











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Los matorrales son comunidades que se encuentran generalmente dominadas por plantas leñosas de 0.5 m a 5 m o más de altura con los tallos ramificados desde la base (Medrano, 2003). Estas comunidades se distribuyen principalmente en las zonas semiáridas, que componen aproximadamente 40% de la superficie del país, por lo tanto conforman el más vasto de todos los tipos de vegetación de México (Instituto Nacional de Estadística y Geografía [Inegi], 2009).
El matorral espinoso tamaulipeco (MET) cubre una superficie de 200 000 km² del noreste de México y sur de Texas, desde Llera de Canales y los límites de la Sierra Azul en Tamaulipas (González, 1985) hasta el Altiplano Edwards (“Edwards Plateau”) en Texas (Diamond, Risking y Orzell, 1987), y de la Sierra Madre Oriental hasta el Golfo de México (Jurado y Reid, 1989). Esta vegetación está conformada por una alta riqueza (> 35) y densidad (> 15 000 individuos·
Existen algunos estudios en donde se observa la diversidad y distribución territorial del matorral espinoso tamaulipeco, así como su relación con los diversos factores que influyen y determinan la composición de las comunidades vegetales resultantes. Entre estos estudios se puede mencionar a Villegas (1972), quien describe un total de seis unidades de vegetación, y destaca como más importante el matorral alto subinerme cubriendo 35% de la superficie de Linares y Hualahuises (Nuevo León, México). Heiseke y Foroughbahkck (1985) caracterizaron la estructura de dos tipos de matorral en Linares, N.L., citando los datos más relevantes de las especies más comunes: frecuencia, abundancia, dominancia y valor de importancia. Jurado (1986) analizó la influencia, asociación y correlación de las especies con respecto a los factores físicos del suelo, la topografía y el disturbio. Jurado y Reid (1989) caracterizaron un área del matorral espinoso tamaulipeco, analizando la influencia que el disturbio y los factores edáficos y topográficos tienen sobre la distribución de sus especies. Reid, Marroquín y Beyer (1990) realizaron un estudio sobre la variación florística y estructural en el matorral tamaulipeco en el noreste de México. Rodríguez (1994) determinó la composición florística y estructural de dos comunidades diferentes de matorral en el área de Linares, comparándolos a través del uso de diferentes índices de diversidad. Así como González (1996) quien analizó la vegetación secundaria del matorral espinoso tamalupeco presente en el municipio de Linares, N.L.
La amplia diversidad de plantas nativas del matorral espinoso tamaulipeco es un ejemplo típico de una gran plasticidad en la respuesta a factores físicos extremos (Reid et al., 1990), en especial, a las sequías que se presentan año con año en la región. En particular, las características del clima y suelo del noreste de Nuevo León no son uniformes (Moya, Ramírez, Foroughbackhch, Hauad y González, 2002).
La planicie costera del noreste en México (Rzedowski, 1978) abarca casi la totalidad de Tamaulipas, los dos tercios nororientales de nuevo león, además de pequeñas áreas de Coahuila, de San Luis Potosí y del extremo norte de Veracruz. La vegetación está constituida en su mayor parte por el bosque espinoso y por matorrales xerófilos. Existen investigaciones dirigidas a identificar las comunidades vegetales presentes dentro de la planicie costera del noreste en donde se considera el gradiente altitudinal existente en la zona. Muller (1939) definió para el norte del estado de Nuevo León el matorral espinoso tamaulipeco, Edwards (1939) señaló que hacia la parte norte de la Sierra Madre Oriental se presenta una asociación desértica arbustiva de Acacia-Prosopis en la planicie y una vegetación arbustiva dominada por Helietta parvifolia en las colinas bajas. Rojas-Mendoza (1965), en su trabajo sobre la vegetación de Nuevo León, indicó que en la zona se presentan matorrales bajos subperennifolios con Acacia-Leucophyllum-Cordia, matorrales micrófilosuculentos con Prosopis-Bouteloua-Opuntia, matorrales bajos subcaducifolios con Cordia-Acacia-Karwinskia y matorrales altos subperennifolios con Acacia-HavardiaHelietta. Sin embargo existe escasa investigación enfocada a conocer las diferencias y similitudes en comunidades vegetales en condiciones de loma y valle del matorral espinoso tamaulipeco.
