










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkMadera y bosques
versión On-line ISSN 2448-7597versión impresa ISSN 1405-0471
Madera bosques vol.21 no.3 Xalapa sep./dic. 2015
Artículos científicos
Estructura y diversidad arbórea en sistemas agroforestales de café en la Sierra de Atoyac, Veracruz
Structure and diversity of arboreal vegetation in coffee agroforestry systems in Sierra de Atoyac, Veracruz
Luis E. García Mayoral1*, Juan I. Valdez Hernández1, Mario Luna Cavazos2 y Rosalío López Morgado3
1 Programa Forestal. Colegio de Postgraduados, campus Montecillo. Estado de México, México. *Autor para correspondencia. garcia.luis@colpos.mx
2 Programa de Botánica. Colegio de Postgraduados, campus Montecillo. Estado de México, México.
3 Centro de Investigación Regional Golfo Centro. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Veracruz, México.
Manuscrito recibido el 19 de mayo de 2014.
Aceptado el 16 de junio de 2015.
Resumen
Se estudió la estructura y diversidad arbórea en sistemas agroforestales de café (SAF-Café): rusticano (RU), policultivo complejo (PC) y policultivo simple (PS), así como en una selva mediana subperennifiolia (SM) de la Sierra de Atoyac, Veracruz, estableciéndose tres unidades de muestreo (UM) por cada franja altitudinal (600-900 y 900-1200) m snm en cada sistema y selva mencionados, para un total de 24 UM de 600 m2 (20 m x 30 m) cada una. Caracterizándose cada sistema mediante la distribución diamétrica y de alturas de sus individuos así como con el cálculo de índices de valor de importancia (IVI), de valor forestal (IVF), de diversidad (Shannon: H\ α Fisher) y de semejanza florística (Sorensen: Si). Para analizar patrones de distribución de especies, se aplicaron técnicas de clasificación (agrupamiento: β flex) y ordenación (correspondencia canónica: CCA) de la vegetación. Fueron determinadas 79 especies arbóreas, la riqueza por sistema fue: SM=65; PC=61; RU=32; PS=18. Cordia alliodora (Ruiz & Pav.) Cham. resultó ser la especie más importante estructuralmente en los SAF-café, Bursera simaruba (L.) Sarg. y Myriocarpa longipes Liebm. en la SM. La diversidad fue significativamente mayor en PC (H'=3.56) y SM (H'=3.58) sin presentar diferencias significativas (p < 0.05) entre estos. La semejanza florística mayor fue entre SM y PC (Si =0.69). Asimismo la altitud fue el factor ambiental que más influyó en la composición florística de los sistemas.
Palabras clave: clasificación, índices de diversidad, ordenación, semejanza florística, valor de importancia, valor forestal.
Abstract
Structure and tree diversity was studied in coffee agroforestry systems (SAF-café): rustic (RU), polyculture complex (PC), simple polyculture (PS), and as semi-evergreen forest (SM) from the Sierra de Atoyac, Veracruz, establishing three sampling units (UM) per altitudinal band (600-900 and 900-1200) m on each system and forest above mentioned, for a total 24 UM of 600 m2 (20 m x 30 m) each one. Each system was characterized by the diameter and height distribution of individuals as well as the calculated importance value (IVI), forest value (IVF), diversity (Shannon H ', α Fisher) and floristic similarity (Sorensen: Si) índices. To analyze patterns of species distribution of vegetation classification (cluster analysis: β flex) and ordination (canonical correspondence analysis: CCA) techniques were applied. 79 tree species were determined, richness per system was: SM= 65; PC= 61; RU= 32; PS= 18. Cordia alliodora (Ruiz & Pav.) Cham. was the most important structurally species in the SAF-café coffee and Myriocarpa longipes Liebm., and Bursera simaruba (L.) Sarg. in the SM. The diversity was significantly higher in PC (H '= 3.56) and SM (H '= 3.58) without significant differences (p < 0.05) between them. The highest floristic similarity was between SM and PC (Si = 0.69). Also the altitude was the most influential environmental factor in the floristic composition of the systems.
Keywords: classification, diversity indices, ordination, florístic similarity, importance value, forest value.
Introducción
Las características físicas, climáticas, orográficas y geológicas que presenta el estado de Veracruz ha generado una riqueza de plantas vasculares calculada entre 7700 (Ramamoorthy et al., 1993) y 9136 especies (Vázquez-Torres, 1998), lo que representa alrededor de 27.7% del total publicado para el país (Llorente-Bousquets y Ocegueda, 2008). El bosque tropical perennifolio (sensu Rzedowski, 1978) es el tipo de vegetación que alberga más especies de plantas en el estado, alrededor de 2230 especies; de las cuales 160 están bajo alguna categoría de riesgo (54 son árboles) de acuerdo con la Norma Oficial Mexicana NOM-059-SEMARNAT-2001 (Castillo-Campos et al., 2011).
Debido a que los ecosistemas primarios se han sustituido por actividades agropecuarias, en el estado de Veracruz, la vegetación prístina ha mermado su superficie original en más de 85% (Castillo-Campos et al., 2011). Esta situación ha ocasionado que Veracruz sea la segunda entidad, con mayor cambio de uso del suelo, en una magnitud de 77% de su superficie (SEMARNAT, 2013). Sin embargo aún quedan áreas que mantienen masas de vegetación forestal como la Sierra de Atoyac, considerándose a esta dentro de las áreas prioritarias para la conservación de la biodiversidad (hotspot) en el estado. No obstante debido a la expansión agrícola, ganadera y urbana, así como la explotación forestal y extracción de especies (Ellis et al., 2011), la masa forestal está siendo disminuida a un grado acelerado, viéndose amortiguada esta situación por la presencia de plantaciones de café bajo sistemas agroforestales (Villavicencio y Valdez, 2003).
