











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
Characterized by exceptional concentrations of biodiversity and endemicity (Scatena, Bruijnzeel, Bubb, & Das, 2011), tropical montane cloud forests (TMCF) are subjected to significant deforestation and degradation due to land use change, fragmentation and unauthorized selective logging (Scatena et al., 2011; Toledo-Aceves, Meave, González-Espinoza, & Ramírez-Marcial, 2011). In Mexico, only 28% of the original TMCF cover remained by 2002 and, of this, 52.4% corresponded to degraded or secondary forests (Challenger et al., 2009). Important triggers of TMCF loss and degradation include government programs promoting agriculture and cattle grazing in areas of TMCF distribution, a lack of support for sustainable forestry operations and for commodity market production (Bray & Merino, 2005; Martínez et al., 2009; Toledo-Aceves et al., 2011). Frequently located on very steep slopes and with low productivity, both mature and secondary TMCF are not regarded as of high value for timber harvesting (Scatena et al., 2011; Toledo-Aceves et al., 2011). Nevertheless, unplanned selective logging is a customary subsistence practice among forest owners (Toledo-Aceves et al., 2011; Bárcenas & Ordóñez, 2008; Ortiz-Colín, Toledo-Aceves, López-Barrera, & Gerez-Fernández, 2017). Extraction of TMCF tree species from remnant forest fragments with no management planning contributes to increased degradation and the depletion of valuable species in these forests and can ultimately lead to their transformation into agriculture land and pine plantations (Ramírez-Marcial, González-Espinosa, & Williams-Linera, 2001; Haeckel 2006; Rüger, Williams-Linera, Kissling, & Huth, 2008; Paré & Gerez 2012). The lack of management strategies is exacerbated by a lack of pertinent information for the silvicultural management of TMCF species (Comisión Nacional para el Conocimiento y Uso de la Biodiversidad [Conabio], 2010). As a result of TMCF deforestation and illegal logging, 60% of TMCF tree species in Mexico are in some category of threat (González-Espinoza, Meave, Lorea-Hernández, Ibarra-Manríquez, & Newton, 2011).
TMCF species have shown promising potential for timber harvesting where appropriate management schemes are put into place (aus der Beek & Saenz, 1996; Saénz & Guariguata, 2001; Guariguata & Saénz, 2002). Planned selective logging could provide income through the commercialization of timber and promote the maintenance of forest cover and habitats as well as contributing to ensuring the provision of ecosystem services (Porter-Bolland et al., 2011; Putz et al., 2012). Previous studies indicate that several TMCF species-such as Alnus acuminata, Fraxinus uhdeii, Clethra macrophyla and various species of Quercus-produce good quality timber for construction, poles, furniture and paper (Tamarit Urias, 1996; Bárcenas & Ordóñez, 2008). However, the growth rate of most TMCF species remains to be determined (Williams-Linera, 1996); the lack of this key information prevents the determination of sustainable harvesting rates for these native tree species (Conabio, 2010). Information on individual tree diameter growth, and its response to environmental variables, is necessary to select which tree to cut, to estimate cutting cycles and to prescribe adequate silvicultural treatments (Pereira et al., 2002).
Tree growth in natural populations usually exhibits a wide variation that is affected by biotic and abiotic factors (Harper, 1977; Burkhart & Tomé, 2012). Climatic factors such as temperature and precipitation have a strong influence on tree growth (Dong et al., 2012), as do biotic interactions such as competition for light, water and nutrients (Canham et al., 2006; Saenz & Guariguata, 2001). Individual tree growth in young secondary evergreen broadleaf forest in China was found to be largely a function of competition with neighbouring trees (von Oheimb et al., 2011). Light availability is one of the most important factors regulating tree growth and is a key factor that is frequently manipulated in silvicultural schemes to promote tree development (Lamprecht, 1989). However, the influence of environmental factors, such as the light availability and the competition with neighboring trees, on variation in the growth rates of TMCF tree species has not been assessed. While the potential role of secondary tropical forests as a timber source is widely recognized, there is limited information available with which to design effective management schemes (Henao, Ordóñez, de Camino, Villalobos, & Carrera, 2015). Understanding the factors that influence tree growth in early successional stands is important to accurately determine timber yields in secondary tropical forests, which are characterized by mixed-species and uneven-aged stands (Adame, Brandeis, & Uriarte, 2014).