Objetivo
Identificar las diferencias y similitudes de la estructura de dos comunidades vegetales en diferentes áreas (loma y valle) del matorral espinoso tamaulipeco en el noreste de México.
Materiales y métodos
Área de estudio
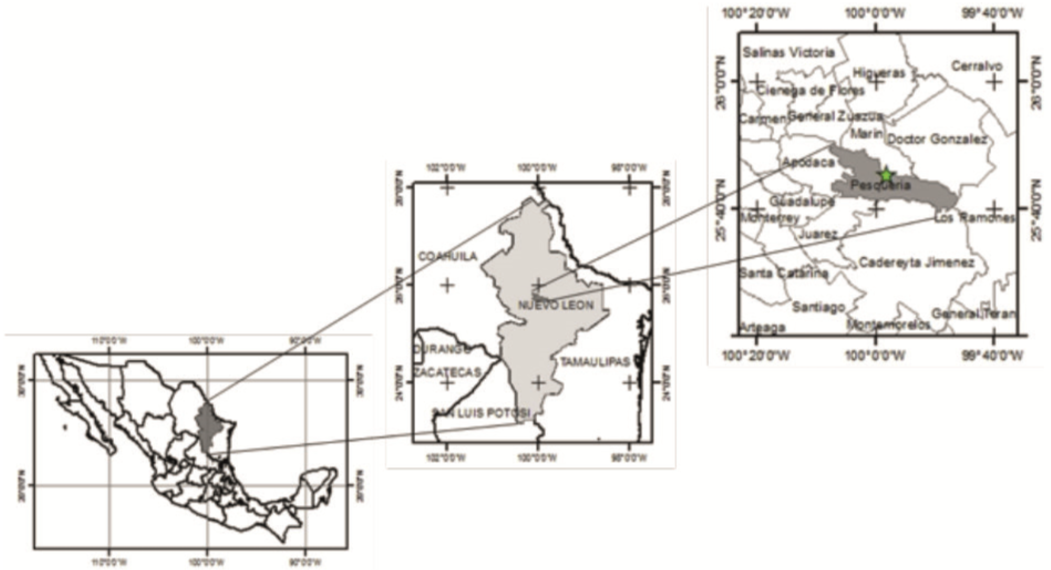
El trabajo de campo se desarrolló en el municipio de Pesquería, Nuevo León (Noreste de México, Fig. 1). Dentro de la reserva ecológica de flora y fauna de la empresa Ternium. Las coordenadas de ubicación son 25°45´17´´ de latitud Norte y 99° 58´01´´ de longitud Oeste; a 330 m snm. El clima predominante es muy seco semicálido (BWhw), con una temperatura media anual entre 20 °C y 21 °C (García, 1964). La precipitación anual oscila entre 500 mm y 700 mm (Inegi, 2010). Los tipos de suelos presentes son Kastañozem, Vertisol, Calcisol, Leptosol, Chernozem y Fulisol (Inegi, 2010).
Inventario florístico
Se establecieron tres sitios de muestreo en dos localidades diferentes (loma y valle) del matorral espinoso tamaulipeco. Lo que equivale a 1.5% y 3.4% respectivamente dentro del área de estudio. El matorral de porte bajo se ubica en lomeríos, donde el suelo es somero, más pedregoso, la disponibilidad de agua en la zona se limita a las temporadas de lluvia de la región y tiene una superficie de 32 ha. Mientras que el matorral de porte alto se localiza en una zona de caudal intermitente el cual aparece en temporadas de lluvias y forma un cuerpo de agua temporal, los suelos son profundos y tiene una superficie de 14 ha. Las parcelas de muestreo fueron cuadros de 1600 m2 (40 m × 40 m) apropiados para vegetación densa (Canizales et al., 2009). En los sitios de muestro se realizó un censo de todas las especies arbóreas y arbustivas (
Análisis de datos
Para cada especie se determinó su abundancia, de acuerdo con el número de individuos, su cobertura, en función del área de copa, y su frecuencia con base en su presencia en los sitios de muestreo. Las variables relativas se utilizaron para obtener un valor ponderado a nivel de taxón denominado Índice de Valor de Importancia (IVI), que adquiere valores porcentuales en una escala de 0 a 100 (Mostacedo y Fredericksen, 2000). Para la estimación de la abundancia relativa se empleó la siguiente ecuación:
 (Ec. 1)
(Ec. 1)
donde ARi es la abundancia relativa de la especie i respecto a la abundancia total y Ai es la abundancia absoluta de la especie i (N/ha). La dominancia se evaluó mediante la ecuación 2 (Romahn and Ramírez, 2006).