Las plantaciones de café bajo sombra son importantes en la conservación de la diversidad biológica, debido a que ciertos sistemas productivos conservan en gran medida la estructura y funcionamiento de los bosques nativos que reemplazaron (Perfecto et al., 1996; Moguel y Toledo 2004; Schroth et al., 2004); es decir, desempeñan un importante papel como refugio para la vida silvestre (Richter et al., 2007; Toledo y Moguel, 2012), propician la infiltración de agua y conservación del suelo (Beer et al., 1998; Soto-Pinto et al., 2001), favorecen la captura de carbono (Pineda-López et al., 2005; Roncal-García et al., 2008) y proporcionan otros servicios ambientales que aún no han sido valorados (Bishop y Landell-Mills, 2003; Manson et al., 2008). Además de ofrecer diferentes productos que contribuyen en la economía de los productores (Moguel y Toledo, 1999).
Así, el estudio de los sistemas agroforestales de café (SAF-Café) ha ganado importancia en los últimos años (Manson et al., 2008), vistos estos como una alternativa que contribuye a disminuir la degradación de los recursos base; agua, suelo y biodiversidad, en las regiones cafetaleras del país (López et al., 2013). No obstante, son escasos los estudios que describen las características estructurales y de la diversidad de especies arbóreas en cafetales (Soto-Pinto et al., 2001; Méndez et al., 2007), ya que frecuentemente se analizan de manera indirecta o en el mejor de los casos de manera general.
Las plantaciones de café bajo sombra en México son descritas, en su estructura arbórea, como un agroecosistema complejo (Soto-Pinto et al., 2001; Martínez et al., 2007). En México, se estima que el café se cultiva por lo menos en dos terceras partes bajo los llamados sistemas sombreados (Moguel y Toledo, 2004), en un gradiente amplio de modalidades que van desde los sistemas tradicionales con árboles de la vegetación original y sombra diversificada (rusticano y de policultivo), pasando por un sistema de policultivo comercial con árboles escasos y especies introducidas, hasta los sistemas especializados: uno con sombra monoespecífica y otro sin sombra (Perfecto et al., 1996; Beer et al., 1998; Toledo y Moguel, 2012).
La caracterización de estos sistemas mediante el uso de índices que midan elementos de su diversidad es el indicador principal de la salud de los mismos (Magurran, 1988). Asimismo estos indicadores permiten comparar comunidades modificadas y naturales, a fin de medir el efecto de las actividades humanas sobre los ecosistemas (Halffter, 1998). Lo anterior posibilita diseñar estrategias de conservación a escala local (Moreno, 2001).
Objetivo
Caracterizar la estructura y diversidad de la vegetación arbórea en tres sistemas agroforestales de café y en una selva mediana subperennifolia de la Sierra de Atoyac, Veracruz. Debido a que esta selva representa la cobertura forestal original, se espera que su riqueza y diversidad de especies arbóreas sea mayor que en los tres sistemas agroforestales que la han modificado o parcialmente reemplazado.
Materiales y métodos
Área de estudio
Se localiza en la zona centro de Veracruz, en la región cafetalera de Córdoba, entre las coordenadas 18° 55'' a 19° 00' de latitud norte y 96° 54' a 96° 49' de longitud oeste. Abarca los municipios de Atoyac, Amatlán de los Reyes y Córdoba, los cuales presentan elevaciones de 298 m hasta 1800 m snm; precipitación anual de 1930 mm y temperatura media anual de 21.3 °C (Guajardo et al., 2013). El tipo de clima es (A) C (m) w"o y la vegetación se clasifica como selva mediana subperennifolia, debido a que entre 25 y 50 por ciento de los arboles pierden sus hojas en lo más acentuado de la época seca (Miranda y Hernández, 1963).
La Sierra de Atoyac se ubica dentro de la provincia geomorfológica de la Sierra Madre del Sur, en la subprovincia de sierras orientales de Oaxaca (Lugo y Córdova, 1992), su unidad geomorfológica es de montañas de laderas abruptas (pendiente >35°), de altura superior a 600 m con relieve modelado en barrancas del plioceno-cuaternario, de origen estructural por plegamiento, constituidas por rocas sedimentarias mesozoicas (Geissert y Enríquez, 2011). Hidrológicamente, el área de estudio se encuentra ubicada en la cuenca del río Papaloapan, formando parte de la cuenca del río Jamapa; y tiene como principal afluente al río Atoyac, que nace en este sistema montañoso.
Muestreo
Se llevaron a cabo recorridos previos con habitantes de las comunidades de la zona de estudio en dos franjas altitudinales (600 a 900 y 900 a 1200) m snm para localizar cuatro sistemas a estudiar: selva mediana subperennifolia (SM), rusticano (RU), policultivo complejo (PC) y policultivo simple (PS), en los cuales se establecieron aleatoriamente tres unidades de muestreo (UM) por cada franja altitudinal en cada sistema y selva mencionados, obteniendo un total de 24 UM. Las superficie de cada UM fue de 600 m2 (30 m x 20 m), que se dividió en seis cuadros de 100 m2 (10 m x 10 m) en donde se cuantificaron todos los arboles con un diámetro normal (DN: 1.3 m sobre el suelo) > 2.5 cm. A cada individuo se le registraron las siguientes variables: altura total (con flexómetro en individuos < 8 m y con clisímetro para aquellos > 8m), diámetro mayor y menor de proyección de copa (con cinta métrica), DN (con cinta diamétrica) y área basal (a partir de la variable anterior).
