi and Tetrameryx shuleri from Mixtequilla in Veracruz (Polaco, 1995).
Paleontological research in Pleistocene localities from Hidalgo has allowed the recovery of antilocaprid specimens that represent two species. The purpose of this paper is to describe these specimens and to comment about the taxonomic status of some Pleistocene species of antilocaprids from Mexico and comment on some paleobiological aspects of the identified species.
STUDY AREA
The material was recovered from two nearby localities in the southeastern sector of Hidalgo. The localities are formally known as El Barrio (HGO-47: 20°07'41"N -98°56'02.7"W; 2184 m a.s.l.) and Barranca del Berrendo (HGO-28: 20°01'20.7"N - 98°37'37.9"W; 2460 m a.s.l.) (Figure 1). Outcrops of Tertiary to Quaternary volcanic and volcano sedimentary rocks are recognized in the region (INEGI, 1992). The fossil bearing unit consists of clay, silt, and sand moderately consolidated, including scarce conglomeratic lenses along the exposure, in a fluvial depositional environment (Figure 2).
The associated mammalian fauna includes rodents, bears, dire wolves, giant armadillos, giant ground sloths, llamas, deers, bison, mammoths, and mastodons (Castillo-Cerón et al., 1996; Bravo-Cuevas, 2001, 2002; Bravo-Cuevas et al., 2009, 2011, 2012; Cabral-Perdomo, 2001). Material referable to the genus Bison has been recovered from both localities, including a horn core fragment (HGO-47) and several isolated teeth (HGO-28, HGO-47); the presence of this taxon is indicative of a Rancholabrean North American Land Mammal Age (Bell et al, 2004).
MATERIAL AND METHODS
Studied sample and taxonomic identification. The fossil material includes a partial skull, three skull fragments with horn cores, isolated upper and lower molariforms, and two mandible fragments. The material is housed at the Sección de Macromamíferos, Museo de Paleontología, Universidad Autónoma del Estado de Hidalgo (UAHMP), Hidalgo, México, under the numbers UAHMP-330, UAHMP-331, UAHMP-1117, UAHMP-1118, UAHMP-1119, UAHMP-1121, and UAHMP-3925.
The material was compared with selected specimens of antilocaprids housed at the vertebrate paleontology collections of the Natural History Museum of Los Angeles County and the George C. Page Museum of La Brea Discoveries (LACM), Los Angeles, California; Frick collections of American Museum of Natural History (F:AM); and with descriptions and illustrations of specimens housed at the New Mexico Museum of Natural History and Science (NMMNH); University of Kansas Museum of Natural History (KU); and the Paleobiology Collection of the UTEP Biodiversity Collections (formerly the Laboratory for Environmental Biology), Department of Biological Sciences and Centennial Museum, University of Texas at El Paso (UTEP).
Selected measurements of cranial elements are those of Skinner (1942: fig. 12, p. 190-191). The dental nomenclature is that of Bärmann y Rössner (2011). The crown height, anteroposterior length and transverse width were measured at the occlusal surface (Morgan and Morgan, 1995) using a digital caliper. All measurements are in mm.
The tooth wear stages of Jiménez-Hidalgo and Carranza-Castañeda (2011:535) were related to a particular individual category as follows: Stage I, very young individual; Stage II, young individual; Stage III, young-adult individual; Stage IV, adult individual; Stage V, old individual. We estimate the body mass using the ruminant's prediction equations of Janis (1990) for first lower molar length (FLML) and first molar area (FLMA).
Dietary characterization. The small-sized antilocaprid considered in present study is represented by dental elements. Thus, we selected three molars to characterize its dietary behavior by means of δ13C isotopic analysis in carbonate tooth enamel apatite. Tooth enamel flakes were extracted and chemically treated following MacFadden and Cerling (1996) and Koch et al. protocols (1997). The samples were analyzed by using a Finnigan MAT 253 mass spectrometer attached to a Finnigan Gas Bench II on-line gas preparation system within the Laboratorio Universitario de Geoquímica Isotópica (LUGIS), Universidad Nacional Autónoma de México. Each sample was dissolved in 100% orthophosphoric acid at 25 °C and during 54 hours to create CO2.
Isotopic values are expressed in standard δ-notation and are reported relative to the V-PDB standard (Pee Dee Belemnite Limestone) (after Craig, 1957). The carbon isotope values are related to a particular dietary preference as follows: values of δ13C < -10%o indicate a diet consisting primarily in C3 plants (= browsers); values of δ13C > -1% indicate a diet consisting primarily in C4 plants (= grazers); and values of δ13C ranging between -1% and -10% indicate a diet consisting of both C3/C4 plants (= mixed feeders) (Koch et al., 1992; Quade et al., 1992; Cerling et al., 1997). The percentage of C4 plants consumption was estimated using the mass balance equation of Koch et al. (2004).
Anatomical abbreviations. APL/apl, upper/lower anteroposterior tooth length; CH/ch, upper/lower crown height; d, deciduous; L, left; M/m, upper/lower molar; P/p, upper/lower premolar; R, right; TW/tw, upper/lower transverse tooth width.
Statistical abbreviations. N, sample size; x, sample mean; s, sample standard deviation; V, sample coefficient of variation; OR, sample observed range.
Other abbreviations. ICZN, International Code of Zoological Nomenclature; Ka, thousand years; Ma, million years; NALMA, North American Land Mammal Age.
SYSTEMATIC PALEONTOLOGY
Order Artiodactyla Owen, 1848
Suborder Ruminantia Scopoli, 1777
Family Antilocapridae Gray, 1866
Subfamily Antilocaprinae Brook, 1876
Tribe Stockoceratini Frick, 1937
Genus Capromeryx Matthew, 1902
Capromeryx minor (Taylor, 1911)
(Figures 3 - 5 (4), Tables 1 and 2)


Breameryx minor, Furlong, 1946: 138, Pl. 3 (fig. 6). Capromeryx mexicana, Furlong, 1925: 146 - 148, figs. 7-9. Breameryx mexicana, Furlong, 1946: 138, Pl. 3 (fig. 5).