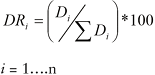 (Ec. 2)
(Ec. 2)
donde DRi es la dominancia relativa de la especie i respecto a la dominancia total y D es la dominancia absoluta de la especie i (
 (Ec. 3)
(Ec. 3)
 (Ec. 4)
(Ec. 4)
donde Fi es la frecuencia absoluta (porcentaje de presencia en los sitios de muestreo), fi es el número de sitios en la que está presente la especie i, N es el número de sitios de muestreo y FRi es la frecuencia relativa de la especie i respecto a la frecuencia total. El índice de valor de importancia (IVI) se define a través de la ecuación 5 (Whittaker, 1972).
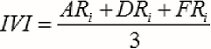 (Ec. 5)
(Ec. 5)
Para estimar la diversidad alfa se utilizó el índice de Margalef (
 (Ec. 6)
(Ec. 6)
 (Ec. 7)
(Ec. 7)
donde S es el número de especies presentes, N es el número total de individuos y ni es el número de individuos de la especie i.
La similitud entre los matorrales se determinó mediante el método de Sørensen cuantitativo (IS), el cual está basado en la relación presencia-ausencia del número de especies compartidas en cada sitio y el número total de especies de los dos sitios que están siendo comparados. Se expresa mediante la ecuación 8 (Magurran, 1988).
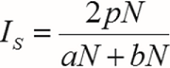 (Ec. 8)
(Ec. 8)
Donde:
aN = número total de individuos en el sitio A,
bN= número total de individuos en el sitio B,
pN= sumatoria de la abundancia más baja de cada una de las especies compartidas entre ambos sitios.
Para determinar las diferencias en las variables de abundancia, dominancia, índice de Margalef e índice de Shannon entre los matorrales se estimaron los valores promedio de los sitios de muestreo. Una vez que los datos cumplieron con los criterios de normalidad y homocedasticidad se realizó una prueba de t con un factor (α = 0.05). Asimismo, se realizó un análisis de componentes principales (PCA) con los datos de abundancia y dominancia absoluta de cada especie de cada área. Los análisis mencionados se realizaron mediante el paquete estadístico SPSS 15.0 (SPSS Inc., 2006).
Resultados
En los dos tipos de matorral se registraron 17 especies pertenecientes a 11 familias y 15 géneros. La familia con mayor presencia en el estudio fue Fabaceae con siete especies, el resto de las familias presentó solamente una especie. El género con más especies fue Acacia con tres especies registradas. De acuerdo con su forma de vida, cinco fueron arbóreas, 11 arbustivas y una palma (Tabla 1).
Tabla 1 Nombre científico, común, familia y forma de vida de las especies presentes en los dos sitios de estudio.

Del total de las 17 especies registradas en el estudio, 14 fueron identificadas en el matorral de porte alto y siete en el matorral de porte bajo. Cuatro especies tuvieron presencia en ambos tipos de matorral.
En la tabla 2se puede observar que en el matorral de porte bajo las especies con los valores más altos en abundancia, dominancia e importancia fueron Leucophyllum frutescens, Cordia boissieri y Acacia amentacea, sumando 79.92% de importancia de la comunidad. Las cuatro especies restantes suman 20.08% de importancia. En el matorral de porte alto las especies que presentan los mayores valores registrados en abundancia dominancia e importancia fueron Prosopis glandulosa, Acacia amentacea, Havardia pallens, Acacia farnesiana, Parkinsonia texana y Celtis pallida, las cuales en conjunto suman 69.17% de importancia de la comunidad. Las restantes ocho especies concentran 30.83%.
Tabla 2 Datos de abundancia, dominancia, frecuencia, IVI y alturas para ambos tipos de matorrales.

Las especies están ordenadas de manera decreciente de acuerdo al Índice de Valor de Importancia (IVI). IVI= Índice de Valor de Importancia, Max= Máxima, Min= Mínima.