Asimismo, a cada individuo se le determinó su identidad taxonómica con base en los trabajos realizados anteriormente (Acevedo, 1988; Gómez Pompa, 1978; Pennington y Sarukhán, 2005); además, se recolectaron especímenes por triplicado los cuales fueron procesados y herborizados (Lot y Chiang, 1986), para cotejarlos en los herbarios XAL (Instituto de Ecología, A.C.) y CHAPA (Colegio de Postgraduados, Campus Montecillo); quedando depositados en este último.
Análisis de datos
Los atributos estructurales de área basal, densidad, altura promedio y cobertura se analizaron con una prueba de Shapiro Wilk para determinar si se ajustaban a una distribución normal. En los casos necesarios, los datos fueron transformados logarítmicamente para obtener dicha distribución. Posteriormente, se realizó un ANOVA multifactorial (franja altitudinal, sistema, UM) y cuando se encontraron diferencias significativas (P < 0.05) se realizaron comparaciones de medias mediante la prueba de Tukey Honest Significant Difference (HSD). Los análisis estadísticos se realizaron utilizando el programa Minitab versión 17 (Mckenzie et al., 1997). Las distribuciones de frecuencias de individuos por clase diamétrica (5 cm de amplitud) se obtuvieron basándose en Zarco et al. (2010). La delimitación de estratos verticales se llevó a cabo a partir de las inflexiones de una curva generada entre el número de individuos y su altura (López et al., 2012) y la curva de acumulación de especies se elaboró al graficar el número de especies encontradas respecto de la superficie muestreada (Greig-Smith, 1983).
Se calcularon en cada UM, de manera independiente así como por cada sistema, valores de densidad, dominancia y frecuencia por especie y familia, con el fin de obtener dos índices de valoración estructural:
a) Índice de valor de importancia (IVI = densidad relativa + dominancia relativa + frecuencia relativa) (Curtis y McIntosh, 1951; Villavicencio y Valdez, 2003).
b) Índice de valor forestal (IVF= DN relativo + altura relativa + cobertura relativa) (Corella et al., 2001; López et al., 2012; Carreón y Valdez, 2014).
Estos índices jerarquizan la importancia ecológica de las especies, sin embargo el primero lo hace de manera horizontal y el segundo, de manera bidimensional al incluir el parámetro altura.
Para comparar la diversidad entre sistemas y UM se calculó el índice de Shannon-Wiener (H' = -Σ(pi ln pi), donde pi es la proporción de individuos de la i-ésima especie. Del mismo modo, para contrastar la diversidad con otros estudios, se obtuvo el índice alfa de Fisher (α): {S = α ln[1 + (N / α)]}, donde S es el número de especies y N el número de individuos. Asimismo, se evaluó la equidad de las especies (E = H / ln S) y la semejanza florística con el coeficiente de Sorensen [Si = 2C / (A + B)], el cual relaciona el número de especies en común (C) con la media aritmética de las especies en ambos sitios comparados (A y B) (S0rensen, 1948; Magurran, 1988; Zarco et al., 2010).
Para determinar la existencia de diferencias significativas entre la diversidad de H de los sistemas estudiados, se aplicó el método de Hutcheson para calcular el valor de "t" modificado (Magurran, 2004; Villavicencio y Valdez, 2003; Carreón y Valdez, 2014); para la diversidad alfa y equidad se utilizó una prueba de aleatoriedad por "boots-trapping" (1000 muestras de pares al azar), la cual fue realizada por medio del programa PAST (Hammer et al., 2001).
Se llevó a cabo un análisis de agrupamiento (técnica multivariada para clasificación; programa PC-ORD Ver. 6.0) usando datos de ausencia-presencia de especies en cada UM, tomando beta flexible (β = -0.25) como método de unión de grupos, ya que puede variar la distancia entre estos y en consecuencia aumentar la agrupación de los elementos (McCune y Grace, 2002). Como medida de proximidad, se empleó el coeficiente de Sorensen por ser de los más robustos para datos ecológicos (McCune y Mefford, 1999) y evitar el efecto de encadenamiento generado con otros métodos de unión (Digby y Kempton, 1987).
Se efectuó un análisis de correspondencia canónica (técnica multivariada para ordenación; programa PC-ORD Ver 6.0) usando el IVI como atributo estructural de la vegetación para la primera matriz de datos, que se ajustó de acuerdo con lo sugerido por McCune y Grace (2002), al eliminar especies raras (presencia menor al cinco por ciento del total de sitios muestreados), quedando solo 64 especies. Además, los datos se balancearon mediante una estandarización por el máximo, al dividir el valor del atributo entre el mayor valor del mismo en la columna respectiva; esto se hizo para igualar la importancia relativa de las especies en el análisis (McCune y Grace, 2002; Peck, 2010). En la segunda matriz se utilizaron cinco variables ambientales (altitud, inclinación y exposición de la pendiente, temperatura media anual y precipitación total anual) las cuales fueron obtenidas a partir de lecturas en campo (tres primeras) y del WorldClim Global Climate GIS database (Hijmans et al., 2004); las variables ambientales fueron estandarizadas mediante transformación logarítmica, de acuerdo con (McCune y Mefford, 1999), para reducir el efecto de diferentes unidades de medida.
