











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCTION
The Tlayúa quarry is a remarkable Early Cretaceous Konservat-Lagerstätte locality (Applegate et al., 2006; Alvarado-Ortega et al., 2007), near Tepexi de Rodriguez town, Puebla State, central Mexico (Figure 1). The fossil bearing lithographic limestones of this site belong to the Middle Member of the Tlayúa Formation of Albian age (Pantoja-Alor, 1992; Benammi et al., 2006), which were deposited under shallow marine conditions with sporadic freshwater influences, into a tropical and semi-enclosed coastal lagoon (Espinosa-Arruebarrena and Applegate, 1996). The rich fossil assemblage recovered in this site represents algae, foraminifers, ostracods, sponges, corals, gastropods, arthropods, ammonites, and other invertebrates, as well as plant remains, reptiles, and fishes (Applegate et al., 2006; 2009.
Since 1981, the Tlayúa quarry has been the focus of a regional paleontological project led by researchers and students of the Instituto de Geología of UNAM (Universidad Nacional Autónoma de México). In this project, two members of the Aranguthy Contreras family, owners and workers of this quarry, were hired by the UNAM to support in the recovering of fossils from this and others fossil localities nearby. They deliver all these fossils to be housed into the Colección Nacional de Paleontología (IGM). Thanks to this project, today we account with a large collection of Tlayúa fossils with more than 7,000 macrofossils, of which about 5% are clupeomorphs.
In two extensive works to order the previous chaotic Superorder Clupeomorpha, Grande (1982, 1985) recognized two groups, simply named as Divisions 1 and 2. Although the validity of his Division 1 is currently untenable (Arratia, 1996; Hermus and Wilson, 2001; Hermus et al., 2004; Forey, 2004), his Division 2 represented by the orders Clupeiformes and Ellimmichthyiformes plus ArmigatusGrande, 1982, has proved to be more consistent. Grande (1982) named the order Ellimmichthyiformes, a group commonly called double armored herrings, to include some extinct Early Cretaceous-Eocene clupeomorphs. In addition to a mosaic of primitive characters, this order is well characterized by a row of scutes with large distinctive lateral wings, the so called predorsal scutes series, that is located between the occiput and the dorsal fin.
When it was named, the composition of the Ellimmichthyiformes was a comparatively simple task because this order taxon only included five species, representing three different genera gathered in a single family (Grande, 1982, p. 22): (1) The type genus, then represented by a single species, Ellimmichthys longicostatus (Cope, 1886); (2) three species of DiplomystusCope, 1877 (D. dentatusCope, 1877; D. birdiWoodward, 1895; and D. dubertretiSigneux, 1951); and (3) Armigatus brevissimus (Blainville, 1818), whose position within the Ellimmichthyiformes has been a problem because the shape of its predorsal scutes is rather ovoid than winged (Zaragüeta-Bagils, 2004; Alvarado-Ortega et al., 2008; Murray and Wilson, 2013; among others). Currently, this taxonomic framework has become much more complex after the application of phylogenetic methodologies and the increasing number of species recently described (Figure 2). Today this order is represented by at least 35 nominal species belonging to 18 genera (Table 1) (also, see Taverne, 1997; Bannikov and Bacchia, 2000; Chang and Maisey, 2003; Zaragüeta-Bagils, 2004; Forey, 2004; Malabarba et al., 2004; Alvarado-Ortega et al., 2008; Newbrey et al., 2010; Bannikov, 2015; Vernygora and Murray, 2016; Vernygora et al., 2016; Marramà and Carnevale, 2016; Figuereido and Ribeiro, 2016, 2017; among others).

Figure 2 Composition and relations of the Ellimmichthyiformes according to different authors (the taxa currently considered part of this order are indicated by the black circles on the cladograms; the species names were abbreviated with the first two letters; the specific names were omitted in the monospecific genera).
Table 1 Nominal species already included in the order Ellimmichthyiformes (based on Grande, 1985; Zaragüeta-Bagils, 2004; Alvarado-Ortega et al., 2008; Murray and Wilson, 2013; Marramà and Carnevale, 2016; Nelson et al., 2016; among others).
| Species | Age | Distribution |
|---|---|---|
| Armigatus brevissimus (Blainville, 1818). | Cenomanian | Heckel and Hajula, Lebanon |
| Armigatus alticorpusForey, Yi, Patterson, and Davies, 2003. | Cenomanian | Namoura; Lebanon |
| Armigatus namouroensisForey, Yi, Patterson, and Davies, 2003. | Cenomanian | Namoura; Lebanon |
| Armigatus oligodentatus Vernigora and Murray, 2016. | Cenomanian / Turonian | Agoult, Morocco |
| Armigatus dalmaticusMurray, Vernygora, Japundžić, Radovčić, Wilson, Bardack, and Grande, 2016. | Campanian? | Dalmatia, Croatia |
| Codoichthys carnavaliiSilva-Santos, 1994. | Aptian | São Luís-Grajaú Basin, Brazil |
| Diplomystus dentatusCope, 1877. | Eocene | Green River Fm., USA |
| Diplomystus birdiWoodward, 1895. | Cenomanian | Heckel and Hajula, Lebanon |
| Diplomystus dubertretiSigneux, 1951. | Senonian | Sahel Alma, Lebanon |
| Diplomystus solignaciGaudant and Gaudant, 1971. | Senonian | Gabès, Tunisia |
| Diplomystys shengliensisZhang, Zhou, Qin, 1985. | Eocene | Shandong, Chiana |
| Ellimma branneri (Jordan, 1910, 1913). | Hauterivian -Aptian | Muribeca Fm., Sergipe, Brazil. |
| Ellimma cruziSilva-Santos 1990. | Aptian | Cabo Fm. Pernambuco, Brazil |
| Ellimmichthys longicostatus (Cope, 1886). | Hauterivian-Barremian | Recôncavo Fm., Bahía, Brazil |
| Ellimmichthys goodiEastman, 1912. | Aptian-Albian | Cocobeach Series, Equatorial Guinea |
| Ellimmichthys maceioensisMalabarba, Do Carmo, Gómez-Pérez, and Queiroz-Neto, 2004. | Aptian-Albian | Maceio Fm., Sergipe-Alagoas Basin, Brazil |
| Ezkutuberezi carmenaePoyato-Ariza, López-Horgue, and García-Garmilla, 2000. | Valanginian-Barremian | Villaro Fm., Arratia Valley, Spain |
| Foreyichthys loonensis Vernygora, Murray, and Wilson, 2016. | Albian | Loon River Fm. Northwest Territories, Canada |
| Gateroclupea branisaiSigneux, 1964. | Late Cretaceous-Paleocene | Cayara, Bolivia |
| Horseshoeichthys armaserratusNewbrey, Murray, Brinkman, Wilson, and Neuman, 2010. | Maastrichthian | Horseshoe Canyon Fm., Alberta, Canada |