En la tabla 2 se observa que las especies que se ven favorecidas en ambientes propicios son Prosopis glandulosa, Acacia amentacea, Havardia pallens, Acacia farnesiana, Parkinsonia texana y Celtis pallida ya que se desarrollan en áreas con suelos más profundos, ricos en nutrientes y con regímenes favorables de agua, pues en la zona de matorral de porte alto existen arroyos intermitentes, los cuales se activan en temporadas de lluvia. Mientras que las especies más tolerantes a ambientes hostiles registradas son Leucophyllum frutescens, Cordia boissieri y Acacia amentacea, ya que concentran el mayor valor de importancia en el matorral de porte bajo donde los suelos son menos profundos y la disponibilidad de agua es relativamente limitada a época de lluvias. De igual manera se observa que las especies que registran los mayores valores en altura promedio para ambos tipos de matorral son Parkinsonia texana y Acacia amentacea, seguida de Forestiera angustifolia y Cordia boissieri. Asimismo se puede observar la influencia que tiene las condiciones de suelo, humedad y nutrientes en el matorral con porte alto ya que en él se encuentran las mayores alturas promedio observadas en comparación con las registradas en el matorral en monte de porte bajo.
En el matorral de porte alto se registró una altura promedio de 5.56 m ± 1.48 m, observando que la mayoría de individuos están concentrados en un intervalo de 4 m a 6 m de altura, mientras existe un poca cantidad de individuos que su altura sea superior a los 8 m. Por otra parte, en el matorral de porte bajo se registró una altura promedio de 2.25 m ± 0.51 m, se aprecia que la mayoría de los individuos registrados se encuentran en un intervalo de 2 m a 4 m, mientras que un escaso número de individuos son los que superan una altura de 6 m en este tipo de comunidad vegetal.
La mayor densidad de individuos registrada en el área de matorral de porte bajo fue de 808
individuos

(a) Densidad, (b) área de copa, (c) índice de Margalef, (d) índice de Shannon. Valores promedio ± desviación estándar. Letras diferentes indican diferencia significativa (p = 0.05)
Figura 2 Diferencias entre el matorral de porte alto (monte alto) y porte bajo (monte bajo) evaluados.
El índice de Margalef mostró diferencia significativa entre ambos matorrales (P < 0.001). El matorral con menor valor fue el matorral con porte bajo registrando 0.96 ± 0.13 mientras que el matorral con porte alto registró valores de 2.13 ± 0.13 (Fig. 2c). El índice de Shannon registró valores de 1.20 ± 0.05 para el área de matorral con porte bajo y 1.81 ± 0.16 para el matorral con porte alto, mostrando también diferencias significativas (P = 0.003. Fig. 2d).
De igual manera se evaluó el índice de Sorensen cuantitativo para conocer la similitud existente entre el área de matorral de porte alto y el área de matorral de porte bajo. Se determinó que la similitud entre las comunidades vegetales es baja, presentando un valor de 19% de similitud. Este bajo porcentaje de similitud está influenciado por que tan solo comparten cuatro de las 17 especies registradas.
Los dos primeros componentes del PCA realizado a la matriz de datos de abundancia explicaron 71.6% de la varianza (47.7% el primer eje y 23.9% el segundo componente). Las especies que más peso tienen en el primer componente son, por este orden, Celtis pallida, Guaiacum angustifolium, Sideroxylon celastrinum, Prosopis glandulosa, Diospyros palmeri y Havardia pallens. (Tabla 3). Por su parte, el segundo componente está principalmente condicionado por la abundancia de Acacia schaffneri y Zanthoxylum fagara y en menor medida y con peso negativo, A. farnesiana. La representación gráfica de los puntos de muestreo tras este análisis muestra dos grupos bien definidos en donde se aprecia que hay diferencias entre los dos tipos de matorrales (Fig. 3). Así, las parcelas del matorral de porte alto presentan valores positivos del primer componente, indicando una mayor abundancia de las estas especies que el matorral de porte bajo. Por su parte, a lo largo del segundo componente, el matorral con porte bajo presenta valores en su mayoría positivos, asociados a A. schaffneri y Zanthoxylum fagara; mientras que el matorral con porte alto presenta en su mayoría valores negativos que se relacionan con alta abundancia de A. farnesiana.
Tabla 3 Peso de las abundancias de las distintas especies sobre los dos primeros componentes del PCA.