Description.
Skull and horn cores. The specimen UAHMP-331 includes the dorsal rim of the orbit; a small portion of the frontal bone, the posterior horn core that is nearly complete (lacking an estimated 15 mm of the tip) and the anterior horn core is missing. The core-base is over the orbit and its long diameter is 22.27 mm. The horizontal diameter of the orbit at the lower limits of preservation is 25.59 mm. The posterior horn core is laterally flattened and oval-shaped in cross section; the horn core rises upward and towards the tip is gently curved; there is a deep sulcus on the posterior side of the horn core, which extends from the base to the tip (Figure 3).
Upper dentition. There is only a RM3 (UAHMP-3925); this tooth is hypsodont (CH along the mesostyle is 30.28 mm), and the styles and ribs are prominent. The protocone and metaconule are V-shaped and about same size. The fossae are simple and U-shaped. The paracone and metacone are separated and well differentiated. A well-developed heel is observed in this tooth (Figure 4).
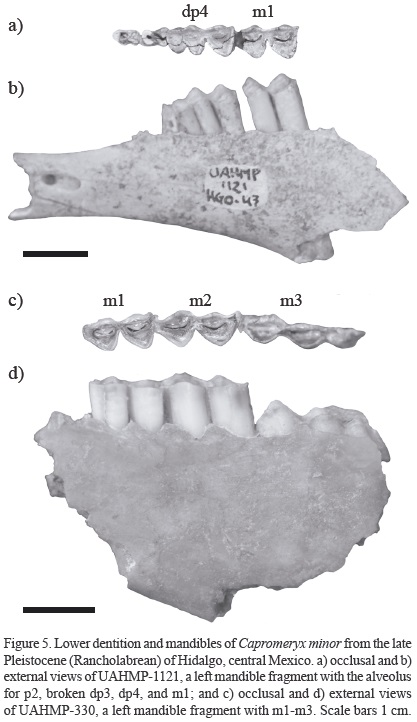
Mandible and lower dentition. The mandible fragment UAHMP-1121 is shallow and slender. The diastema is long and the diastemal crest is poorly developed. The mental foramen is large, deep, and oval in shape. In both mandible fragments (UAHMP-330 and UAHMP-1121) the mandibular depth increases backwardly (mean depth below m1 is 21.47 mm) (Figure 5).
The dp3 is broken and only the posterolingual column of the tooth is preserved; its occlusal pattern is indistinguishable. The dp4 is three-lobed; the anterolabial conid is U-shaped, whereas the protoconid and hypoconid are V-shaped; the anterolabial conid and the protoconid are similar in size, and the hypoconid is the largest conid of the labial border; the anterolingual conid is straight, the meta-conid is narrow and is well-separated from the entoconid, which is broader; the mesofossa and the posterior fossa are anteroposteriorly elongated, the mesofossa is constricted in its middle part and is smaller than the posterior fossa; the entostylid is slightly prominent (Figure 5a).
The m1's are hypsodont (mean ch ≈ 26 mm) and two-lobed; the protoconid and hypoconid are V-shaped, and similar in size; the anterior and posterior fossae are anteroposteriorly elongated and constricted in their middle region; the mesostylid and entostylid are prominent; and the ribs are well-developed. The occlusal pattern of m2 is comparable to that of ml, but this tooth is larger. The m3 is three-lobed, a well-developed talonid is observed, and this tooth is beginning to erupt (Figure 5a and 5c).
Age categories. The specimen UAHMP-1121 is considered in the tooth wear Stage I, because it shows the deciduous premolars (dp3 and dp4), suggesting a very young individual. The specimen UAHMP-330 is considered in the tooth wear Stage II, because the m1 is slightly worn and the m3 begin to erupt, which in turn suggests a young individual.
Referred material. Loc. El Berrendo (HGO-28): UAHMP-330, a left mandible fragment with m1-m3; UAHMP-331, skull fragment with the posterior horn core. Loc. El Barrio (HGO-47): UAHMP-1121, a left mandible fragment with the alveolus for p2, broken dp3, dp4, and m1; UAHMP-3925, isolated RM3.
Age and occurrence. Chronological range restricted to the Rancholabrean NALMA. The species is known from several localities in California, Texas, and New Mexico, USA (Davis, 2007); and the Mexican states of Sonora (Rancho La Brisca) (Van Devender et al., 1985), State of Mexico (Tequixquiac) (Furlong, 1925; Maldonado-Koerdell, 1955) and Hidalgo (El Berrendo and El Barrio localities) (this report).
Discussion.
Taxonomic status of late Pleistocene Capromeryx species. It has been shown that Capromeryx underwent a progressive diminution in body size through the Pliocene and Pleistocene (Morgan and Morgan, 1995). The trend towards size reduction is reached in the Rancholabrean small-sized species C. furcifer, C. minor, and C. mexicanus.
Recently, White and Morgan (2011) described material from the Rancholabrean of New Mexico as belonging to C. furcifer. The taxonomic assignment of the sample is extensively discussed, showing that the horn core morphology of this species is clearly distinctive from the other Rancholabrean small-sized species.
The taxonomic status and features employed to distinguish C. minor from C. mexicanus are controversial (see Hibbard and Taylor, 1960: 188; Morgan and Morgan, 1995: references therein; White and Morgan, 2011:646). The species C. minor was named by Taylor (1911) to distinguish mandible fragments and postcranial elements belonging to a small antilocaprid from Rancho La Brea. Later, Chandler (1916) described additional specimens, including cranial material.
On the other hand, the species C. mexicanus was subjectively defined by Furlong (1925) based on a partial skull of an old individual (Holotype, UCMP 26648) recovered from Tequixquiac, State of Mexico; additional material from Tequixquiac and El Cedazo (Aguascalientes, central Mexico) was assigned acritically to this species (Maldonado-Koerdell, 1955; Mooser and Dalquest, 1975).