RESULTADOS
Se registraron un total de 917 individuos que corresponden con 90 taxones; 79 se determinaron a nivel de especie, cuatro a género, tres solo familia y cuatro desconocidos. Las especies identificadas quedaron distribuidas en 32 familias y 65 géneros. Las familias más ricas en especies (cerca de 40% del total) fueron Fabaceae con 11, Moraceae con siete, Lauraceae con seis, Euphorbiaceae y Malvaceae con cinco cada una. Los géneros más ricos en especies fueron: Coccoloba y Ficus con cuatro cada uno, así como Cordia e Inga con tres cada uno.
La selva mediana subperennifolia (SM) acumuló 65 especies, mientras que los tres sistemas agroforestales de café (SAF-café) acumularon en conjunto 73 especies: policultivo complejo (PC) con 61, rusticano (RU) con 32 y poli-cultivo simple (PS) con 18 (Fig. 1).
Se observaron dos estratos verticales (inferior y superior) generados por los siguientes puntos de inflexión: a los 10 m para SM y RU, a los 12 m para PS y a los 14 m para PC (Fig. 2). Las especies más importantes del estrato inferior y superior fueron: Myriocarpa longipes Liebm. y Cordia alliodora (Ruiz & Pav.) Cham., respectivamente. En el estrato superior se encontraron árboles emergentes de más de 30 m de altura, tales como: Ficus tecolutensis (Liebm.) Miq.; Bursera simaruba (L.) Sarg. (SM); Myroxylon balsamum (L.) Harms y Bernoullia flammea Oliver (PC); Cupania dentata DC. (PS); C. alliodora y Sideroxylon persimile (Hemsl.) T. D. Penn. (RU).
El porcentaje acumulado de individuos en las tres primeras categorías diamétricas fue mayor para la SM (79.8%), seguida del RU (77.6%), PC (64.3%) y el PS (56.7%). En la última categoría diamétrica (> 45 cm) el RU presentó un porcentaje de individuos superior respecto a los demás sistemas (8.6%) (Fig. 3). Las especies con mayor DN fueron Coccoloba hirtella Lundell y F. tecolutensis (SM), Sideroxylon capiri (DC.) Cronq. (PC), S. persimile (Hemsl.) T. D. Penn., Heliocarpus appendiculatus Turcz., Cronq. y Trichospermum mexicanum (DC.) Baill (RU).
La SM presentó los valores más elevados en densidad y área basal, el RU en cobertura (Tabla 1). En esta última variable, más la altura y DN, la SM fue significativamente (p < 0.05) menor que los tres SAF-Café.
Las especies con mayor importancia estructural fueron M. longipes y B. simaruba en la SM, así como C. alliodora y C. dentata en dos de los tres SAF-Café (PC y RU). La mayor cantidad de especies exclusivas se encontró en la SM (14), seguida por el PC (10) y el RU (6) (Tabla 2).
En general, los valores de diversidad (Shannon H', Fisher α) fueron significativamente (p < 0.05) mayores en la SM y el PC que en el RU y el PS; en contraste, los valores de equidad fueron significativamente (p < 0.05) menores en el RU que en el PC (Tabla 3).
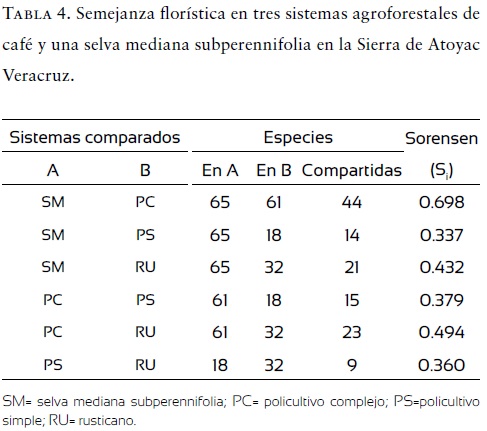
La semejanza florística fue máxima entre la SM y el PC (70%) y mínima entre la SM y el PS (Tabla 4).
El dendrograma presentó un porcentaje de encadenamiento bajo (4.81%), lo que indica buena formación de grupos. Se observan dos grupos principales (1, 2) que se diferencian por la franja altitudinal a la que pertenecen la mayoría de las UM. Dentro de estos, se observan subgrupos bastante homogéneos (1.3, 2.2) y heterogéneos (1.1, 1.2, 2.1), estos últimos con UM de la SM y PC principalmente, lo que describe la semejanza florística entre ellos (Fig. 4).
En el análisis de correspondencia canónica, las pruebas de permutación de Monte Carlo resultaron significativas (p=0.02) entre la relación especies-ambiente y los ejes de ordenación; estos últimos explicaron 20.3% de la variación de los datos. Las variables ambientales más importantes fueron: altitud (eje 1; -0.903), exposición y precipitación media anual (eje 2; 0.485 y -0.497 respectivamente).
DISCUSIÓN
La selva mediana subperennifolia (SM), en el área del presente estudio coincide en su composición de familias botánicas con Acevedo (1988) y al nivel de géneros con Chiang (1970). Asimismo especies del estrato superior como Aphanante monoica (Hemsley) Leroy, Astronium graveloens Jacq., Bursera simaruba (L.) Sarg., Brosimum alicastrum Sw., Bernulia flamea Oliver, Manilkara zapota (L.) Royen, Myroxilon balsamum (L.) Harms, Sideroxylon capiri (DC.) Cronq. y del estrato inferior como Guarea glabra Vahl, Protium copal (Schltdl. & Cham.) Engl. y Simira salvadorensis (Standl.) Steyerm., coinciden con lo descrito por Castillo-Campos et al. (2003) y Pennington y Sarukhán (2005).