| Kwangoclupea dartevellei (Casier, 1965). | Cenomanian | Kwango, Zaire |
| Paraclupea chetungensisSun, 1956. | Lower Cretaceous | Chawan Fm., Zhenjian and Fujian Provinces, China |
| Ranulfoichthys dorsonudumAlvarado-Ortega, 2014. | Albian | Tlayúa Fm. Puebla, Mexico |
| Rhombichthys intoccabilisKhalloufi, Zaragüeta-Bagils, and Lelièvre, 2010. | Cenomanian | Ein Yabrud, Palestine |
| Scutatoclupea bachiaiBannikov, 2015. | Cenomanian | Namoura, Lebanon |
| Scutatoclupea applegatei (Alvarado-Ortega and Ovalles-Damián, 2008). | Cenomanian | Sierra Madre Fm., Chiapas, Mexico |
| Scutatuspinosus itapagipensisSilva-Santos and Corrêa, 1985. | Neocomian | Recôncavo Fm., Bahía, Brazil |
| Sorbinichthys elusivoBannikov and Bacchia, 2000. | Cenomanian | Namoura, Lebanon |
| Sorbinichthys africanusMurray and Wilson, 2011. | Cenomanian/Turonian | Akrabou Fm., Agoult, Morocco |
| Thorectichthys rhadinusMurray and Wilson, 2013. | Cenomanian/Turonian | Akrabou Fm., Agoult, Morocco |
| Thorectichthys marocensisMurray and Wilson, 2013. | Cenomanian/Turonian | Akrabou Fm., Agoult, Morocco |
| Triplomystus oligocostatusForey, Yi, Patterson, and Davies, 2003. | Cenomanian | Namoura, Lebanon |
| Triplomystus nooraeForey, Yi, Patterson, and Davies, 2003. | Cenomanian | Namoura, Lebanon |
| Tycheroichthys dunveganensisHay, Cumbaa, Murray, and Plint, 2007. | Cenomanian | Dunvegan Fm., Alberta, Canada. |
Applegate (1996) reported the occurrence of an ellimmichthyiform Diplomystus-like in Tlayúa, which later Chang and Grande (1997, Fig. 7e-f) suggested as an indeterminate species of Ellimmichthys; however, no major progresses were made in relation to these fishes or other basal clupeomorphs recovered in this site. After fifteen years, authors of the present paper take over this challenge. Up to now, Melgarejo-Damián (2011) conducted a comparative study on the body shape of the double armored herrings from Tlayúa, in which she suggested the occurrence of at least four different ellimmichthyiforms in the site. More recently, Alvarado-Ortega (2014) described Ranulfoichthys dorsonudum, a peculiar streamlined basal clupeomorph without the predorsal scute series. When it was described, Ranulfoichthys was considered as a basal clupeomorph, neither ellimmichthyiform nor clupeiform; however, Vernygora et al. (2016, fig. 4) have pointed it out as an ellimmichthyiform.
The aim of the present work is to provide an accurate description of a new ellimmichthyiform species from the Tlayúa quarry, as well as to support its inclusion into the family Paraclupeidae as a new species of the genus ParaclupeaSun, 1956. This record represents an important contribution to the diversity and biogeography of Paraclupea during the Aptian-Cenomanian; this Mexican fossil fish represent the second species of the genus and increases its distribution from eastern Asia (China) to the southeastern region of North America (Mexico); hence, the new species is an interesting element that should be integrated into the future efforts to understand the evolution and biogeographical distribution of the Ellimmichthyiformes and basal clupeomorphs.
MATERIAL AND METHODS
Preparation methods
The cleaning acid technique of Toombs and Rixon (1959) was used to prepare some of the specimens described in this work. Pin vices and needles were used under a binocular microscope to clean remaining limestone patches. As noted in the figure captions, prepared specimens were photographed under long-wave UV (254 nm) light and were also coated with magnesium dust to get black and white photographs of high contrast under white light.
Anatomical abbreviations
Nomenclature and anatomical abbreviations used in this paper and listed in the figure captions follow Grande (1982, 1985), Forey et al. (2003), Di Dario (2004), and Di Dario and de Pinna (2006).
Institutional abbreviations
CME, Colección de Material Extranjero para Comparación, a subcollection of IGM (see below). BMNH P, Paleontological Collection of the British Museum of Natural History, England. DNPM, Departamento Nacional de Produção Mineral, Brazil. IGM, Colección Nacional de Paleontología, Instituto de Geología, Universidad Nacional Autónoma de México. IHNECH, Museo Eliseo Palacios Aguilera, Instituto de Historia Natural y Ecología de Chiapas, México. INAH, Instituto Nacional de Antropología e Historia, Chiapas, México.
Comparative materials
The following specimens were studied for comparison. Armigatus brevissimus (Blainville, 1818): seven specimens of IGM cataloged as CME 874 to CME 880, all from the marine Cenomanian deposits of Heckel, Lebanon. Armigatus namourensisForey, Yi, Patterson, and Davies, 2003: BMNH P.62528 and BMNH P.63146, from Namoura, Lebanon. Diplomystus birdi, Woodward, 1895: BMNH P.86, BMNH P.96, and BMNH P.96a from Cenomanian deposits of Heckel, Lebanon; BMNH P.13878 from Cenomanian deposits of Hajula, Lebanon. Diplomystus dentatusCope, 1877: BMNH P.52501, BMNH P.24790, BMNH P.63596, BMNH P.5218, CME 883, all from Eocene lake deposits of the Green River Shales, Wyoming, USA. Ellimmichthys longicostatus(Cope), 1886: DNPM 551-P; Rio de Pixuma, Candeiras, Bahia, Brazil. Paraclupea-like (Alvarado-Ortega et al., 2008): IHNECH-2332 and IHNECH-2331 (part and counterpart of the same specimen); the El Espinal Quarry, Cintalapa Member of Sierra Madre Formation, Chiapas, Mexico. Scutatuspinosus itapagipensisSilva-Santos and Corrêa, 1985: DNPM 1207-P, 1208-P, 1209-P; Recôncavo Basin, Ilhas Group, Salvador, Bahia, Brazil. Scutatoclupea applegatei (Alvarado-Ortega and Ovalles-Damían, 2008) described as Triplomystus applegatei: INAH 1941 (holotype); the El Espinal Quarry, Cintalapa Member of Sierra Madre Formation, Chiapas, Mexico. Triplomystus nooraeForey, Yi, Patterson, and Davies, 2003: BMNH P.63325 and BMNH P.62517, from the Cenomanian deposits of Namoura, Lebanon.
SYSTEMATIC PALEONTOLOGY
Subdivision Teleostei Müller, 1845
Cohort Clupeocephala Patterson and Rosen, 1977
Superorder Clupeomorpha Greenwood et al., 1966
Order Ellimmichthyiformes Grande, 1982
Family Paraclupeidae Chang and Chou, 1977
Genus ParaclupeaSun, 1956
Paraclupea seilacheri sp. nov.

Figure 3 Paraclupea seilacheri sp. nov.: a) IGM 4717, holotype; b) outline drawing of the same specimen.