A comparison of UCMP 26648 with cranial material belonging to C. minor from La Brea shows that diagnostic features of C. mexicanus are arguable. The specimen UCMP 26648 shows a slight expansion of the tines just above the base from which the horn cores rise. Some specimens of C. minor have a similar appearance, such as LACMHC 49, which has robust and laterally expanded horn cores with the widest area distal to the base.
Furlong (1925) considered that anterior horn cores of C. mexicanus are relatively longer than in C. minor. Nevertheless, the long diameter of the anterior horn core of UCMP 26648 (21.43 mm) and the C. minor specimens LACMHC Z8523 (22.43 mm) and LACMHC 49 (23.75 mm) is ≈ 22 mm, with a difference of about 2 mm.
A forward curvature of the posterior horn core has been considered as a remarkable feature of C. mexicanus, although we observed that specimens of C. minor show a similar condition. The resemblance is particularly apparent with LACMHC 49 and LACMHC 1126 (skull of a composite skeleton that is on the exhibit mount in the Page Museum), given that in both specimens the posterior margin of the posterior horn core shows a forward curvature that begins above the line of union of the two horn cores and extends upwardly and forwardly as in UCMP 26648. It should be noted that forward curvature of UACMP 26648 is more pronounced (Furlong, 1925: 147, fig. 8), a condition that could be explained by age variation. This contention is in agreement with differences observed between horn cores of young and adult animals of the medium-sized antilocaprid Stockoceros conklingi, considering that the inclination of horn cores is most pronounced in old animals of this species (Furlong, 1943). Also, difference in inclination can be the result of intraspecific variation.
The comparison shows that prong morphology of C. minor and C. mexicanus from Tequixquiac (State of Mexico) is very similar or the same taking into account individual variation.
Therefore, we proposed that the Mexican material described and figured by Furlong (1925) as C. mexicanus, should be referred as C. minor in accordance with the Statement of the Principle of Priority (ICZN, 1999: Article 23). Other authors had adopted a similar taxonomic criterion, considering UCMP 26648 as the type of C. minor (see White and Morgan, 2011: 646). We are not prepared to argue that material referable as to C. mexicanus from El Cedazo has to be considered also as C. minor pending a formal revision of this material.
Taxonomic assignment. The sample from Hidalgo shows several diagnostic features of Capromeryx, such as horn cores supraorbitally located and parallel to each other, posterior horn core laterally flattened, deep sulcus in the posterior side of the posterior horn core, and m3 lacking a fourth lobe (Jannis and Manning, 1998).
Based on the Rancholabrean age of the strata and the small size of the taxon considered, the comparative study was restricted to the species Capromeryx minor and C. furcifer.
The cranial material from Hidalgo (UAHMP-331) described in this report shares with the tiny antilocaprid Capromeryx minor the following suite of diagnostic features: (1) horn cores arise from a common base; (2) the posterior horn core is long, slender and circular to oval in cross section; and (3) a posterior horn core that is bending upward and forward (Furlong, 1925; Kurten and Anderson, 1980). The size of the posterior horn core of UAHMP-331 is closely comparable to LACMHC 49 (a horn core) from La Brea, and it is slightly smaller (≈ 15%) to that of LACMHC Z8523 (left horn core) from La Brea and UCMP 26648 (= holotype) from Tequixquiac, State of Mexico (Table 1). These subtle differences in size can be explained by intraspe-cific variation related to age and/or sex of the individuals.
Mooser and Dalquest (1975) described a complete horn core (FC 626) from El Cedazo that was assigned to C. mexicanus (op. cit., 1975: 814). The maximum length estimated for this specimen ( ≈ 69 mm) is similar to that of UAHMP-331 (≈ 65 mm); nevertheless, our comparison is limited considering that the horn core morphology of the material from Aguascalientes is improperly described and it has not been figured.
The length of posterior horn core of UAHMP-331 is smaller (about 40%) than specimens belonging to Capromeryx furcifer, such as NMMNH 50387 and UTEP 9366.G.989.337.1 from the Rancholabrean of New Mexico (Table 1). The specimen from Hidalgo also has a posterior horn core curved upward and forward, quite unlike the straight-angled posterior horn cores of C. furcifer. Furthermore, this species differs in having posterior horn cores with a shallow groove on the anterior margin (White and Morgan, 2011).
The dental size and morphology between the small-sized species of Capromeryx is relatively homogeneous, with only few features, such as the depth of posterolingual inflection on p4 and the development of a heel on M3, showing some taxonomically significant variation (see Morgan and Morgan, 1995; Jiménez-Hidalgo et al., 2004). The size of the lower molars from Hidalgo is similar to that of dental elements belonging to C. furcifer from southwestern Kansas and is slightly larger than that of lower molars of C. minor from La Brea. Nevertheless, the lengths and widths of the sample from Hidalgo are within the observed range of the species considered in our comparison. It should be noted that an overlapping of the compared datasets is observed, thus suggesting that dental size may be inadequate for taxonomic distinction of the sample considered in this study (Table 2).
The specimen UAHMP-3925 (RM3) from Hidalgo shows a well-developed heel (Figure 4). This condition is commonly observed in the upper third molars of Capromeryx minor, such as the M3's of the specimen UCMP 26648 from Tequixquiac, State of Mexico (Furlong, 1925:148, fig. 9), and an isolated RM3 catalogued as LACM15 from McKittrick Tar Seeps, Rancholabrean of California. In C. furcifer, the development of a heel on M3 varies from absent to well developed (Saysette, 1999).