Los sistemas agroforestales de café (SAF-Café) en el área de estudio concuerdan en la presencia de familias más ricas en especies con lo encontrado por Williams-Linera y López-Gómez (2008) en Veracruz (Fabaceae, Moraceae, Lauraceae) y en la riqueza de especies arbóreas (74) encontradas en cafetales bajo sombra por Soto-Pinto et al. (2007) en Chiapas. El policultivo complejo (PC) fue más rico en especies que el rusticano (RU) y el policultivo simple (PS), confirmando lo descrito por López-Gómez et al. (2008) mediante técnicas de rarefacción.
La vegetación arbórea en el área de estudio mostró dos estratos verticales para la SM, lo cual difiere de los tres estratos identificados para otras selvas medianas subperennifolias por Basáñez et al. (2008), Godínez-Ibarra y López-Mata (2002) y Castillo-Campos et al. (2003). Asimismo, el PC tuvo dos estratos verticales, a diferencia de la estructura multiestrato señalada por Raintree (1990), la cual puede deberse a la presencia de distintas cohortes generadas por el manejo de los productores para varios usos (p.ej. frutal, maderable, ornamental) (Moguel y Toledo, 2004).
La distribución diamétrica en la SM y el PC coinciden con lo descrito por Bongers et al. (1988) para un comportamiento tipo I: alta cantidad de individuos en las primeras clases diamétricas, disminuyendo gradualmente hacia las últimas, lo que indica buena reproducción, establecimiento y regeneración natural. En contraste, un comportamiento tipo II: alta cantidad de individuos en las primeras e intermedias clases diamétricas, disminuyendo hacia las últimas, se presentó en mayor medida en el PS y en menor medida en el RU. Lo anterior puede deberse a prácticas de manejo para fomentar especies de interés económico (Citrus spp., Musa sp., Chamaedorea sp.) en el estrato inferior (Moguel y Toledo 1996) y de interés maderable (Cordia alliodora (Ruiz & Pav.) Cham., Cedrella odorata L., Ocotea puberula (Rich.) Nees) con individuos de DN grandes para su aprovechamiento y sombreado de café (Inga vera Willd.), en el estrato superior (Bandeira et al., 2005).
Los valores en área basal, densidad y altura de los SAF-Café en el área de estudio coinciden con los hallados para sistemas similares en Veracruz (López-Gómez et al., 2008) y Chiapas (Peeters et al., 2003; Bandeira et al., 2005). Pero esto no ocurrió con los valores de área basal para la SM (33.6 m2 ha-1), los cuales fueron menores a los obtenidos en Vega de Alatorre (38.6 m2 ha-1; Godínez-Ibarra y López-Mata, 2002) y Papantla (76.7 m2 ha-1; Basáñez et al., 2008), Veracruz. Lo anterior puede deberse a que individuos de grandes diámetros (> 35 cm), los cuales contribuyen a tener grandes áreas basales (Galloway, 2003), representan el menor porcentaje en la SM (< 10%); a diferencia de lo calculado en RU, donde estos individuos (> 35 cm DN) representan casi el doble en porcentaje (18%).
Las especies con alto valor de importancia estructural en los tres SAF-Café del área de estudio, C. alliodora (Ruiz & Pav.) Cham., C. dentata DC. y C. odorata L., concuerdan con lo encontrado tanto por Villavicencio y Valdez (2003) como por Soto-Pinto et al. (2007) en otros SAF-Café; esto puede explicarse en el contexto del beneficio económico que se genera, por ejemplo, al vender su madera (Bandeira el al., 2005), ya que otra especie con valor similar en el mercado local es Ocotea puberula (Rich.) Nees y, al igual que las anteriores, son apreciadas y cultivadas por los productores de café (López et al., 2013).
Los valores obtenidos del índice de Shannon para la SM en este estudio (H' = 3.5) fueron superiores a los encontrados en las selvas medianas subperennifolias de Vega de Alatorre (H' = 3.3; Godínez-Ibarra y López Mata, 2002) y Papantla (H' = 2.1; Basáñez et al., 2008). Con base en los resultados obtenidos, la SM difiere estructuralmente de los tres SAF-Café, tanto en las variables dendrométricas como en las especies más importantes; sin embargo, no se encontraron diferencias estadísticas significativas del índice de Shannon entre la SM y el PC, por lo que existen evidencias para apoyar la hipótesis que este tipo de SAF-Café conserva en gran medida la diversidad de la vegetación original en el área de estudio (Perfecto et al., 1996; Moguel y Toledo, 1999).
El análisis de clasificación mostró la semejanza entre los sistemas de PC y SM lo que apoya la afirmación de que los SAF-café (policultivo complejo) tienen potencial en la conservación de la diversidad de especies arbóreas (Gordon et al., 2003); ya que a pesar de que la altitud fue la variable ambiental de mayor influencia en la composición florística, la semejanza entre los sistemas mencionados está presente tal como lo afirma Hernandéz-Martínez (2008), mediante técnicas de ordenación, lo que afirma la importancia de estudios para la caracterización de los SAF-Café.
CONCLUSIONES
La especie más importante en la estructura de los SAF-café fue Cordia alliodora (Ruiz & Pav.) Cham., con características maderables apreciadas por los productores de café, así como Bursera simaruba (L.) Sarg. y Myriocarpa longypes Liebm. para la selva mediana subperennifolia.
Valores semejantes en riqueza y diversidad se encontraron entre el policultivo complejo y la selva mediana subperennifolia, sin presentar diferencias estadísticas significativas respecto al índice de Shannon Wiener. Valores intermedios fueron para el sistema rusticano y los más bajos para el policultivo simple.