Figure 5 Outline drawing of the head of IGM 4720, a paratype of Paraclupea seilacheri sp. nov. (also see Figure 3). Abbreviations: ao, antorbital; ang, anguloarticular; as, abdominal scutes; asph, autosphenotic; br, branchiostegal ray; cl, cleithrum; den, dentary; dsph, dermosphenotic; ect, ectopterygoid; ent, endopterygoid; epo, epioccipital; epn, epineurals; exo, exoccipital; ext, extrascapular; fr, frontal; hm, hyomandibular; iop, interopercle; le, leteral ethmoid; mes, mesethmoid; mp, metapterygoid; mx, maxilla; na, nasal; n6, neural arch of centrum 6; op, opercle, os, orbitosphenoid; pa, parietal; pal, palatine; pcl, postcleithrum; pd, predorsal; pds, predorsal scutes; pf, pre-epiotic fossa; pop, preopercle; ps, parasphenoid; pto, pterotic; ptt, posttemporal; q, quadrate; rar, retroarticular; sc, sclerotic; scl, supracleithrum; smx, supramaxilla; soc, supraoccipital; sop, subopercle; sy, symplectic; v, vomer; numbers indicate the position of centra.
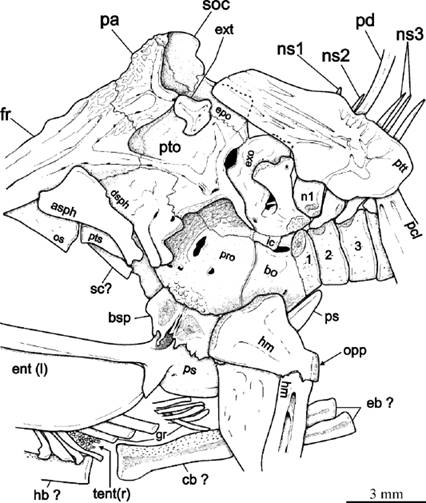
Figure 6 Outline drawing of posterior part of the head of Paraclupea seilacheri sp. nov. preserved in the paratype IGM 4721. Abbreviations: asph, autosphenotic; bo, basioccipital; cb, ceratobranchial; dsph, dermosphenotic; eb, epibranchials; ent (l), left endopterygoid; epo, epioccipital; exo, exoccipital; ext, extrascapular; fr, frontal; gr, gill racker; hb, hypobranchial; hm, hyomandibular; ic, intercalar; ns, neural spine; opp, opercular process of the hyomandibular; os, orbitosphenoid; pa, parietal; pcl, postcleithrum; pd, predorsal; pro, prootic; ps, parasphenoid; pto, pterotic; pts, pterosphenoid; ptt, posttemporal; sc, sclerotic; soc, supraoccipital; tent (r), teeth of the right entopterygoid; numbers indicate the position of centra.

Figure 7 Photo (a) and outline drawing (b) of premaxilla, maxilla and dentary bones of Paraclupea seilacheri sp. nov. preserved in IGM 4720. Abbreviations: den, dentary; pmx, premaxilla; mx, maxilla; smxa, supramaxilla anterior; smxp, supramaxilla posterior; arrows show maxilla and dentary teeth; and (l) and (r) indicate left and right side.

Figure 8 Photo (a) and outline drawing (b) of the ceratohyal bones of Paraclupea seilacheri sp. nov. preserved in IGM 4723. Abbreviations: ac, anterior ceratohyal; bf, beryciform foramen; br, branchiostegal rays; dh, dorsal hypohyal; gha, groove for the afferent hyoidean artery; pc, posterior ceratohyal; vh, ventral hypohyal.

Figure 9 Photo (a) and outline drawing (b) of a closeup of the anterior middle part of the trunk of IGM 4717, holotype of Paraclupea seilacheri sp. nov., showing the abdominal centra, predorsal scutes, and dorsal fin. Abbreviations: cl, cleithrum; dfr, dorsal fin ray; dfs, dorsal fin stay; epl, epipleural; epn, epineural; mpt, middle pterygiophore; pcl, postcleithrum; pd, predorsal; pds, predorsal scute; ptt; posttemporal; r, rib; scl, supracleithrum.
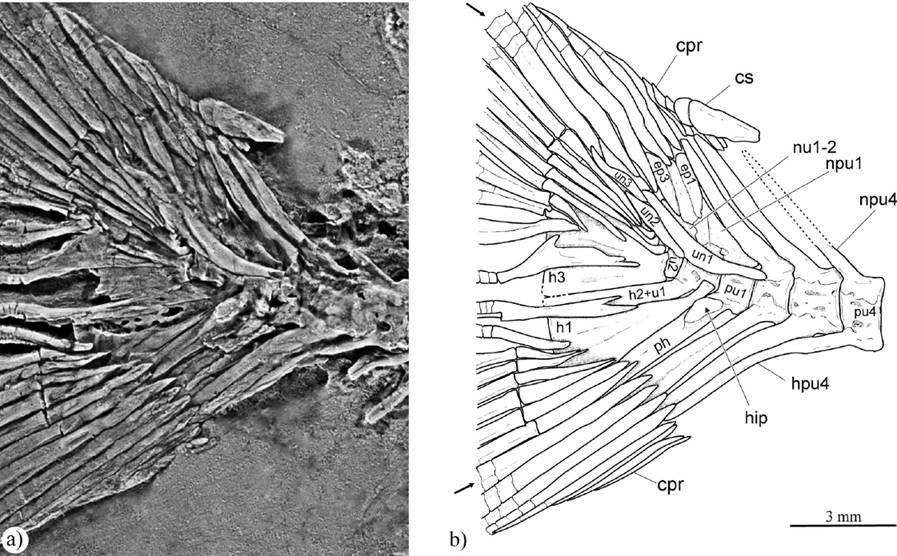
Figure 10 Photo (a) and outline drawing (b) of the caudal fin preserved in IGM 4720, a paratype of Paraclupea seilacheri sp. nov. Abbreviations: cpr, caudal procurrent ray; cs, caudal scute; ep, epural; h, hypural; hpu, haemal arch of preural centrum; npu, neural arch of preural centrum; nu, neural arch of ural centrum; ph; parhypural; pu, preural centrum; u, ural centrum; un, uroneural; arrows show the principal caudal fin rays.
Holotype. IGM 4717, part and counterpart (IGM 4717a and IGM 4717b, respectively) of a complete specimen exposing the left side of the body of 92.6 mm of standard length (SL) (Figure 3), collected in the Tlayúa quarry, stratigraphic level “Zone XIII” and section “IGM-locality” 2772.
Paratypes. These specimens were collected in different stratigraphic levels (= Zones) and sections (= IGM-localities) of the Tlayúa quarry. IGM 4718, complete specimen of 103.6 mm of SL that exposes the right side of the body, from the Zone XIII and IGM-locality 2781. IGM 4719, a complete specimen of 98.2 mm of SL exposing the right side of the body (Figure 4), collected in the Zone XV and IGM-locality 2432. IGM 4720, complete specimen of 92.6 mm of SL exposing the right side of the body (Figures 4, 5, 7, 9, 10), from the Zone XVII and IGM-locality 2432. IGM 4721, complete specimen of 93.2 mm of SL exposing the left side of the body (Figure 6); its stratigraphic level within IGM-locality 3661 is unknown. IGM 4722, complete specimen of at least 118.8 mm of SL exposing the right side of the body, from unknown stratigraphic level of IGM-locality 2828. IGM 4723, complete specimen of 110.4 mm of SL exposing the right side of the body, largely preserved as impression (Figure 8), from the Zone XIII and IGM-locality 2432.