Objectives
In order to contribute to the sustainable management of TMCF, the objective of this study was to determine the diametric growth rates of three common tree species of commercial value in secondary TMCF and to evaluate the influence of tree size, crown class and competition on their growth rates in Mexico.
Materials and methods
Species
The tree species selected for study were Alnus acuminata Kunth (Betulaceae), Liquidambar styraciflua L. (Altingiaceae) and Quercus xalapensis Bonpl. (Fagaceae), all of which are locally important for the production of timber, firewood and charcoal. Alnus acuminata and L. styraciflua are light-demanding or pioneer species that require open areas for early establishment (Niembro, Morato, & Cuevas, 2004), while Q. xalapensis is of intermediate shade tolerance, establishes under the closed forest canopy and can display high growth rates in open areas (Álvarez-Aquino, Williams‐Linera, & Newton, 2004; Niembro et al., 2004; Muñiz-Castro, Williams‐Linera, & Benítez‐Malvido, 2015). According to the Red List of Mexican Cloud Forest Trees, A. acuminata and L. styraciflua are classified as being of least concern, while Q. xalapensis is critically endangered (González-Espinosa et al., 2011).
Study sites
The study region is located in the mountainous watershed of the River Pixquiac in central Veracruz, Mexico. The annual mean temperature in the region is 14 °C - 16 °C and annual precipitation ranges between 1500 mm and 1800 mm (Paré & Gerez, 2012). The study was carried out from June 2012 to July 2014, in secondary TMCF fragments with no management, and which met the following requirements: forest fragment area ~1 ha, an absence of cattle inside the forest, slope ≤ 40°, dominance in the canopy of the tree species selected including natural regeneration, a minimum number of 30 trees per species covering a range of sizes, and permission from the forest owners. All the sites were the result of natural regeneration after pasture abandonment. Based on the land use dynamics in the region, the estimated age of study sites is ~30 years (Paré & Gerez, 2012). The selected tree species were studied in two different sites (Table 1). The soil at all the sites is classified as an Umbric Andosol (Geissert, 2007).
Table 1 Characteristics of tropical montane cloud forest sites for the assessment of diameter tree growth rates, Veracruz, Mexico.
| Site | Alnus acuminata | Liquidambar styraciflua | Quercus xalapensis | |
| Coordinates | 1 | 19° 30' 20" N 97° 00' 77" W |
19° 31' 65" N 96° 59' 23" W |
19° 31' 32" N 96° 58' 51" W |
| 2 | 19° 31' 53" N 97° 02' 33" W |
19° 30' 73" N 97° 01' 01" W |
19° 31' 98" N 97° 00' 48" W |
|
| Elevation (m a.s.l.) | 1 | 1720 | 1467 | 1451 |
| 2 | 2205 | 1849 | 1672 | |
| Aspect | 1 | NE | E | SE |
| 2 | SE | E | E | |
| Slope (°) | 1 | 35 | 30 | 15 |
| 2 | 30 | 25 | 15 | |
| Basal area (m2/ha) | 1 | 9.1 ± 0.91 | 16 ± 1 | 16 ± 0.5 |
| 2 | 9.0 ± 0.60 | 13 ± 1 | 11 ± 0.9 |
Experimental design and measurements
For each species, 30 trees were selected at each site, comprising a range of sizes between 10 cm and 45 cm of diameter at breast height (dbh). In total, 60 individuals per species were measured. The selected trees had to meet the following criteria: healthy, with no bifurcation below 1.3 m in height and no mechanical damage. Based on crown position, every individual was classified into one of the following crown categories: dominant, co-dominant and supressed (Jennings, Brown, & Sheil, 1999). Dominant tree crowns extended over the canopy, receiving full sunlight on the top of the crown and partial sunlight on the sides. Co-dominant trees form the general canopy, receiving full sunlight on top but minimal light at the sides of the crown. Suppressed trees are positioned below the general canopy, receiving no direct sunlight on either the top or sides of the crown; some juvenile trees may have been included in the latter category.
On each tree, a steel band-dendrometer (Liming, 1957) was fixed to the trunk at height 1.30 m to measure growth circumference with digital callipers at 0 months, 12 months and 24 months. In temperate species, growth studies measure annual rings or use stem analysis, but most tropical species lack annual growth rings, and therefore permanent plots and periodic measurements of diameter and height are used. Given that it remains to be evaluated whether most TMCF tree species in the study region form annual rings, we used band dendrometers, which are reported as one of the most accurate methods for detecting even slight growth (Baker, Affum-Baffoe, Burslem, & Swaine, 2002; Sheil, 2003).