Se registraron especies catalogadas en la NOM-059 dentro de la categoría de amenazadas, tales como: Astronium graveolens Jacq. y Spondias raldkoferi J. D. Smith., con lo cual se apoya la hipótesis de que algunos SAF-café conservan en gran medida la diversidad de especies de la vegetación original.
Reconocimientos
El primer autor agradece al Conacyt por la beca otorgada para la realización de estudios de Maestría en Ciencias. A la Línea Prioritaria de Investigación 1 (Manejo sustentable de recursos naturales) del Colegio de Postgraduados, por su apoyo económico para la realización de este estudio. Al C. Hilario Barragán y familia por su amplia disponibilidad y atenciones. Al C. Adrián López Cervantes por su apoyo en los muestreos en campo.
Referencias
Acevedo R., R. 1988. La vegetación en la Sierra de Atoyac, Veracruz. México. Tesis de Licenciatura. Universidad Veracruzana. Xalapa, Veracruz. México. 102 p. [ Links ]
Bandeira, F.P., C. Martorell, J.A. Meave y J. Caballero. 2005. The role of rustic coffee plantations in the conservation of wild tree diversity in the Chinantec region of Mexico. Biodiversity and Conservation 14(5):1225-1240. [ Links ]
Basáñez, A.J., J.L. Alanís y E. Badillo. 2008. Composición florística y estructura arbórea de la selva mediana subperennifolia del ejido "El Remolino", Papantla Veracruz. Avances en Investigación Agropecuaria 12(2):3-21. [ Links ]
Beer, J., R. Muschler, D. Kass y E. Somarriba. 1998. Shade management in coffee and cacao plantations. Agroforestry Systems 38:139-164. [ Links ]
Bishop, J. y N. Landell-Mills. 2003. Los servicios ambientales de los bosques: información general. In: S. Pagiola, J. Bishop y N. Landell-Mills, eds. La venta de servicios ambientales forestales: mecanismos basados en el mercado para la conservación y el desarrollo. SEMARNAT-INE. México. p:43-75. [ Links ]
Bongers, F., J. Pompa, J. Meave del Castillo y J. Carabias. 1988. Structure and floristic composition of the lowland rain forest of Los Tuxtlas, México. Vegetatio 74(1):55-80. [ Links ]
Carreón-Santos, R.J. y J.I. Valdez-Hernández. 2014. Estructura y diversidad arbórea de vegetación secundaria derivada de una selva mediana subperennifolia en Quintana Roo. Revista Chapingo Serie Ciencias Forestales y del Ambiente 20(1):119-130. [ Links ]
Castillo-Campos, G., R. Robles G. y M.E. Medina A. 2003. Flora y vegetación de la Sierra Cruz Tetela, Veracruz, México. Polibotánica 15:41-87. [ Links ]
Castillo-Campos, G., S. Avendaño-Reyes y M.E. Medina-Abreo. 2011. Flora y Vegetación. In: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO), ed. La biodiversidad en Veracruz: Estudio de Estado. CONABIO, Gobierno del Estado de Veracruz, Universidad Veracruzana, Instituto de Ecología, A.C. México, D.F. p:163-180. [ Links ]
Chiang C., F. 1970. La vegetación de Córdoba, Ver. Tesis. Facultad de Ciencias. Universidad Nacional Autónoma de México. México D.F. 51 p. [ Links ]
Corella J., F., J.I. Valdez H., V.M. Cetina A., F.V. González C., A. Trinidad S. y J.R. Aguirre R. 2001. Estructura forestal de un bosque de mangles en el noreste del estado de Tabasco, México. Ciencia Forestal en México 26(90):73-102. [ Links ]
Curtis, J.T. y R.P. McIntosh. 1951. An upland forest continuum in the prairie-forest border region of Wisconsin. Ecology 32:476-496. [ Links ]
Digby, P.G.N. y R.A. Kempton. 1987. Multivariate analysis of ecological communities. Chapman and Hall Ltd. Londres. 206 p. [ Links ]
Ellis, E.A., M. Martínez-Bello y R. Monroy-Ibarra. 2011. Focos rojos para la conservación de la biodiversidad. In: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO), eds. La biodiversidad en Veracruz: Estudio de Estado. CONABIO, Gobierno del Estado de Veracruz, Universidad Veracruzana, Instituto de Ecología, A.C. México, D.F. p:351-368. [ Links ]
Galloway, G. 2003. Plantaciones forestales de pequeña escala. In: J. Cordero y D. Boshier, eds. Árboles de Centroamérica: un manual para extensionistas. Oxford Forestry Institute-Centro Agronómico Tropical de Investigación y Enseñanza (OFICATIE). Turrialba, Costa Rica. p:243-282. [ Links ]
Geissert K., D. y E. Enríquez Fernández. 2011. Geomorfología. In: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO). La biodiversidad en Veracruz: Estudio de Estado. CONABIO, Gobierno del Estado de Veracruz, Universidad Veracruzana, Instituto de Ecología, A.C. México, D.F. p:53-68. [ Links ]
Godínez-Ibarra, O. y L. López-Mata. 2002. Estructura, composición, riqueza y diversidad de árboles en tres muestras de selva mediana subperennifolia. Anales del Instituto de Biología serie Botánica 73(2):283-314. [ Links ]
Gómez-Pompa, A. 1978. Ecología de la vegetación del estado de Veracruz. Editorial Continental. México. 75 p. [ Links ]