The comparison of the cranial material from Hidalgo with selected species of Capromeryx allowed identifying it as C. minor. We observe that dental size and morphology does not seem to provide much useful taxonomic information to distinguish between the small-sized species of Capromeryx; although the development of heel on M3 of the sample from Hidalgo is typical of that observed in specimens referable to C. minor. Nevertheless, we consider that the mandibular fragments and dental elements belong to the same species, given their association with diagnostic cranial material.
Dietary behavior. The δ13C isotope values of Capromeryx minor from Hidalgo vary from -5.94% to -9.57%. A mean value of -8.06% suggests a mixed feeding behavior, consisting primarily of C3 plants and in lesser extent of C4 plants (30% of C4 plants consumption). An intermediate feeder characterization has been previously considered for antilocaprids from the Wisconsinan of New Mexico and Nevada, including specimens referable to Capromeryx (Connin et al., 1998). Here, we provide additional information regarding the feeding behavior of this tiny antilocaprid, which appears to have had a diet consisting of variable plant resources with preference for C3 plants.
Genus Stockoceros Skinner, 1942
Stockoceros conklingi (Stock, 1930)

?Tetrameryx conklingi: Stock, 1930:6, figs. 1 - 3.
Tetrameryx onusrosagris (=Stockoceros onusrosagris) (Roosevelt and Burden, 1934)
Description.
Skull and horn cores. The description of skull morphology is based on UAHMP-1117 that is the specimen best preserved. It shows a cranial displacement of 20 ° in relation to the sagittal plane. The frontal is expanded anteroposteriorly; its lower area is anterior to the orbit and continues upward acquiring a slightly convex surface. A large and rounded supraorbital foramen is located anterior and internal to the horn core base. The dorsal margin of the orbit is rounded and prominent, whereas the orbit is large (anteroposterior diameter is 40 mm) and rounded. The parietal is small and occupies an area that is rectangular in outline. The temporal extends downward and the temporal ridge is well differentiated. The external auditory meatus is large and rounded, directed upward and rearward; the anomalous position of this structure resulted from distortion of the skull (Figure 6a). The nuchal crest is prominent and trapezoidal. The external occipital protuberance is well differentiated from the nuchal crest. The occipital condyles are kidney-shaped. The foramen magnum is large, deep, and oval. The jugular apophyses are short, wide, and roughly rectangular. The left auditory bulla is preserved, which is large and inflated. The oval foramina are rounded and located anterior to the auditory bulla. The basilar part of the occipital bone is flattened and occupies an area more or less triangular in outline (Figure 6b).
The horn cores are divergent (angle of divergence is 65 °) and arise from a wide and low base that is parallel to the medial line of the skull. The long diameter of the horn cores is ~ 31 mm. The anterior horn core is nearly complete (lacking an estimated 10 to 15 mm of the tip), is flared outwardly and located over the orbit; this horn core is oval in cross section, the anterior and posterior margins are straight and directed upward and forward. The posterior horn core is located posteriorly to the orbit, this horn core is circular (UAHMP-1118) to oval (UAHMP-1117 and UAHMP-1119) in cross section, its anterior margin is straight and the posterior margin is convex, and it is directed upward. The specimen UAHMP-1117 is considered an adult individual, given that horn cores show a tendency to flare outwardly and a pronounced inclination (Furlong, 1943).
Referred material. Loc. El Barrio (HGO-47): UAHMP-1117, partial skull with the right anterior and posterior horn cores; UAHMP-1118, skull fragment with the right and left posterior horn cores; UAHMP-1119, skull fragment with the base of the right posterior horn core.
Age and occurrence. Chronological range restricted to the Rancholabrean NALMA. The species is known from Arizona (Ventana Cave and Papago Springs Cave) and New Mexico (Shelter Cave), USA (Davis, 2007); and the Mexican states of Nuevo León (San Josecito Cave) (Arroyo-Cabrales and Johnson, 2008), Veracruz (La Mixtequilla) (Polaco, 1995), San Luis Potosí (Mina San Antonio and La Presita localities) (Polaco and Butron, 1997), Aguascalientes (Cedazo) (Mooser, 1958; Mooser and Dalquest, 1975), Puebla (Hueyatlaco) (Guenther and Bunde, 1973), State of Mexico (Tequixquiac) (Furlong, 1925; Maldonado-Koerdell, 1955), and Hidalgo (El Barrio) (this report).
Discussion.
Taxonomía assignment. The specimen UAHMP-1117 shares with Stockoceros the presence of paired and diverging horn cores; horn cores circular to oval in cross section; horn cores arise from a low core base that is parallel to the medial line of the skull; anterior horn core more outwardly flaring than posterior; a large and rounded supraorbital foramen located anterior and internal to the horn core base; and large and inflated auditory bulla (Skinner, 1942). The genus Tetrameryx is distinguished from the Hidalgo material by its larger size, asymmetrical horn cores, and posterior horn core significantly larger (about twice) than anterior horn core (Kurtén and Anderson, 1980; Davis, 2007).
On the other hand, the only surviving antilocaprid Antilocapra americana differs from the studied sample in having a larger skull, lacking an anterior horn core, and posterior horn core laterally compressed and blade-like (Kurtén and Anderson, 1980).
The species of Stockoceros are S. conklingi and S. onusrosagris. The horn core morphology between these species is closely similar and they have been distinguished by its size. The species S. onusrosagris has been considered larger than S. conklingi (Roosevelt and Burden, 1934; Skinner, 1942; Kurtén and Anderson, 1980).
The anterior and posterior horn core diameters of UAHMP-1117 are similar to those of the specimens LACM (CIT) 3164 and LACM (CIT) 3005, belonging to S. conklingi from San Josecito Cave, Nuevo León, northern Mexico. On the contrary, the horn cores of the specimens F:AM 42534 and F:AM 42539, referable to S. onusrosagris from Papago Springs Cave in Arizona, are longer (about 35%) than the material from Hidalgo (Table 3).