The basal area of neighbouring trees was used as a surrogate for competition (Biging & Dobbertin, 1992). For this, the basal area of all the trees surrounding each individual study tree was estimated using a Cruz-All basal area forest stand gauge (Jim-Gem®) (Kerhoulas, Kolb, & Koch, 2013). The number of neighbouring trees around the measured tree was quantified (using the English basal area factor 5) and then multiplied by 1.148, as proposed by Larsen (2007), to obtain the total basal area (m2 ha-1) of neighbouring trees. The factor 5 was used given the low tree density and predominance of small trees at the study sites (Avery & Burkhart, 1983).
Data analysis
A general linear model (GLM) was used to evaluate the fixed effects of initial tree diameter (covariate), neighbouring tree basal area (covariate), crown class (factor with three levels), and site (factor with two levels) on the diameter growth rate of each species. Site was considered as a fixed factor based on the recommendation by Bolker (2015). Model fit was assessed by evaluating residual plots, which presented in all cases a distribution that was close to normal. The best model was selected based on the Akaike information criterion (AIC). All statistical analyses were run in R, version 3.3.3 (R Core Team 2017, https://www.R-project.org/). The mean growth values calculated from two years of measurements were used.
Results
Overall, the highest diameter growth rate was recorded for Acuminate (mean ± SE; 1.62 cm yr-1 ± 0.08 cm yr-1), followed by Q. xalapensis (0.91 cm yr-1 ± 0.07 cm yr-1) and L. styraciflua (0.71 cm yr-1 ± 0.08 cm yr-1). Based on the AIC, initial tree size (dbh) was removed from the models explaining the growth of Acuminate and Q. xalapensis, and in the case of L. styraciflua, diameter growth decreased with tree size (Table 2; Fig. 1). Crown class had a significant effect on diameter growth rates; dominant trees had higher growth rates than supressed trees in all three species studied (Table 2; Fig. 2). The analyses showed that the neighbouring tree basal area values have a strong influence on tree growth of the species studied; in these secondary forests with no management, diameter growth rates decreased with increasing neighbouring tree basal area (Fig. 3). Growth of Acuminate differed between stands, with a higher growth rate recorded in stand 1 than in stand 2 (Table 2).
Table 2 General linear model for the effect of tree size, crown class (dominant, co-dominant and supressed), neighbouring tree basal area and stand on the diameter growth rates of cloud forest tree species.
| Alnus acuminata | Liquidambar styraciflua | Quercus xalapensis | ||||
| Coefficient | P | Coefficient | P | Coefficient | P | |
| R 2 | 0.462 | 0.465 | 0.322 | |||
| Intercept | 2.24 ± 0.161 | <0.0001 | 1.981 ± 0.228 | <0.0001 | 1.536 ± 0.169 | <0.0001 |
| DBH | -0.018 | 0.014 | ||||
| Crown class 2 | 0.11 ± 0.128 | 0.38 | -0.304 ± 0.134 | 0.028 | -0.225 ± 0.128 | 0.085 |
| Crown class 3 | -0.48 ± 0.203 | 0.021 | -0.517 ± 0.194 | 0.010 | -0.672 ± 0.179 | 0.0004 |
| BA | -0.040 ± 0.014 | 0.008 | -0.041 ± 0.009 | <0.0001 | -0.031 ± 0.012 | 0.014 |
| Site | -0.510 ± 0.119 | <0.0001 | ||||
DBH = diameter at breast height, BA = neighbouring tree basal area. B = Coefficient ± SE, P = probability of Type I error.