Locality and age. Albian (Early Cretaceous) laminated limestones of the Tlayúa quarry belonging to the Middle Member of the Tlayúa Formation, near Tepexi de Rodríguez, Puebla State, central Mexico (Pantoja-Alor, 1992; Benammi et al., 2006; among others).
Etymology. The specific name honors our colleague, friend, and teacher, the German professor Adolf Seilacher (1925-2014), for his contributions to the study of Konservat-lagerstätten sites.
Diagnosis. Paraclupea species that differs from the type species of the genus in the following meristic characters. This species has 7 predorsal bones (versus 8-9); 16 dorsal fin rays (versus 17-18); 38 to 39 total vertebrae (versus 41); 33 to 34 abdominal scutes including 8 to 9 postpelvic scutes (versus 38 to 43, with probably 10 postpelvic scutes).
Description
General features and proportions
Table 2 summarizes the measurements and body proportions of the specimens described here as Paraclupea seilacheri sp. nov. The overall body shape of this species resembles that of other double armored herrings, as Paraclupea chetungensisSun, 1956, and Ellimmichthys goodi (Eastman, 1912) (see Chang and Grande, 1997, figs. 3, 7). In lateral view, P. seilacheri is a dumpy looking fish with a rounded abdominal border, in which the maximum standard length (SL) known is 118.8 mm and corresponds to specimen IGM 4721, whose total length exceeds 154.4 mm. In this species, the maximum body depth and the head length represent 50.9 and 32.9 % of the SL, respectively. The maximum depth of the trunk is present just before the half body length, in the point where the dorsal fin arises (= predorsal length) and the dorsal body profile forms an obtuse angle, at 49.3% the SL. The depths of the head and caudal peduncle are about two thirds and one third of maximum body depth, respectively. The head is triangular; slightly longer than deep.
Table 2 Measurements and proportions of specimens of Paraclupea seilacheri sp. nov. from the Tlayúa quarry studied here. All percentages are calculated on the basis of the standard length. Abbreviatures: Dorsal-VC depth = distance between the highest point of the dorsal border and the vertebral column. Ventral-VC depth = distance between the lowest point of the ventral border and the vertebral column.
| IGM 4717 | IGM 4718 | IGM 4719 | IGM 4720 | IGM 4721 | IGM 4722 | IGM 4723 | Average | |
|---|---|---|---|---|---|---|---|---|
| Measurements and proportions | ||||||||
| Total length | 122.6 | 135.1 | 114.5+ | 122.6 | - | 154.4? | - | - |
| Standard length | 92.6 | 103.6 | 98.2 | 92.6 | 93.2 | ≈118.8 | 110.4+ | |
| Head length | 31.3 | 34.1 | 30.7 | 31.3 | 30.2 | 37.3 | 37.4 | |
| 34.80% | 32.90% | 31.20% | 34.80% | 32.4 | 31.40% | - | 32.90% | |
| Predorsal length | 44.8 | 50.9 | 48.7 | 44.8 | 48.4 | 58.3 | 58.3 | |
| 48.30% | 49.10% | 49.50% | 48.30% | 51.90% | 49% | - | 49.30% | |
| Dorsal fin length | 17.8 | 19.2 | 18.5 | 17.8 | 15.8 | 23.1 | - | |
| 19.20% | 18.50% | 18.80% | 19.20% | 17% | 19.40% | - | 18.60% | |
| Prepelvic length | 52.2 | 59.3 | 58.6 | 52.2 | 51.4 | 71.7 | 69.6 | |
| 56.30% | 57.20% | 59.60% | 56.30% | 55.1 | 60.30% | - | 57.50% | |
| Preanal length | 72.8 | 77.9 | 72.6 | 72.8 | 71.2 | 96.9 | - | |
| 78.60% | 75.20% | 73.90% | 78.60% | 76.30% | 81.50% | - | 77.30% | |
| Anal fin length | 17.7 | 18.8 | 19.3 | 17.7 | 18.3 | 21.5 | - | |
| 19.10% | 18.10% | 19.60% | 19.10% | 19.60% | 18.10% | - | 18.90% | |
| Body depth | 48.5 | 50.3 | 50.1 | 48.5 | 53.5 | 58.1 | 60.4 | |
| 52.30% | 48.50% | 51.50% | 52.30% | 57.40% | 43.90% | - | 50.90% | |
| Dorsal-VC depth (A) | 18.2 | 17.8 | 19 | 18.2 | 18.9 | 21.2 | 21.6 | |
| 19.60% | 17.10% | 19.30% | 19.60% | 20.2 | 17.80% | - | 18.90% | |
| Ventral-VC depth (B) | 30.3 | 32.5 | 31.1 | 30.3 | 32.4 | 38.3 | 37.7 | |
| 31.40% | 31.30% | 31.60% | 31.40% | 34.70% | 32.2 | - | 32.10% | |
| B/A ratio | 1.66 | 1.82 | 1.63 | 1.66 | 1.82 | 1.8 | 1.74 | 1.73 |
| Accounts | Mode | |||||||
| Total vertebral centra | 38 | 38 | 39 | 38 | 39 | ≈38 | - | 38 |
| Abdominal | 20 | 22 | 21 | 20 | 21 | 20 | - | 20 |
| Caudal+ural | 18 | 16 | 18 | 18 | 18 | ≈18 | - | 18 |
| Predorsal scutes | 18 | 18 | 10+ | 18 | - | 18 | - | 18 |
| Abdominal scutes | 33 | 34 | 33 | 33 | 33 | 33 | - | 33 |
| Prepelvic scutes | 24 | 25 | 25 | 24 | 24 | 24 | - | 25 |
| Postpelvic scutes | 9 | 9 | 8 | 9 | 9 | 9 | - | 9 |
| Dorsal fin rays | ii-16 | ii-16 | ii-14+ | ii-16 | - | ii-16 | - | ii-16 |
| Dorsal pterygiophores | 16 | 16 | 16 | 16 | 16 | 16 | - | 16 |
| Anal fin rays | - | 5+ | 8+ | - | - | - | - | - |
| Anal pterygiphores | 15 | 13 | 17 | 15 | 16 | 17 | - | 17 |
| Predorsal bones | 7 | 8 | 7 | 7 | 7 | 7 | - | 7 |
In this new species, the caudal fin is almost as depth as the trunk. Both caudal lobes are triangular and deeply forked. Unpaired fins are triangular, relatively smaller, and not opposed to each other. On the back, the dorsal fin represents about 18.6 % of the SL, and it is located along the 49.3 and 67.9 % of the SL. In contrast, the anal fin occupies about 18.9% of the SL on the ventral body edge, and it is located at the beginning of the posterior half of the body, along the 77.9 and 96.2 % of the SL.