Gordon, J.E., W.D. Hawthorne, G. Sandoval y A.J. Barrance. 2003. Trees and farming in the dry zone of southern Honduras II: the potential for tree diversity conservation. Agroforestry Systems 59(2):107-117. [ Links ]
Greig-Smith, P. 1983. Quantitative Plant Ecology. 3a edition. University of California Press. Berkeley, CA. 347 p. [ Links ]
Guajardo P., R., G. Díaz P., R. López M., R.H. Manson y G. Hernández M. 2014. Ordenamiento e implementación de un observatorio cafetalero en la zona centro del estado de Veracruz (OC-ZCEV). In: R. López M., G. Díaz P., eds. Cafeticultura en la zona centro del estado de Veracruz: diagnóstico, productividad y servicios ambientales. INIFAP-CIRGOC. Campo Experimental Cotaxtla, Veracruz. 405 p. [ Links ]
Halffter, G. 1998. A strategy for measuring landscape biodiversity. Biology International 36:3-17. [ Links ]
Hammer, 0, D.A.T. Harper y P.D. Ryan. 2001. PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4(1):9. [ Links ]
Hernández-Martínez, G. 2008. Clasificación agroecológica. In: R.H. Manson, V. Hernández-Ortiz, S. Gallina y K. Mehltreter, eds. Agroecosistemas cafetaleros de Veracruz: Biodiversidad, manejo y conservación. Inecol, INE-SEMARNAT. México, D.F. p:15-34. [ Links ]
Hijmans, R.J., S.E. Cameron, J.L. Parra, P.G. Jones y A. Jarvis. 2004. The WorldClim Interpolated Global Terrestrial Climate Surfaces. Version 1.3 Computer program available at http://biogeo.berkeley.edu/ (Cons. 02/2013). [ Links ]
Llorente-Bousquets, J. y S. Ocegueda. 2008. Estado del conocimiento de la biota. In: Capital natural de México, vol. I: Conocimiento actual de la biodiversidad. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. México. p:283-322. [ Links ]
López-Gómez, A.M., G. Williams-Linera y R.H. Manson. 2008. Tree species diversity and vegetation structure in shade coffee farms in Veracruz, Mexico. Agriculture, Ecosystems and Environment 124(3):160-172. [ Links ]
López M., R., J.M.P. Vázquez A., R. Martínez V. y M.A. López R. 2013. Rentabilidad de fincas de café. In: R. López M., G. Díaz P. y A. Zamarripa C., eds. El sistema producto café en México: problemática y tecnología de producción. INIFAP-CIRGOC. Campo Experimental Cotaxtla, Veracruz. 462 p. [ Links ]
López T., J.F., J.I. Valdez H., M.A. Pérez F. y V.M. Cetina A. 2012. Composición y estructura arbórea de un bosque tropical estacionalmente seco en la Reserva de la Biósfera la Sepultura, Chiapas. Revista Mexicana de Ciencias Forestales 3(12):43-56. [ Links ]
Lot, A. y F. Chiang. 1986. Manual de herbario: administración y manejo de colecciones, técnicas de recolección y preparación de ejemplares botánicos. UNAM, Instituto de Biología. México, D.F. 142 p. [ Links ]
Lugo, H.J. y F.C. Córdova. 1992. Regionalización geomorfológica de la República Mexicana. Investigaciones Geográficas Boletín del Instituto de Geografía 25:25-63. [ Links ]
Magurran, A.E. 1988. Ecological diversity and its measurement. Cambridge University Press. 179 p. [ Links ]
Magurran, A.E. 2004. Measuring biological diversity. Oxford: Blackwell Science. 256 p. [ Links ]
Manson, R.H., A. Contreras y F. López-Barrera. 2008. Estudios de la biodiversidad en cafetales. In: R.H. Manson, V. Hernández-Ortiz, S. Gallina y K. Mehltreter, eds. Agro-ecosistemas cafetaleros de Veracruz: Biodiversidad, manejo y conservación. Inecol, INE-SEMARNAT. México, D.F. p:1-14. [ Links ]
Martínez, M.A., V. Evangelista, F. Basurto, M. Mendoza y A. Cruz-Rivas. 2007. Flora útil de los cafetales en la Sierra Norte de Puebla, México. Revista Mexicana de Biodiversidad 78(1):15-40. [ Links ]
McCune, B. y J.B. Grace, 2002. Analysis of ecological communities. MjM Software Design, Gleneden Beach, Oregon. 304 p. [ Links ]
McCune, B. y M.J. Mefford. 1999. PC-ORD. Multivariate analysis of ecological data. Version 4.0. MjM Software Design. Gleneden Beach, Oregon. 237 p. [ Links ]
Mckenzie, J.; R.L. Shaefer y E. Farber. 1997. The student edition of minitab for windows, statistical software. Adapted for education. Addison Wesley Publishing Company. Menlo Park, California, USA. 622 p. [ Links ]
Méndez, V.E., S.R Gliessman y G.S. Gilbert. 2007. Tree biodiversity in farmer cooperatives of a shade coffee landscape western El Salvador. Agriculture Ecosystems Environment 119(1):145-159. [ Links ]
Miranda, F. y E. Hernández X. 1963. Los tipos de vegetación de México y su clasificación. Boletín de la sociedad Botánica de México 28:29-79. [ Links ]
Moguel, P. y V. M. Toledo. 1999. Biodiversity conservation in traditional coffee systems of Mexico. Conservation Biology 13(1):11-21. [ Links ]
Moguel, P. y V.M. Toledo. 2004. Conservar produciendo: biodiversidad, café orgánico y jardines productivos. Biodiversitas 55:2-7. [ Links ]