Valores próximos a +1 y -1 indican un elevado peso directa e inversamente proporcional respectivamente, de la especie sobre el componente, mientras que valores próximos a 0 indican escasa importancia de esa especie.

Figura 3 Representación gráfica de los dos tipos de matorrales en los dos primeros componentes del PCA de la abundancia de especies presentes.
En el análisis realizado sobre los datos de dominancia, los dos primeros ejes explicaron 71.4% de la varianza (44.3% el primer eje y 27.1% el segundo). La representación de los resultados muestra bien definido el grupo del matorral con monte alto, mientras que en el grupo de matorral de monte bajo los resultados se muestran dispersos. En este caso, la dominancia de Diospyros palmeri, Guaiacum angustifolium, Acacia amentacea, Celtis pallida, Havardia pallens, Acacia schaffneri y Zanthoxylum fagara condicionan de manera positiva el primero de los componentes, mientras que Foresteria angustifolia, Acacia schaffneri y Zanthoxylum fagara, son las que determinan la distribución de las especies a lo largo del segundo componente (Tabla 4). Los dos tipos de matorrales se distribuyen en estos dos ejes registrando en su mayoría valores positivos, con la única diferencia que una minoría de los valores observados en el matorral con monte alto a lo largo del primer componente presenta valores negativos (Fig. 4).
Tabla 4 Peso de las dominancias de las distintas especies sobre los dos primeros componentes del PCA.

Valores próximos a +1 y -1 indican un elevado peso directa e inversamente proporcional, respectivamente, de la especie sobre el componente, mientras que valores próximos a 0 indican escasa importancia de esa especie.
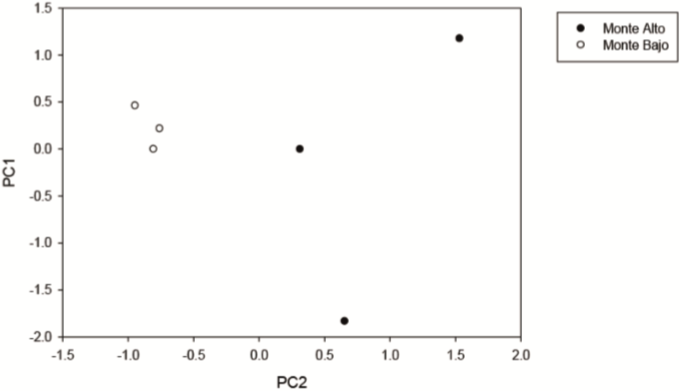
Figura 4 Representación gráfica de los dos tipos de matorrales en los dos primeros componentes del PCA de la dominancia de especies presentes.
En la Tabla 4 se encuentra el valor de la dominancia de cada especie por los dos tipos de matorrales. En general, se comparten cinco especies que condicionan de manera positiva en los dos componentes para dominancia y abundancia registrando valores similares en ambos análisis.
Discusión
El total de las especies registradas en las dos áreas de matorral espinoso tamaulipeco para este estudio fue de 17, pertenecientes a 11 familias y 15 géneros. Estos valores son inferiores comparados con la riqueza especifica registrada por Molina, Rechy, Alcalá y Alanís (2014), quienes en un estudio de composición y diversidad vegetal del matorral mediano subinerme aledaña al área de estudio, registraron un total de 32 especies pertenecientes a 19 familias y 29 géneros en una zona de matorral espinoso tamaulipeco. También son inferiores a los valores registrados por Jiménez, Alanís, González, Aguirre y Treviño (2013) quienes registraron una riqueza específica de 29 especies, pertenecientes a 14 familias y 29 géneros en un estudio de regeneración de especies maderables en áreas con diferentes historiales de uso silvoagropecuario en el matorral espinoso tamaulipeco. De igual manera, los valores registrados en este estudio resultan bajos comparados con los registrados por Domínguez et al (2013), quienes realizaron un estudio de diversidad estructural del MET durante la época seca y húmeda, registrando para la primera un total de 38 especies pertenecientes a 17 familias y 20 géneros, mientras que durante la época húmeda registraron 53 especies pertenecientes a 24 familias y 28 géneros. Es importante mencionar que en su trabajo de investigación tomaron en cuenta todos los tipos de vida de las especies vegetales mientras que en la presente investigación solo fueron consideradas arbóreas y arbustivas (
Para el matorral con porte bajo, la especie con mayor índice de valor de importancia fue Leucophyllum frutescens registrando 37.84%, seguida de Cordia boissieri y Acacia amentacea quienes en conjunto concentran un total de 79.92% de IVI. En el matorral con porte alto Prosopis glandulosa es la especie con el valor máximo registrado de índice de valor de importancia (15.95%), seguida por Acacia amentacea, Havardia pallens y Acacia farnesiana concentrando un total de 55.94%, mientras que el restante 44.06% es dividido en las 10 especies restantes presentes. Este comportamiento es común en las comunidades vegetales maduras del matorral espinoso tamaulipeco, donde pocas especies son altamente importantes y un elevado número de especies presentan poca importancia (Mora et al., 2013a). Cuatro de las 14 especies registradas en el matorral con porte alto fueron observadas en un estudio similar realizado por Heiseke y Foroughbakhch (1985), para un matorral en planicie con condiciones similares a las áreas de estudio de esta investigación, siendo Forestiera angustifolia la especie con mayor valor de IVI (18.7%), mientras que en esta investigación en el área de monte con porte alto Forestiera angustifolia registró un valor de IVI de 3.36%.