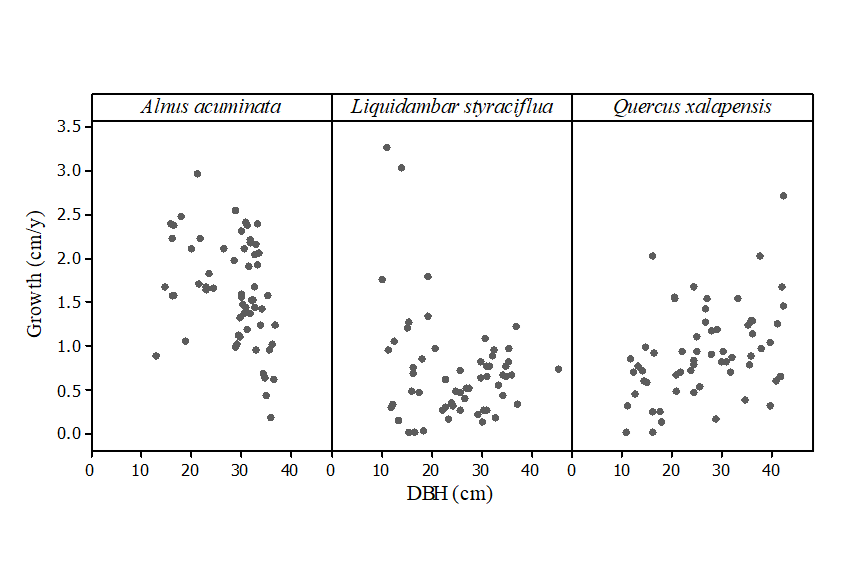
Figure 1 Diameter growth rate (cm yr-1) and tree diameter at breast height (DBH; cm) in tropical montane cloud forest tree species (N = 60). Diameter growth decreased with DBH in Liquidambar (P = 0.014; see table 2 for the full model).
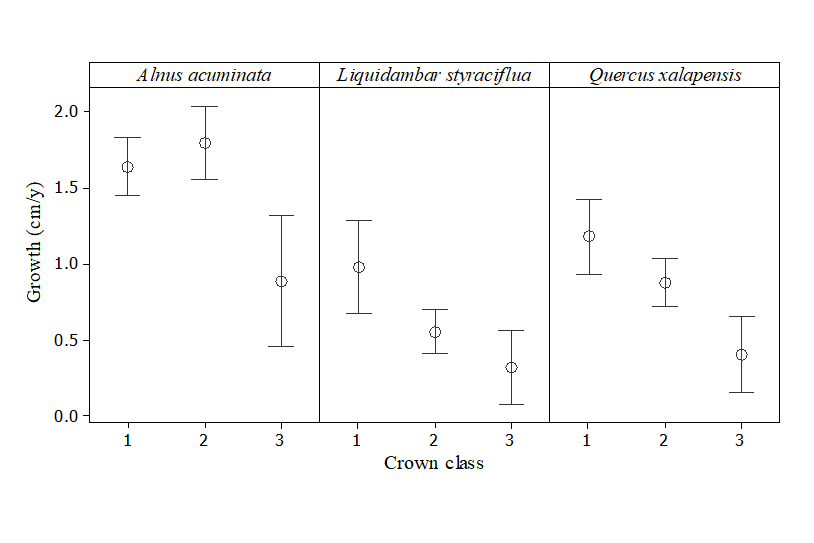
Figure 2 Diameter growth rates per crown class in tropical montane cloud forest tree species: 1 = dominant, 2 = co-dominant and 3= supressed (N = 20). Growth rate was higher in dominant trees than in suppressed trees in all three study species (P < 0.05; see table 2 for the full model).

Figure 3 Relationship between diameter growth rate (cm yr-1) and the neighbouring tree basal area (m2 ha-1) in tropical montane cloud forest tree species (N = 60). Diameter growth decreased with increasing neighbouring tree basal area in all three study species (P < 0.05; see table 2 for the full model).
Discussion
At present, secondary and degraded forests cover a larger area than primary forest in tropical landscapes (Chazdon, 2003; Food and Agricultural Organization [FAO], 2015). In Mexico a steady expansion of secondary forests has been reported (Rosete-Vergés et al., 2014). Their role in the conservation of biodiversity and provision of timber and non-timber products has been the subject of increasing attention (Finegan, 1992; Chazdon et al., 2009). The estimated growth rates of the species studied demonstrate their potential for inclusion in management programs for timber production in secondary TMCF. The mean growth rate found for A. acuminata is very high, considering that the study was conducted in forest sites with no management, and is similar to the growth rate of 1.725 cm yr-1 reported for a monospecific plantation of this species in Costa Rica (Centro Agronómico Tropical de Investigación y Enseñanza [CATIE], 1995). By comparison, Pinus patula was reported to have a growth rate of 1.03 cm yr-1 in managed forests in the studied region (Alvarez, 2001). In the study region, forest owners are encouraged to replace areas of secondary TMCF with P. patula plantations as part of reforestation programs (Paré & Gerez, 2012). This pine species, endemic to Mexico, is the most popular species for reforestation programs in humid montane forests and the establishment of commercial forest plantations in Latin America (Sáenz-Romero, Snively, & Lindig-Cisneros, 2003; Wright, DiNicola, & Gaitan, 2000). Our results show that the TMCF tree species studied, such as A. acuminata, can display similar or even superior growth rates to those of Pinus species in managed forests in the same region (Álvarez, 2001).