The paired fins of Paraclupea seilacheri sp. nov. are relatively short. The pectoral fin is located behind the pectoral girdle, slightly below the midpoint between the vertebral column and ventral body edge. The larger pectoral rays cover the anterior half of the distance between the bases of the pectoral and pelvic fins. The pelvic fin is placed below the posterior half of the dorsal fin at 57.5 % of the SL; its larger rays are two thirds of the larger pectoral rays.
Skull
The head is laterally preserved in all specimens of Paraclupea seilacheri sp. nov. studied here (Figures 3 - 5). The head is triangular, about 1.5 times longer than deep; its otico-occipital, ocular, and ethmoid sections are about of the same length.
The frontals roof large part of the skull; they are flat and roughly triangular, narrow anteriorly and broaden posteriorly (Figure 5). In their posterior portion, these bones suture with the parietals, pterotic and autosphenotic. Frontals are joined together throughout their length and there is no fontanelle between them. Frontals are smooth bones except for a small posterior area, near the parietals, where they are strongly ornamented with irregular ridges. The supraorbital sensory canal runs along the frontals enclosed by bone.
Parietals are small sub-square bones that meet each other along the middle line of the skull and separate the supraoccipital and posterior end of frontal bones. There is no temporal foramen. Parietals are strongly ornamented with fine ankylosed sigmoid ridges forming a reticular pattern. The supratemporal commissure runs, enclosed by bone, along side the posterior edge of the parietals. The supraoccipital is a small bone in the posterior margin of the skull that carries part of the supratemporal commissure; it is restricted to the tip of a stout supraoccipital crest (Figures 5, 6). Beyond the parietal, the posterolateral end of the supratemporal canal communicates with the extrascapular canal.
The pterotic is a rectangular bone in the lateral skull wall; it meets the parietal and frontal medially, the epiotic and epioccipital posteriorly, and the large triangular dermosphenotic anteriorly (Figure 5). A small thin pre-epiotic fossa is present between the pterotic and epiotic. There is no evidence of the recessus lateralis on lateral skull bones. The hyomandibular fossa is roofed by the pterotic and dermosphenotic bones (Figure 5). The epioccipital is short, massive and restricted to dorsoposterior part of the skull. Bones in the lateroposterior and otic skull areas are obscured.
The orbital section of the parasphenoid is an edentulous rod like ossification; at the end of the orbit, it carries a small basipterygoid process that is projected anteroventrally (Figure 5). In IGM 4721, the postorbital part of the parasphenoid is partially exposed (Figure 6); here, this bone is extended along the entire ventral skull edge. Above, the prootic is exposed showing a medial irregular surface with appearance of strawberry that here is interpreted as a superficial expression of the bulla as previously interpreted in other fossil clupeomorphs by Forey (1975) and Grande (1985, p. 253). This specimen also shows the triple joint surface on the anterior face of the first vertebral centrum, which includes a large ventral surface to joint with the basioccipital and two small dorsal surfaces to joint the couple of intercalar bones. The first vertebral centrum also reveals a dorsal surface to joint with the respective halves of the neural arch.
The mesethmoid is a massive bone with a rounded massive edge and a flat rear overlapped by the frontals. On each side, this bone shows a lateral process ending in a facet for the respective palatine head. The lateral ethmoid is a thin roughly rectangular bone that encloses the posterior ends of the nasal capsule, and is located between the frontal and parasphenoid. The nasal is a small and posteriorly tapered tubular bone placed over the ethmoid area. Only small parts of the pterosphenoid and orbitosphenoid bones are exposed in some specimens studied here (Figures 5, 6).
Upper jaw
Upper jaw consists of the premaxilla, maxilla and two supramaxillae. The premaxilla is a sickle-shaped toothless bone, about one-fourth the maxillar length. The maxilla has two sections forming an obtuse angle. The anterior toothless maxilla section is a smooth and rod-like structure tilted upward; in contrast, the toothed maxilla section is a flat, almost straight, and horizontally lamina, slightly longer than the last, in which the external surface is ornamented with scarce reticular grooves. A single row of small, uniformly spaced and sized, conical teeth is present along the ventral edge of the last maxillar section. Two drop-shaped supramaxillae are present overlapping the dorsal edge of the toothed maxillar section; the anterior is slightly smaller and rounded than the posterior one that has a long and sharp dorsal process. External surfaces of supramaxillae also are ornamented with scarce reticular shallow grooves (Figures 5, 7).
Lower jaw
This jaw consists of dentary, anguloarticular, and retroarticular bones. The lower jaw shows a shallow and short ventral postarticular process. The dentary is triangular with a shallow symphysis, a high coronoid process, and a sinuous alveolar border relative short and tilted upward. A row of small conical teeth is present along the alveolar border of this bone; the shape, size, and distribution of these teeth resemble those of the maxilla (Figures 5, 7).
The anguloarticular occupies about two thirds of the lower jaw length; it forms the bulk of the postarticular process and the articular facet for the quadrate. The small retroarticular bone occupies the ventral end of the postarticular process. The mandibular sensory canal runs alongside the ventral border of lower jaw; this sensory canal opens in six or seven pores that are present from the postarticular process to dentary symphysis (Figure 5).
Circumorbital bones
These bones are flimsy and flat. Since these are strongly fractured in all the specimens studied; their number and shapes are only partially known. The circumorbital series includes the antorbital, dermosphenotic, and probably four infraorbitals. Among these bones, the first four infraorbitals are located below the orbit covering part of the ectopterygoid and endopterygoid. The infraorbital sensory canal runs near the dorsal edge of the infraorbital bones (Figure 5).
Suspensorium
The hyomandibular is a hatchet-like bone with a single stout head and a narrow long shaft ventrally projected. This bone shows a well developed opercular process located at the base of the back of its head. The quadrate bone is triangular; its articular condyle is inclined forward and lies beneath the anterior half of the orbit. The spiny posterior process of the quadrate is as high as this bone. The symplectic is about twice longer than the posterior process of the quadrate (Figures 5, 6).
The metapterygoid is a roughly square bone that bears numerous rounded and regular sized teeth on the anterior a middle part of the lingual surface. The ectopterygoid is a smooth, narrow, and curved bone, in which the vertical and horizontal limbs form an angle a little greater than 90°. The ectopterygoid meets the ventral border of the endopterygoid and the anterior border of the quadrate. The endopterygoid is a long and semi-ovoid flat bone, wider than the ectopterygoid; its entire lingual surface bears numerous rounded and regular sized teeth, like those that cover part of the metapterygoid. The palatine is as long bone with a small stout head that dorsally articulates with the lateral process of the mesethmoid and ventrally with the maxillar palatine facet.
Opercular bones
The preopercle shows an inverted L shape, in which the length of the horizontal anterior limb is about one half the height of its vertical dorsal limb. The preopercular sensory canal runs close to the inner edges of this bone, it presents three to four branches on the horizontal limb and only one in the vertical limb (Figure 5).