The specimen UAHMP-1117 has larger horn cores than those of LACM (CIT) 3164 and LACM (CIT) 3005, with a mean difference of 30% (Table 3). The horn cores of UAHMP-1117 are smaller (about 15%) than those of F:AM 42539 and F:AM 42534 belonging to S. onusrosagris (Table 3).
The outward flaring of the horn cores of UAHMP-1117 is closely comparable to that observed in specimens of S. conklingi, while the horn cores of specimens belonging to S. onusrosagris show a greater outward flaring of the horn cores (Table 3).
The material from Hidalgo has an intermediate size between specimens of Stockoceros conklingi from San Josecito Cave and S. onusrosagris from Papago Springs Cave (Table 3). Furlong (1943) studied a large sample of S. conklingi from San Josecito Cave and Shelter Cave, as well as specimens of S. onusrosagris from Papago Springs Cave. He concluded that between these samples the size differences could be related to age variation, suggesting that there was not enough evidence to regard them as specifically distinct. Our comparison supports this contention, considering that differences between the samples from Hidalgo, San Josecito Cave and Papago Springs Cave are probably due to age variation, geographic variation, and/or clinal. Hence, we propose that Stockoceros conklingi and S. onusrosagris are the same species; thus the latter species should be considered as a junior synonym (ICZN, 1999: Article 23).
PALEOBIOLOGICAL SIGNIFICANCE
Geographic distribution
The biochronological range of Capromeryx minor and Stockoceros conklingi is restricted to the Rancholabrean (Kurtén and Anderson, 1980; Davis, 2007). By the late Pleistocene, the record of these antilocaprids mainly comes from localities within temperate North America between 20° and 40° Lat N. Its recovery in southeastern Hidalgo represents one of the southernmost evidences for a temperate climate in the Rancholabrean (Figure 7).

For Mexico, the tiny antilocaprid Capromeryx minor shows a scattered distribution, occurring in one locality at the Northwestern Plains and Sierras (Sonora) and one at the Trans-Mexican Volcanic Belt (State of Mexico). The medium-sized species Stockoceros conklingi is more widespread, having been recorded in several localities within the Sierra Madre Oriental (Nuevo León and San Luis Potosí), the Central Plateau (Aguascalientes), the Trans-Mexican Volcanic Belt (State of Mexico and Puebla), and the Gulf Coastal Plain (Veracruz) (Figure 7). The presence of both species in Hidalgo extends their distribution for eastern-central Mexico within the Trans-Mexican Volcanic Belt morphotectonic province.
Within North America, C. minor is recorded in localities that range from 0-100 m a.s.l. in the San Francisco Bay region, to 1500-2000 m a.s.l. in Nevada and Mexico. The localities where S. conklingi has been recorded range from 1000 to 2000 m a.s.l.
Paleoecology
Previous work demonstrated that the genus Capromeryx underwent a progressive diminution in body size, reaching its smallest form during the Rancholabrean (Morgan and Morgan, 1995; White and Morgan, 2011). The late Pleistocene species C. minor was a small-sized anti-locaprid that weighed about 10 kg (Kurten and Anderson, 1980). The estimations of body mass of specimens from Hidalgo using the length and area of the first molar differ in an interval of 15 kg. We observed a similar pattern with estimations for specimens of C. minor from Rancho La Brea (Table 4). It should be noted that other estimations for specimens of C. minor indicate a weight of 12-13 kg and 8-11 for specimens of C. mexicanus (Sayssette, 1999). These values are close to those derived by the first molar length. This species is the only very small ruminant artiodactyl known from the late Pleistocene mammalian assemblage in southeastern Hidalgo, central Mexico.

The species Capromeryx minor shows morphological and ecological features (such as small body size, simple horn cores, and variable diet), related to organisms that dwell in copses of trees and shrubs for protection of its predators (see Downs et al., 1959; Jarman, 1974). Bravo-Cuevas et al. (2011) suggested that the mammalian association of southeastern Hidalgo is indicative of a heterogeneous environment, consisting of open grassland areas with shrubs and trees. Hence, is probable that Capromeryx minor from the late Pleistocene of Hidalgo thrived along the transition zones of open and closed habitats. This is an appropriate scenario for having a selective diet. Our contention is in agreement with the mixed feeder category proposed for this species by means of isotopic analysis.
The antilocaprid Stockoceros conklingi weighed about 50 kg and had an intermediate size between Capromeryx minor and the existing pronghorn Antilocapra Americana (Kurtén and Anderson, 1980; Alroy, 2001). Analysis of dental wear patterns at micro- and macroscopic inspection indicates a mixed diet in Stockoceros onusrosagris (Rivals and Semprebon, 2006; Semprebon and Rivals, 2007).
It has been shown that Capromeryx minor and Stockoceros conklingi were antilocaprids significantly different in size. Hence, it seems probable that this condition reduced competition for food resources between these species from the late Pleistocene of southeastern Hidalgo, both considered to be selective feeders.
The presence of Capromeryx minor in North American localities of lower altitude (0-100 m a.s.l.) compared to those of Stockoceros conklingi (1000-2000 m a.s.l.), suggests that the tiny species was able to use some plant resources that were not suitable or not available to S. conklingi.
CONCLUSIONS
The comparative study of fossil antilocaprid material (cranial and dental) from the late Pleistocene of Hidalgo (Rancholabrean) recognizes the presence of the small-sized Capromeryx minor and the medium-sized Stockoceros conklingi. The record from Hidalgo represents one of the southernmost occurrences of these species within North America.
By means of isotopic analysis we characterize the small-sized Capromeryx minor from southeastern Hidalgo as a selective feeder. A similar dietary behavior is indicated for the medium-sized Stockoceros conklingi.
Although Capromeryx minor and Stockoceros conklingi were selective feeders, body size differences between the species and altitude differences between some of their localities suggest that competition for resources between these pronghorn was not strong.