In comparison to the growth rate reported in a protected area in Xalapa, Veracruz, Mexico, the mean growth rate found was lower for L. styraciflua and higher for Q. xalapensis (1.47 cm yr-1 and 0.43 cm yr-1, respectively; Williams-Linera, 1996). Given the fact that both crown position and basal area of neighbours had significant influence on growth rates, comparisons are difficult since these variables were not assessed in the study of Williams-Linera (1996). Given the limited studies of TMCF, we compared our results with data reported for a lowland secondary forest in Puerto Rico and found that the growth rates measured in our study are considerably higher (0.21 cm y-1 reported in Weaver & Birdsey, 1990). However, it should be considered that tree growth rates in secondary forests are highly variable, with important differences between species, stand ages, densities and forest types (Brown & Lugo, 1990; Akindele & Onyekwelu, 2011).
Initial tree size had a significant effect on growth rate in L. styraciflua; growth rates were slower in bigger trees, a common pattern in tropical tree species (Lamprecht, 1989). In agreement with our findings, Clark and Clark (1999) concluded that the diameter growth pattern is highly dependent on tree size. This information could be useful in determining cutting cycles for these species in secondary TMCF. The small contribution of tree size to explaining the variation in growth in the other species was probably due to the important role played by competition in the unmanaged forests studied. Since increased tree growth occurs as a result of reduced competition via harvesting (Burkhart & Tomé, 2012), selective cutting as part of silvicultural treatments can be expected to increase growth rates. Previous studies in Costa Rican montane forests report a significant increase in the growth rate of juvenile (Saenz & Guariguata, 2001) and adult (aus der Beek & Saenz, 1996) trees, following timber harvesting intensity of 20% to 30% of stand basal area. Evaluating the effects of silvicultural experiments, such as liberation thinning to release future crop trees and increase their growth rates, would provide valuable information with which to advance the management of secondary TMCF.
Since band-dendrometers are sensitive to millimetric fluctuations in diameter, measurements can be affected by differences in both timing and climate (Sheil, 2003). Thus, measurements taken over longer periods of time would be required to assess the influence of environmental factors with confidence. The only species that displayed any differences between sites was A. acuminata, the higher growth rate in site 1 could be associated with its location at an elevation 500 m lower than that of site 2, reflecting the plasticity of this species in response to changes in environmental factors. Warmer temperatures at lower elevations have been reported to enhance tree growth (Way & Oren, 2010).
Promoting local management strategies that integrate the economic needs of forest owners with diverse alternatives and intensities of silvicultural treatments is fundamental to the conservation and sustainable management of TMCF at landscape level (Toledo-Aceves et al., 2011). The study species could provide timber, firewood and charcoal to meet local demand if planned management schemes for secondary forest are applied. Quercus species in general are preferred for the production of firewood and charcoal (Haeckel, 2006), fuelwood sources that play an important role in the energy security of rural communities and, more recently, meet an increased urban demand due to use in restaurants (Mwampamba, Ghilardi, Sander, & Chaix, 2013). Alnus acuminata is valued for the construction of furniture due to its moderately light wood and excellent workability (Ruso, 1994). It has also been shown to be useful for soil improvement since it is a nitrogen-fixing species (Ngom et al., 2016). Furthermore, the shade created by A. acuminata plantations could assist in the suppression of the bracken fern Pteridium aquilinum, an invasive species that can arrest secondary succession in degraded soils (Avendaño-Yañez, Sánchez-Velásquez, Meave, & Pineda-López, 2014). Overall, our results show that the studied species have potential for the diversified management of TMCF, which could have a positive effect on both biodiversity conservation and the local economy.
Conclusions
Based on the high growth rates recorded, our results support that the species studied have great potential for timber, firewood and charcoal production in secondary TMCF forests. The negative influence of the basal area of neighbouring trees on growth rates indicates that the application of silvicultural treatments to reduce competition may serve to increase the productivity of the tree species studied. The design of appropriate management schemes for harvesting secondary TMCF could enable the conservation of these species, while contributing to the maintenance of these forests and local livelihoods.