The opercle is a kidney-shaped flat bone, higher than long, with the anterior edge straight and thick. Its surface is covered with fine straight parallel grooves that radiate from the area of its hyomandibular facet. The subopercle is a semicircular bone with a straight dorsal border, a gently curved ventral edge, and a small upward spiny anterior process. The interopercle, located below the preopercular horizontal limb, is a smooth, flat, and spathiform bone (Figure 5).
Branchiostegal rays and branchial arch
Bones of the branchial arch are partially or totally covered in the specimens studied (Figures 5, 8). The anterior ceratohyal is rectangular, about twice longer than high; this bone is pierced by a large beryciform foramen and has a horizontal groove for the afferent hyoidean artery. The center of the semicircular posterior ceratohyal is also pierced by a small foramen. In front of the anterior ceratohyal, the dorsal and ventral hypohyals have similar size and are strongly attached to each other. There are 10 flat and blade-like branchiostegal rays that progressively are longer and wider. In IGM 4722, the urohyal is elongated and has a small anterior head.
Axial skeleton
In this species, the vertebral column is composed of 38 to 39 centra, including 20 to 22 abdominal, 16 to 18 preural, and 2 ural (Figures 3, 9; Table 2). In the preural region, the vertebral centra are rectangular, slightly higher than long, sculpted with longitudinal ridges and pits. The complete and well articulated specimens studied here seem to have the centra fused with the respective neural and haemal arches; however, IGM 4718, where some centra are disarticulated and scattered, reveals that at least the most anterior abdominal centra are not fused with the neural arches. In the most anterior abdominal centra the apex of the neural spines is bifid. There are 20 pairs of long curved ribs that almost embrace the entire abdominal cavity. The heads of the most anterior ribs are stout and joint with the cavities in the ventrolateral surface of the respective abdominal centra. The last twelve abdominal centra show straight and downward parapophyses; these tend to be longer in the posterior vertebrae. The proximal ends of posterior ribs joint with these parapophyses except the last three, which are floating ribs that do not reach the parapophyses.
Long epineurals are associated with the abdominal centra and the anterior half of preural centra. In the anterior abdominal centra, the epineurals are three centra long, thin, and curved bones, projected laterally upward and backward from the bases of the neural arches. Beyond, these bones are progressively shorter and located on the neural spines, away of the neural arches. In addition, in the preural centra each epineural develops a thin anterior ascending process that gives the Y-shape to this bone.
There are seven epipleurals, thin and slightly curved, associated with the last nine abdominal and first three or four preural centra. These bones rest horizontally below the centra and almost have the length of two of them. In the abdominal area, the epipleurals meet the parapophyses and ribs while those of the preural area do this with the haemal spines.
There are seven predorsal bones (named as supraneurals by Forey (1973)) between the occiput and the dorsal fin base, below the predorsal scute series. These stick-like bones increase in size in anteroposterior order, show ventral tips slightly curved forward, and have small anterior and posterior laminar wings or expansions.
Pectoral girdle and fins
In the top of the pectoral girdle, the posttemporal is a flat, ovoid, long bone with the dorsal process hypertrophied, rectangular and wide; the surface of this bone is excavated by numerous branches of the posttemporal sensory canal. Below, the supracleithrum is a flat an ovoid bone that is slightly curved and long; the cleithrum is S-shaped with the posterior vertical limb being slightly higher than the anterior vertical limb. The pectoral fin is located below the corner between the middle horizontal section and the posterior vertical limb of the cleithrum. Two long, ovate, and flat postcleithra are present behind the posterior vertical limb of the cleithrum. Other bones of the pelvic girdle are obscured in all the specimens studied here (Figure 5).
The pectoral fin consists of 16 branched and segmented rays. The longest of these rays equals the length of six abdominal centra. Although it is difficult to recognize the endochondral elements of this girdle, at least four proximal radials articulate with the pectoral fin rays.
Pelvic girdle and fins
The pelvic fin is triangular, short, and opposed to the posterior half of the dorsal fin. This fin is located just at the beginning of the posterior half of the body, at 58 %of the SL, beneath abdominal centrum 19. At least six pelvic fin rays are present in this species, problably these are a few more because the smaller rays are generally covered in all the specimens studied here. The length of the larger pelvic rays equals the length of three abdominal vertebrae. The pelvic bones are unknown because the abdominal scutes cover them (Figures 3, 4).
Dorsal fin
This triangular fin is short and rises just at the end of the first half of the SL. The length of the dorsal fin base represents 19 % of the SL and is extended above the last ten abdominal vertebrae (12 to 22). This fin consists of two small unsegmented and unbranched rays and 17 branched and segmented rays. The fourth branched ray is the longest in this fin; front and behind it other rays are progresively shorter (Figure 9).
There is a series of 16 proximal pterygiophores supporting the dorsal fin rays; these are rod-like elongate structures with anterior and posterior laminar expansions or wings. The first of these elements is the longest and the only bifurcated in the series and joints with the first two small, unbranched, and unsegmented dorsal fin rays (Figure 9). The posterior proximal pterygiophores become progressively shorter and each of them joints only one ray. Some middle pterygiophores are present between the dorsal heads of the proximal pterygiophores; these are stout rectangular bones. Small rounded distal pterygiophores are also present but it is hard to determine if these are present along the entire series. At end of this fin, there is a small and expanded dorsal fin stay in horizontal position.
Anal fin
The anal fin is short and located far at the back of the trunk, behind the dorsal fin level (Figures 3, 4). The anal fin length is about 17 % of the SL; it rises in the last fifth of the body (at 78 % the SL), below the preural centra 4 to 11. Unfortunately, the rays of this fin are not well preserved in any of the referred specimens. Although the anal rays are unknown; these could be one or two units more than those 16 pterygiophores or proximal radials that internally support this fin. These internal rod-like structures are slightly curved and of decreasing sizes. The distal ends of the pterygiophores almost reach the tip of the nearby haemal spines. Small distal and middle pterygiophores or radials are identifiable in the base of the anal fin.
Caudal fin
The haemal and neural spines of the last four preural and two autogenous ural centra support the elements of dorsal and ventral caudal fin lobes. The neural and haemal spines of the three posterior preural centra are wider than the anterior ones (Figure 10). The caudal fin is deeply forked. The rays that constitute the dorsal and ventral caudal fin lobes are one unbranched, 10 principals, and 6 procurrents, as well as one unbranched, 9 principals, and 7 procurrents, respectively; additionally, there are two and one caudal scutes preceeding these caudal lobes in the same order. The anterior ends of all principal rays are pointed but in the two most internal caudal principal rays, in which ends are expanded and bifurcated.
The stout parhypural and the preural centrum 1 are fused. The first preural centra and ural 1 are of the same size while the ural 2 is smaller. It seems that an expanded neural arch is present in both ural centra. On the first preural centrum, the neural arch and its respective spine form a broad leaf-like structure, so expanded that it fills the space between the spine of the preural 2, the neural arches of both urals, and the epurals. The hypural 1 is an autogenous fan shaped or triangular structure that joints with ural 1. Above hypural 1, six spathiform hypurals are present, placed side by side; all of them joint the ural 2 and seem to be autogenous too but the hypural 2 that is fused with ural 1. There is no a caudal diatema (= hiatus or free space) between the hypurals 2 and 3.