De igual manera, Heiseke y Foroughbakhch (1985) observaron dentro del mismo estudio en Diospyros palmeri el menor valor registrado de IVI, mientras que en la presente investigación el valor registrado fue superior, concentrando 5.06% en el matorral con porte alto.
Referente a la abundancia, los valores registrados son bajos comparados con los registrados por Jimenez et al. (2013) en los 4 historiales de uso silvoagropecuarios, presentando valores máximos de 16 810 individuos
La dominancia presenta valores registrados de 2725 m2
El matorral con menor valor de índice de Margalef fue el de porte bajo, registrando 0.96 ± 0.13; mientras que el matorral con porte alto registró valores de 2.13 ± 0.13. El valor del matorral con porte alto coincide con lo registrado por Mora et al. (2013b), quienes observaron valores de 2.16 ±0.17 en un matorral de referencia en el matorral espinoso tamaulipeco.
El índice de Shannon registró valores de 1.20 ± 0.05 para el área de matorral con porte bajo y 1.81 ± 0.16 para el área con matorral con porte alto. Estos valores son similares a los registrados por Mora et al. (2013a), quienes evaluaron tres áreas del matorral espinoso tamaulipeco, un área de referencia (1.95±0.17), un área de regeneración (1.31±0.02) y un área regenerada postganadería (1.21±0.06). También son similares a los valores registrados por Pequeño et al. (2012), quienes registraron valores de H`= 1.27 en un estudio de regeneración pasiva postpecuaria. Sin embargo, son bajos comparados con los valores registrados por Molina-Guerra et al. (2013), quienes estudiaron la composición y diversidad vegetal de dos sistemas de pastoreo en el MET del noreste de México y obtuvieron valores de diversidad alfa H´= >2.22 y 2.11.
Las áreas de este estudio presentan una similitud baja (19%), ya que solo comparten cuatro de las especies presentes en ambos tipos de matorrales. Autores como Jurado y Reid (1989) mencionan que los factores relacionados con la humedad del suelo son los más importantes en la distribución de la vegetación. De esta manera, la coincidencia de la vegetación más alta con un arroyo temporal concuerda con lo observado en las partes cercanas a cuerpos de agua en la región, donde se encuentra una vegetación más alta, densa y de composición diferente comparada con la de un matorral aledaño donde no se presentan este tipo de condiciones.
Conclusiones
De acuerdo con los resultados de la presente investigación, se concluye que las comunidades vegetales desarrolladas en lomas y valles presentan diferencias significativas en densidad, área de copa, riqueza y diversidad. El matorral presente en la loma presenta menor altura, área de copa, riqueza y diversidad de especies, pero mayor densidad. La comunidad vegetal presente en el valle registra mayores valores de altura, área de copa, riqueza y diversidad de especies. Esto puede deberse al contraste en las condiciones de profundidad, nutrientes y humedad del suelo; ya que en temporada de lluvia en la zona de matorral en monte bajo se crea un arroyo intermitente, el cual puede propiciar las condiciones favorables para el desarrollo de más variedad de especies en la zona. También se concluye que las comunidades presentan una baja similitud florística, compartiendo únicamente cuatro especies, de las cuales solo Acacia amentacea es abundante en ambos matorrales.