Moreno, C.E. 2001. Métodos para medir la biodiversidad. 1ª ed. M&T-Manuales y Tesis SEA. Zaragoza, España. 84 p. [ Links ]
Peck, J.E. 2010. Multivariate analysis for community ecologists: Step by step using PC-ORD. MjM Software Design, Gleneden Beach, Oregon, EUA. 162 p. [ Links ]
Peeters, L.Y.K., L. Soto-Pinto, H. Perales, G. Montoya y M. Ishiki. 2003. Coffee production, timber, and firewood in traditional and Inga-shaded plantations in Southern Mexico. Agriculture, Ecosystems and Environment 95(2):481-493. [ Links ]
Pennington, T. D. y J. Sarukhán. 2005. Árboles Tropicales de México. Manual para la identificación de las principales especies. 3a ed. Ediciones científicas universitarias. UNAM-Fondo de Cultura Económica. México, D.F. 523 p. [ Links ]
Perfecto, I., R.A. Rice, R. Greenberg y M.E. Van der Voort. 1996. Shade coffee: a disappearing refuge for biodiversity. BioScience 46(8):598-608. [ Links ]
Pineda-López, M. del R., G. Ortiz-Ceballos y L.R. Sánchez-Velásquez. 2005. Los cafetales y su papel en la captura de carbono: un servicio ambiental aún no valorado en Veracruz. Madera y Bosques 11(2):3-14. [ Links ]
Raintree, J.B. 1990. Theory and practice of agroforestry diagnosis and design. In: MacDicken, K.G. y N.T. Vergara, eds. Agroforestry: Classification and Management. John Wiley & Sons. Nueva York, EUA. p:58-97. [ Links ]
Ramamoorthy, T.P., R. Bye, A. Lot y J. Fa. 1993. Biological diversity of México: origins and distribution. Oxford University Press. Nueva York. 812 p. [ Links ]
Richter, A., M.A. Klein, T. Tscharntke y M. Jason. 2007. Abandonment of coffee agroforests increases insect abundance and diversity. Agroforestry Systems 69(3):175-182. [ Links ]
Roncal-García, S., L. Soto-Pinto, J., Castellanos-Albores, N. Ramírez-Marcial y B. de Jong. 2008. Sistemas agroforestales y almacenamiento de carbono en comunidades indígenas de Chiapas, México. Interciencia 33(3):200-206. [ Links ]
Rzedowski, J. 1978. Vegetación de México. Limusa. México. 504 p. [ Links ]
Schroth, G., C.A. Harvey y G. Vincent. 2004. Complex Agroforests: Their Structure, Diversity, and Potential Role in Landscape Conservation. In: G. Schroth, G.A.B. Fonseca, C.A. Harvey, C. Gascon, H.L. Vasconcelos y A.M.N. Izac, eds. Agroforestry and biodiversity conservation in tropical landscapes. Island Press, Washington, DC. p:227-260. [ Links ]
SEMARNAT (Secretaria de Medio Ambiente y Recursos Naturales). 2013. Informe de la situación del medio ambiente en México. Compendio de estadísticas ambientales. Indicadores clave y de desempeño ambiental. Edición 2012. México, D.F. 361 p. [ Links ]
Sorensen, T.A. 1948. A method of establishing groups of equal amplitude in plant sociobiology based on similarity of species content and its application to analyses of vegetation in Danish commons. Biologiske Skrifter 5:1-40. [ Links ]
Soto-Pinto, L., V. Villalvazo-López, G. Jiménez-Ferrer, N. Ramírez-Marcial, G. Montoya y F.L. Sinclair. 2007. The role of local knowledge in determining shade composition of multistrata coffee systems in Chiapas, Mexico. Biodiversity and Conservation 16(2):419-436. [ Links ]
Soto-Pinto, L., Y. Romero-Alvarado, J. Caballero-Nieto y G. Segura Warnholtz. 2001. Woody plant diversity and structure of shade-grown-coffee plantations in Northern Chiapas, Mexico. Revista de Biología Tropical 49:977-987. [ Links ]
Toledo, V.M. y P. Moguel. 2012. Coffee and sustainability: the multiple values of traditional shaded coffee. Journal of Sustainable Agriculture 36(3):353-377. [ Links ]
Vázquez-Torres, M., 1998. Humedal de Alvarado: diversidad vegetal. In: M. Vázquez T., ed. Biodiversidad y problemática en el humedal de Alvarado, Veracruz, México. Universidad Veracruzana. Xalapa. p:143-168. [ Links ]
Villavicencio E., L. y J.I. Valdez H. 2003. Análisis de la estructura arbórea del sistema agroforestal rusticano de café en San Miguel, Veracruz, México. Agrociencia 37(4):413-423. [ Links ]
Williams-Linera, G. y A. López Gómez. 2008. Estructura y diversidad de la vegetación leñosa. In: R.H. Manson, V. Hernández-Ortiz, S. Gallina y K. Mehltreter, eds. Agro-ecosistemas cafetaleros de Veracruz: Biodiversidad, manejo y conservación. Inecol, INE-SEMARNAT. México, D.F. p:55-63. [ Links ]
Zarco E., V.M., J.I. Valdez H., G. Ángeles P. y O. Castillo A. 2010. Estructura y diversidad de la vegetación arbórea del Parque Estatal de Agua Blanca, Macuspana, Tabasco. Universidad y Ciencia 21(1):1-17. [ Links ]
Nota
Este documento se debe citar como: García M. L.E., J.I. Valdez H., M. Luna C. y R. López M. 2015. Estructura y diversidad arbórea en sistemas agroforestales de café en la Sierra de Atoyac, Veracruz. Madera y Bosques 21(3):69-82.