There are three wide epurals located behind the spine of preural 1. There is no evidence of urodermals. There is a series of three uroneurals. Uroneural 1 is a long and curved structure in which the anterior end is on the dorsal region of preural centrum 1. Uroneural 2 is almost one half the length of the first, its distal end reaches the distal end of uroneral 1, and it covers a small part of ural 2. Uroneural 3 is the shortest of the series and is located between the distal ends of uroneurals 1 and 2 and the first dorsal principal ray.
Scutes and scales
The entire trunk is covered with ovate cycloid scales and two scute series located on the predorsal and the abdominal edges. The scales are flat, ovate, and deeper than long; they are intensely ornamented with concentric circuli. Unfortunately, it is not possible to recognize the number of scale rows that cover the body trunk of this species.
The predorsal scute series occupies the entire predorsal edge of the body, from the occiput to dorsal fin base. This series includes 18 scutes that increase in size from front to back (Figure 9). Although the firsts ones of these scutes are rather ovoid, slightly longer than wide; those behind them become wider. In dorsal view, those predorsal scutes located in the middle of the series are arrowhead shaped because their middle longitudinal keels are underdeveloped and do not project backward; in contrast, the five posterior predorsal scutes are rectangular, about 3-4 times wider than long, forming lateral wings projected downward from each side of a strong middle longitudinal keel that is projected backward forming a short spine. All these scutes are intensely ornamented with stout and sinuous ridges projected from the middle longitudinal keel.
The abdominal or ventral scute series is located along the ventral edge of the trunk, between the pectoral fin and the anus. This consists of 34 scutes, including 24 prepelvic scutes located between the pectoral and pelvic fins, one pelvic scute located below the pectoral fin, and nine postpelvic scutes located behind (Figure 3). These scutes are four pointed-star shaped, in which the lateral limbs are wide long spatula-like structures, upward projected whereas the anterior and posterior limbs form thick and short keels. The lateral wings of those scutes located in the middle of the abdominal series are so long that cover the ventral half of the abdominal area. In contrast, the scutes located in front and back of this series have smaller lateral wings.
DISCUSSION
The superorder Clupeomorpha is well supported by the occurrence of two synapomorphies (Patterson and Rosen, 1977; Grande, 1985; Arratia, 1997). These are the otophysic connection (a diverticulum of the swim bladder that penetrates the exoccipital and prootic, in which it forms the ossified bulla that superficially shows the appearance of strawberry (Forey, 1975), as well as the supratemporal commissural sensory canal passing through parietals, or through parietals and supraoccipital. Grande (1985) also indicates the presence of abdominal scutes as an additional clupeomorph synapomorphy. Paraclupea seilacheri sp. nov. shows all these diagnostic characters supporting its undeniable inclusion into the suborder Clupeomorpha (Figures 5, 6). The presence of the prootic bulla in this new species, revealed by a small strawberry surface in the prootic, confirms the occurrence of this feature among ellimmichthyiforms firstly suspected in Diplomystus dentatus (Grande, 1985, p. 253).
Since the establishment of the order Ellimmichthyiformes as a natural group by Grande (1982, 1985), multiple efforts have been performed to include an increasing number of species into this order. This has led to the proposal of different and divergent phylogenetic hypotheses, some of them more comprehensive than others (Chang and Maisey, 2003; Zaragüeta-Bagils, 2004; Forey, 2004; Alvarado-Ortega et al., 2008; Murray and Wilson, 2013; Vernygora and Murray, 2016; Vernygora et al., 2016; Figuereido and Ribeiro, 2016, 2017; among others). Among these, the hypotheses published by Vernygora and Murray (2016) and Marramà and Carnevale (2016), the most inclusive and elaborate, show similar topologies; therefore, the first of these is considered to discuss the affinities of the new species described in this paper.
Paraclupea seilacheri sp. nov. displays most of the characters that support the naturalness of the order Ellimmichthyiformes, the family Paraclupeidae, as well as the unnamed clades Scutatuspinosus-Triplomystus, Ellimma-Triplomystus, Tycheroichthys-Triplomystus, Tycheroichthys-Paraclupea, and the genus Paraclupea, as they were defined in the phylogenetic hypothesis recently published by Vernygora and Murray (2016, fig. 4, suppl. data) (Figure 1). Therefore, undoubtedly this new Mexican fossil fish species is located as a new member of all these clades.
In the hypothesis of Vernygora and Murray (2016) (Figure 1), one of the most recently published, the naturalness of the Ellimmichthyiformes is poorly supported in an exclusive set of homoplasies. These characters include the presence of parietals in contact to each other along the midline of the skull, a beryciform foramen piercing the anterior ceratohyal, three epurals in the caudal skeleton, and a series of predorsal scutes. Among teleosts, the first three of these characters are plesiomorphies and the last one is a homoplasy among clupeomorphs (e.g., Patterson and Rosen, 1977; Grande, 1985; Arratia, 1997). The inclusion of Paraclupea seilacheri sp. nov. in the order Ellimmichthyiformes is well supported because this species has these characteristics.
Paraclupea seilacheri sp. nov. shares those five characters that support both unnamed ellimmichthyiform subgroups in the hypothesis of Vernygora and Murray (2016), Ellimmichthyiformes less Sorbinichthys and Ellimmichthyiformes less [Sorbinichthys and Diplomystus] (Figure 1). These include the cleithrum S-like shaped; the lack of interhypural diastema because the third hypural is expanded posteriorly leaving no gap between this and the second hypural; 22 to 30 abdominal scutes; and presence of caudal scutes and at least 6 to 14 predorsal scutes.
Besides, Paraclupea seilacheri sp. nov. displays the synapomorphies that support the family Paraclupeidae in the hypothesis of Vernygora and Murray (2016) (Figure 1). These are the presence of scutes of irregular size along the predorsal series (their sizes increase in anteroposterior order) and the postpelvic abdominal scutes have a strong ventral spine. Additionally, P. seilacheri has predorsal scutes ornamented with radiating ridges. This character represents the single synapomorphy of the unnamed clade Scutatuspinosus—Triplomystus (= Paraclupeidae less [Kwangoclupea and Thorecthychthys]).
According Vernygora and Murray (2016) (Figure 2), the paraclupeid clade Scutatuspinosus-Triplomystus includes two subgroups, Scutatuspinosus—Rhombichthys and Ellimma—Triplomystus. In Paraclupea seilacheri sp. nov., the posterior scutes of the predorsal series are subrectangular (wider than long) and its abdominal scutes are spatula-like (their wings reach most of the height of the adjacent scutes). The inclusion of P. seilacheri into the Ellimma—Triplomystus clade of the hypothesis of Vernygora and Murray (2016) (Figure 2) is undeniable because these characters are synapomorphies of such subgroup. Additionally, in P. seilacheri, the anterior predorsals scutes also are subrectangular shaped (broader than long) as it occurs in other members of the Ellimma—Triplomystus clade; however, this new species does not have a diastema present between second and third hypural, which is a homoplasy of this clade.