ACKNOWLEDGMENTS
We are indebted to Richard S. White and to the anonymous reviewer for critically reviewing and improving the manuscript. We thank to Hannes Loeser, for his kindly support with the editorial process. Thanks to Pedro Morales Puente and Edith Cienfuegos Alvarado, of the Laboratorio Universitario de Geoquímica Isotópica, Universidad Nacional Autónoma de México (LUGIS-UNAM), for their help with the isotopic analysis procedure. The first author thanks Samuel A. McLeod and Vanessa R. Rhue, collection manager and curatorial assistant of the Vertebrate Paleontology collection at the Natural History Museum of Los Angeles County, as well as to Aisling Farrell, collection manager of the Page Museum, La Brea Tar Pits, for their kindly support during his visit to those paleontological collections. At last but not least we acknowledge G. Ruelas-Inzunza for reviewing the English. This study is part of the project CONACyT-Ciencia Básica (CB-2009-01, No. 130712) "El registro de ungulados terrestres en el Pleistoceno de Hidalgo, Puebla y Tlaxcala, centro de México: Interpretación de hábitos alimentarios y caracterización de paleocomunidades terrestres".
REFERENCES
Alroy, J., 2001, A multispecies overkill simulation ofthe end-Pleistocene megafaunal mass extinction: Science 292, 1893-1896. [ Links ]
Arroyo-Cabrales, J., Johnson, E., 2008, Mammalian additions to the faunal assemblages from San Josecito Cave, Nuevo León, México, in Espinosa, L.C.E., Ortega, J. (eds.), Avances en el Estudio de los Mamíferos de México: México, D.F., Asociación Mexicana de Mastozoología, A.C., Publicaciones Especiales II, 65-87. [ Links ]
Bärmann, E.V., Rössner G.E., 2011, Dental nomenclature in Ruminantia: Towards a standard terminological framework: Mammalian Biology, 76, 762-768. [ Links ]
Bell, C.J., Lundelius Jr., E.L., Barnosky, A.D., Graham, R.W., Lindsay, E. H., Ruez Jr., D.R., Semken Jr., H. A., Webb, S. D., Zakrzewski, R. J., 2004, The Blancan, Irvingtonian, and Rancholabrean mammal ages, in Woodburne, M.O. (ed.), Late Cretaceous and Cenozoic mammals of North America: New York, Columbia University Press, 232-314. [ Links ]
Bravo-Cuevas, V.M., 2001, Pleistocene faunal assemblage from south-central region of the state of Hidalgo, Central Mexico, in Sixty-First Annual Meeting Society of Vertebrate Paleontology, Museum of the Rockies Montana State University, Bozeman, Montana: Journal of Vertebrate Paleontology 21 (Supplement to No. 3), 35A. [ Links ]
Bravo-Cuevas, V.M., 2002, Diferenciación geológica y bioestratigráfica de la Formación Tarango para el estado de Hidalgo, Centro de México: México, Hidalgo, Universidad Autónoma del Estado de Hidalgo, Centro de Investigaciones en Ciencias de la Tierra, Technical Report, 59 pp. [ Links ]
Bravo-Cuevas, V.M., Cabral-Perdomo, M. A., Ortiz-Caballero, E., Priego-Vargas, J. 2009, La megafauna del Pleistoceno, in González-Rodríguez K., Cuevas-Cardona, C., Castillo-Cerón, J.M. (eds.), Los fósiles del estado de Hidalgo: México, Hidalgo, Universidad Autónoma del Estado de Hidalgo, 85-96. [ Links ]
Bravo-Cuevas, V.M., Jiménez-Hidalgo E., Priego-Vargas, J., 2011, Taxonomía y hábito alimentario de Equus conversidens (Perissodactyla, Equidae) del Pleistoceno Tardío (Rancholabreano) de Hidalgo, centro de México: Revista Mexicana de Ciencias Geológicas, 28 (1), 65-82. [ Links ]
Bravo-Cuevas, V.M., Jiménez-Hidalgo, E., Cuevas-Ruíz G., Cabral-Perdomo M.A., 2012, A small Hemiauchenia from the late Pleistocene of Hidalgo, central Mexico: Acta Palaeontologica Polonica, 57(3), 497-508. [ Links ]
Brook, V., 1876, Family Bovidae, in Russel, A.E. (ed.), The Geographical Distribution of Animals: London, MacMillan, 222-225. [ Links ]
Cabral-Perdomo, M.A., 2001, Vertebrados pleistocénicos de la región Pachuca-Tulancingo, estado de Hidalgo: México, Distrito Federal, Facultad de Ciencias, Universidad Nacional Autónoma de México, MSc. Thesis, 85 pp. [ Links ]
Carranza-Castañeda, O., Aranda-Gómez, J.J., Wang, X., Iriondo, A., 2013, The Early-Late Hemphillian (Hh2) faunal assemblage from Juchipila basin, state of Zacatecas, México, and its biochronologic correlation with other Hemphillian faunas in central Mexico: Contributions in Science, 521,13-49. [ Links ]
Castillo-Cerón, J.M., Cabral-Perdomo, M.A., Carranza-Castañeda, O., 1996, Vertebrados fósiles del Estado de Hidalgo: Hidalgo, México, Universidad Autónoma del Estado de Hidalgo, 127 pp. [ Links ]
Cerling, T.E., Harris, J.M., MacFadden, B.J., Leakey, M.G., Quade, J., Eisenmann, V., Ehlinger, J.R., 1997, Global vegetation change through the Miocene/Pliocene boundary: Nature, 389, 153-158. [ Links ]
Chandler, A.C., 1916, Notes on Capromeryx material from the Pleistocene of Rancho La Brea: University of California Publications, Bulletin of the Department of Geology, 9, 111-120. [ Links ]
Connin, S.L., Betancourt J., Quade J., 1998, Late Pleistocene C4 Plant Dominance and Summer Rainfall in the Southwestern United States from Isotopic Study of Herbivore Teeth: Quaternary Research, 50, 179-193. [ Links ]
Craig, H., 1957, Isotopic Standards for Carbon and Oxygen and correction factors for mass spectrometric analysis of carbon dioxide: Geochimica et Cosmochimica Acta 12, 133-149. [ Links ]
Davis, E.B. 2007, Family Antilocapridae, in Prothero, D.R., Foss, S.E. (eds.), The Evolution of Artiodactyls: Baltimore, Maryland, Johns Hopkins University Press, 227-240. [ Links ]
Downs T., Howard H., Clementz T., Smith G., 1959, Quaternary animals from Schuiling Cave in the Mojave River: Los Angeles County Museum, Contributions in Science, 29, 1-21. [ Links ]
Frick, C. 1937, Horned ruminants of North America: Bulletin of the American Museum of Natural History, 59, 505-652. [ Links ]
Furlong, E.L., 1925, Notes on the occurrence of mammalian remains in the Pleistocene of Mexico, with a description of a new species Capromeryx mexicana: University of California Publications in Geological Sciences, 15, 137-152. [ Links ]
Furlong, E.L., 1943, The Pleistocene antelope Stockoceros conklingi from San Josecito Cave, Mexico: Carnegie Institution of Washington Publication, 551, 1-8. [ Links ]
Furlong, E.L., 1946, Generic identification of the Pleistocene antelope from Rancho La Brea: Carnegie Institution of Washington Publication, 551, 135-140. [ Links ]
Gray, J.E., 1866, Notes on the pronghorn buck (Antilocapra) and its position in the system: Annals and Magazine of Natural History, 3, 323-326. [ Links ]
Guenther, E.W., Bunde, H., 1973, Investigaciones geológicas y paleontológicas en México durante los años de 1965 a 1969: Comunicaciones, 7, 19-20. [ Links ]
Hibbard, C.W., Taylor, D.W., 1960, Two late Pleistocene faunas from southwestern Kansas: The University of Michigan, Contributions from the Museum of Paleontology, 16(1), 1-223. [ Links ]
Instituto Nacional de Estadística, Geografía e Informática (INEGI), 1992, Síntesis Geográfica del estado de Hidalgo: México, Instituto Nacional de Estadística, Geografía e Informática, 134 pp. [ Links ]
International Commission on Zoological Nomenclature (ICZN), 1999, International code of zoological nomenclature (online): <http://www.iczn.org/iczn/index.jsp> [ Links ]
Janis, C.M., 1990, Correlation of cranial and dental variables with body size in ungulates and macropodids, in Damuth, J., MacFadden, B.J. (eds.), Body size in mammalian paleobiology: New York, Cambridge University Press, 255-299. [ Links ]
Janis, C.M., Manning, E., 1998, Antilocapridae, in Janis, C.M., Scott, K.M., Jacobs, L.L. (eds.), Evolution of Tertiary Mammals of North America. Terrestrial Carnivores, Ungulates, and Ungulatelike Mammals, v. 1: Cambridge, Cambridge University Press, 491-507. [ Links ]
Jarman, P.J., 1974, The social organization of antelope in relation to their ecology: Behavior, 48, 215-267. [ Links ]
Jiménez-Hidalgo, E., Carranza-Castañeda, O., 2004, Los antilocápridos del Terciario tardío de San Miguel de Allende, Guanajuato, in IX Congreso Nacional de Paleontología, Libro de Resúmenes: México, Sociedad Mexicana de Paleontología, p. 79. [ Links ]
Jiménez-Hidalgo, E., Carranza-Castañeda, O., 2011, The Pliocene pronghorn Hexobelomeryx fricki (Mammalia: Artiodactyla) from San Miguel de Allende, Guanajuato, central Mexico: Revista Mexicana de Ciencias Geológicas 28(3), 533-543. [ Links ]
Jiménez-Hidalgo, E., Ferrusquía-Villafranca, I., Bravo-Cuevas V.M., 2002, El registro mastofaunístico miocénico de México y sus implicaciones geológico-paleontológicas, in Montellano-Ballesteros, M., Arroyo-Cabrales, J. (eds.), Avances en los estudios paleomastozoológicos en México: México, Instituto Nacional de Antropología e Historia, INAH, Colección Científica, 47-68. [ Links ]
Jiménez-Hidalgo, E., Carranza-Castañeda, O., Montellano-Ballesteros, M, 2004, A Pliocene record of Capromeryx (Mammalia: Antilocapridae) in México: Journal of Paleontology, 78, 1179-1186. [ Links ]
Koch, P.L., Zachos J.C., Gingerich P.D., 1992, Correlation between isotope records in marine and continental carbon reservoirs near the Paleocene/Eocene boundary: Nature, 358, 319-322. [ Links ]
Koch, P. L., Tuross, N., Fogel, M. L., 1997, The effects of sample treatment and diagenesis on the isotopic integrity of carbonate in biogenic hydroxylapatite: Journal of Archaeological Science, 24, 417-429. [ Links ]
Koch, PL., Diffenbaugh N.S., Hoppe K.A., 2004, The effects of late Quaternary climate and pCO2 change on C4 plant abundance in the south-central United States: Palaeogeography, Palaeoclimatology, Palaeoecology, 207, 331-357. [ Links ]
Kurtén, B., 1975, A new Pleistocene genus of American mountain deer in North America: Journal of Paleontology, 56(2), 507-508. [ Links ]
Kurtén, B., Anderson, E., 1980, Pleistocene mammals of North America: New York, Columbia University Press, 442 pp. [ Links ]
Lundelius, E.L., 1980, Late Pleistocene and Holocene mammals from Northern Mexico and their implications for archaeological Research: Bulletin of the Florida State Museum, Biological Sciences, 27(1), 1-76. [ Links ]