Among paraclupeids, the Tycheroichthys—Triplomystus group in the hypothesis of Vernygora and Murray (2016) is well characterized because the dorsal process of posttemporal bone is wider or hypertrophied (Figure 1). In the present study, Paraclupea seilacheri sp. nov. is also identified as part of this clade because its posttemporal shows the same condition. Moreover, in the same hypothesis, the binomial clade Tycheroichthys—Paraclupea shares three characters; the second uroneural is relatively short and its distal end does not reach the end of the first uroneural; number of predorsal scutes ranges between 16 and 19; and there are more than 32 abdominal scutes. Given that Paraclupea seilacheri sp. nov. has all these characters, it is also nested into this group. Contrary to Tycheroichthys dunveganensis and Paraclupea chetungensis, whose epurals are located far from the spine of preural 2, leaving an open space; the epurals of Paraclupea seilacheri sp. nov. are so wide that there is no open space behind the neural spine of preural 2. Vernygora and Murray (2016) considered the expanded shape of epurals as a primitive condition; therefore, the expanded epurals found in P. seilacheri represent a regressive character.
The morphological comparison between Paraclupea seilacheri sp. nov., Paraclupea chetungensis, and Tycheroichthys dunveganensis reveals the greater number of common characters and closer meristic features sheared by the first two species (Table 3). These observations support the inclusion of P. seilacheri into the genus Paraclupea rather than in Tycheroichthys. Both species of Paraclupea are relatively shallow diamond-shaped fishes with large heads, in which the body depth does not reach 50% of the SL (in these species such proportions are 49.8% and 43-48%, respectively) and the head length represents near 30-35% of SL; in contrast, Tycheroichthys dunveganensis has an extremely high body and short head, in which the body depth is about 90% and the head length is just 24% of SL (also see Chang and Grande, 1997; and Hay et al., 2007). The numbers of anal and dorsal fin rays of both Paraclupea species are less than those present in Tycheroichthys (P. seilacheri has 13-17 anal plus 16 dorsal rays; closely, P. chetungensis shows 17-18 anal plus 14-15 dorsal rays. Contrary, T. dunveganensis shows divergent and greater amounts, 23 anal plus 19 dorsal rays). Both Paraclupea species have 18 predorsal scutes while T. dunveganensis has only 16. The amounts of total vertebrae and abdominal scutes form small clusters; in which P. seilacheri sp. nov. distinctively owns the lowest values of both characters, 38-39 total vertebrae and 33-34 abdominal scutes, which contrast with those 40 vertebrae and 36 abdominal scutes recorded in T. dunveganensis, and 41 vertebrae and 38 to 43 abdominal scutes of P. chetungensis.
Table 3 Comparative meristic data of close relatives of Paraclupea seilacheri sp. nov., Paraclupea chetungensis and Tycheroichthys dunveganensis (based on Chang and Grande, 1997, text and figs. 2 and 3; and Hay et al., 2007).
| Species | Paraclupea chetungensis | Paraclupea seilacheri sp. nov. | Tycheroichthys dunveganensis |
|---|---|---|---|
| Anal fin rays | 17-18 | 13-17 | 23 |
| Dorsal fin rays | 14-15 | 16 | 19 |
| Predorsal bones | 8-9 | 7 | 7 |
| Total vertebrae (abdominal+preural+ural) | 41 (24+15+2) | 38-39 (20-22+14-16+2) | 40 (23+15+2) |
| Predorsal scutes | 18 | 18 | 16 |
| Abdominal scutes | 38-43 | 33-34 | 36 |
| Postpelvic scutes | ≈10? | 8-9 | 10 |
| Body height as SL% | 49.8 | 43-48 | 90 |
| Head length as SL% | ≈30 | ≈31-35 | ≈24 |
Nowadays, it is desirable the confirmation of two characters of Paraclupea chetungensis vaguely reported and illustrated that could be important to understand the morphological diversity of the binomial clade Paraclupea-Tycheroichthys in the hypothesis of Vernygora and Murray (2016) (Figure 2).Chang and Grande (1997, table 1 and figs. 2 and 3) reported the presence of 8 to 9 predorsal bones in P. chetungensis; however, their illustrations do not allow the recognition of these numbers. Furthermore, Chang and Grande (1997) did not report the number of postpelvic scutes in P. chetungensis; however, based on their illustration, these could be around 10 as it occurs in T. dunveganensis. In this framework, P. seilacheri sp. nov. and T. dunveganensis share the presence of only seven predorsal bones, and the presence of only 8 to 9 postpelvic scutes may represent a diagnostic distinctive character of P. seilacheri.
The present discovery reveals that Paraclupea had a wide distribution in the northern hemisphere, comprising two remote continental regions, located on both sides of the Paleopacific Ocean. The presence of this genus in coastal deposits of China and Mexico can only be explained by its swimming capacities to cross along the middle of the Paleopacific Ocean, or by its peripheral dispersion following the epicontinental seas around the Cretaceous lands and the possible existence of some intercontinental connections among Asia and North America. Despite the first of these possible biogeographical scenarios is unlikely, since the rounded body of Paraclupea suggests that these fishes had little chance of being good offshore swimmers; the fossil record of these fish is so scarce that the second scenario cannot be favored now. Today, any biogeographical hypothesis involving Paraclupea is in stalemate due to the lack of additional paleontological evidences.
CONCLUSIONS
Recent studies support a spectacular advance in the understanding of the taxonomic diversity, as well as phylogenetic and biogeographic relationships, of the early clupeomorphs and ellimmichthyiforms; however, these tasks are still far from being concluded. At least in Mexico, a major effort is still required to describe a significant number of these fishes recently recovered in different Cretaceous sites along the country, including the Albian deposits of Tlayúa, in Puebla; the Albian-Cenomanian marine sediments of the Muhi quarry, Hidalgo; and two Cenomanian outcrops, El Chango quarry within the State of Chiapas and the Huehuetla site into Puebla (Pérez-Sánchez and González-Rodríguez, 2010; Alvarado-Ortega, pers. observ.). It is desirable that after the description of these Mexican clupeomorphs, a comprehensive phylogenetic analysis should be carried out to include them together with other basal clupeomorphs.
This article met the goal of describing a new species, P. seilacheri, from the Albian limestone deposits of Tlayúa quarry, Puebla, central Mexico. Also, here we display the supporting characters (e.g., body proportions, occurrence of 18 predorsal scutes) to include this Early Cretaceous species into the genus Paraclupea, previously known only for P. chetungensis, collected in the Chawan Formation deposits along Zhenjian and Fujijan Provinces, China (Sun, 1956; Chang and Grande, 1997). Paraclupea seilacheri clearly differs from P. chetungensis and its close relative, Tycheroichthys dunveganensis, known for a single specimen from Canada (Table 3). Hence, P. seilacheri is the second representative of Paraclupea so far known and the first known in America. Finally, if Scutatoclupea applegatei is considered, P. seilacheri is the second paraclupeid species discovered in Mexico and the third of North American. This finding also indicates the wide distribution of Paraclupea, from China to Mexico.