MacFadden, B.J., Cerling, T.E., 1996, Mammalian herbivore communities, ancient feeding ecology, and carbon isotopes: a 10 million years sequence from the Neogene of Florida: Journal of Vertebral Paleontology, 16(1), 103-115. [ Links ]
Maldonado-Koerdell, M., 1955, Nuevos Hallazgos de antilocápridos en el Pleistoceno Superior de Tequixquiac, Mexico: Acta Zoologica Mexicana 1(7), 1-7. [ Links ]
Matthew, W.D., 1902, List of the Pleistocene fauna of Hay Springs, Nebraska: Bulletin of the American Museum of Natural History, 16, 317-322. [ Links ]
Mead, J.I., Baez, A., Swift, S.L., Carpenter, M.C., Hollenshead, M., Czaplewski, N.J., Steadman, D.W., Jordon, B., Arroyo-Cabrales, J., 2006, Tropical marsh and savanna of the Late Pleistocene in northeastern Sonora, Mexico: The Southwestern Naturalist, 51, 226-239. [ Links ]
Mooser, B.O., 1958, La fauna "Cedazo" del Pleistoceno en Aguascalientes: Universidad Nacional Autónoma de México, Anales del Instituto de Biología, 1-2, 409-452. [ Links ]
Mooser, O., Dalquest, W.W., 1975, Pleistocene mammals from Aguascalientes, Central Mexico: Journal of Mammalogy, 56, 781-820. [ Links ]
Morgan, J.K., Morgan, N.H., 1995, A new species of Capromeryx (Mammalia: Artiodactyla) from the Tauton Local Fauna of Washington, and the correlation with other faunas of Washington and Idaho: Journal of Vertebrate Paleontology, (15)1, 160-170. [ Links ]
Owen, R., 1848, Description of teeth and portions of jaws of two extinct anthracotherioid quadrupeds (Hyopotamys vectianus and Hyopotamys bovinus) discovered by the Marchioness of Hasting in the Eocene deposits on the N.W. coast of the Island of Wright: With an attempt to develop Cuvier's idea of the classification of pachyderms by the number of their toes: Quarterly Journal of the Geological Society of London, 4:103-141. [ Links ]
Polaco, O.J., 1995, Análisis de la arqueofauna de la Mixtequilla: Instituto Nacional de Antropología e Historia, Laboratorio de Paleozoología, Technical Report Z-471. [ Links ]
Polaco, O.J., Butron, M.L., 1997, Mamíferos Pleistocénicos de la Cueva la Presita, San Luis Potosí, México, in Arroyo-Cabrales, J., Polaco, O.J. (eds.), Homenaje al Profesor Ticul Alvarez: México, Instituto Nacional de Antropología e Historia, Colección Científica 279-376. [ Links ]
Quade, J., Cerling T.E., Barry J.C., Morgan M.E., Pilbeam D.R., Chivas A.R., Lee-Thorp J.A., van de Merwe N. J., 1992, A 16-Ma record of paleodiet using carbon and oxygen isotopes in fossil teeth from Pakistan: Chemical Geology (Isotope Geoscience Section), 94, 183-192. [ Links ]
Rivals, F., Semprebon, G. M., 2006, A comparison of the dietary habits of a large sample of the pleistocene pronghorn Stockoceros onusrosagris from the Papago Springs Cave in Arizona to the modern Antilocapra Americana: Journal of Vertebrate Paleontology, 26(2), 495-500. [ Links ]
Roosevelt, Q., Burden J.W., 1934, A new species of antilocaprine, Tetrameryx onusrosagris, from a Pleistocene Cave deposit in southern Arizona: American Museum Novitates, 754, 1-4. [ Links ]
Saysette, J.E., 1999, Postcranial estimators of body mass in pecorans with emphasis on Capromeryx (Mammalia: Artiodactyla): Denver, USA, Colorado State University, PhD Dissertation, 69 pp. [ Links ]
Scopoli, G.A., 1777, Introductio ad historiam naturalem, sistens genera lapidum, plantarum et animalium hactenus detecta, caracteribus essentialibus donata, in tribus divisa, subinde ad leges naturae: Prague, Apud Wolfgangum Gerle, 506 pp. [ Links ]
Semprebon, G., Rivals F., 2007, Was grass more prevalent in the pronghorn past? An assessment of the dietary adaptations of Miocene to Recent Antilocapridae (Mammalia: Artiodactyla): Palaeogeography, Palaeoclimatology, Palaeoecology, 253, 332-347. [ Links ]
Shaw, C.A., 1981, The Middle Pleistocene El Golfo Local Fauna from northwestern Sonora, Mexico: California, USA, California State University, Long Beach, MSc Dissertation, 141 pp. [ Links ]
Skinner, M.F., 1942, The fauna of Papago Springs Cave, Arizona and a study of Stockoceros with three new antilocaprines from Nebraska and Arizona: Bulletin of the American Museum of Natural History, 80, 143-220. [ Links ]
Stock, C., 1930, Quaternary antelope remains from a second cave deposit in the Oregon Mountains, New Mexico: Los Angeles Museum Paleontology Publications, 2, 1-18 [ Links ]
Taylor, W.P.A., 1911, New antelope from the Pleistocene of Rancho La Brea: University of California Publications, Bulletin of the Department of Geology, 6, 191-197. [ Links ]
Van Devender, T.R., Rea, A.M., Smith, M.L., 1985, The Sangamon interglacial vertebrate fauna from Rancho La Brisca, Sonora, Mexico: Transactions of the San Diego Society of Natural History, 21 (2), 23-55. [ Links ]
White, R.S. Jr., Morgan, G.S., 2011, Capromeryx (Artiodactyla: Antilocapridae) from the Rancholabrean Tramperos Creek Fauna, Union County, New Mexico, with a review of the occurrence and paleobiology of Capromeryx in the Rancholabrean of New Mexico, in Sullivan, R.M., Lucas, S.G., Spielmann, J.A. (eds.), Fossil Record 3: New Mexico, New Mexico Museum of Natural History and Science, Bulletin 53, 641-651. [ Links ]

