











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
INTRODUCCIÓN
Los humedales son ecosistemas muy importantes en México, dentro de estos, se encuentran los manglares, los cuales se distinguen por su riqueza natural y los servicios ambientales que sustentan (Agraz-Hernández et al., 2009; Rodríguez-Zúñiga et al., 2013). En los manglares dominan distintas especies arbóreas, que son conocidas como mangles (Rodríguez-Zúñiga, 2013). Las diferentes especies del manglar se pueden agrupar y distribuir de diferentes maneras dentro de su hábitat. Así, se pueden distinguir cinco tipos ecológicos o fisiográficos de manglar según su ubicación: de franja, de cuenca, petén, chaparro y ribereño (Lugo y Snedaker, 1974; Cintrón et al., 1985; Herrera-Silveira y Comín, 2000; Zaldívar-Jiménez et al., 2010). El manglar de tipo ribereño o riverino, se desarrolla en las orillas de los ríos, así como en las zonas donde hay influencia de las mareas y agua salobre (estuarios), frecuentemente se encuentra dominado por Rizophora sp. y en algunas ocasiones por otras especies de mangles, y en donde comúnmente se presentan altas descargas de agua dulce y nutrientes, originando bosques de características estructurales bien desarrolladas (Moreno-Casasola e Infante-Mata, 2016).
Por otro lado, las adaptaciones de las especies que conforman los manglares les permiten sobrevivir en un amplio rango de inundaciones y salinidad, proporcionando múltiples servicios socioambientales de gran valor para el mantenimiento del ecosistema y de los procesos naturales (Agraz-Hernández et al., 2006). El manglar es un hábitat importante para una gran diversidad de especies, en el sotobosque se pueden encontrar algunos helechos y pastos, y en la zona de las raíces una gran diversidad de fauna acuática en sus diferentes etapas de vida, funcionando como guardería de los estadios juveniles. Además, la estructura vertical de los manglares funciona como descanso y anidación de diferentes especies de aves tanto locales como migratorias y, en donde pueden establecerse diferentes especies de plantas trepadoras y epífitas (Rodríguez-Zúñiga et al., 2013; Moreno-Casasola e Infante-Mata, 2016); como aráceas, helechos y orquídeas y, en el Netrópico, las bromelias y cactáceas (Zotz, 2013; Hogart, 2015; Jiménez-López et al., 2018).
Las epífitas son plantas que tienen la capacidad de crecer sobre árboles hospederos durante todo su ciclo de vida, sin obtener sus nutrientes (Bautista-Bello et al. 2021); no son especies parásitas, y únicamente utilizan la estructura vertical de los forófitos para sostenerse del tronco y las ramas (Benzing, 2008). Sin embargo, en algunos casos muy específicos, epífitas como Tillandsia recurvata con elevado éxito reproductivo en zonas semiáridas pueden causar la muerte de las ramas de sus hospederos debido a que pueden inducir cambios estructurales y ecofisiológicos (Pérez-Noyola et al., 2021). De acuerdo con Zotz y Andrade (2002), las epífitas pueden subdividirse en (1) holoepífitas, que son epífitas verdaderas y cumplen todo su ciclo de vida en un hospedero; y (2) en hemiepífitas las cuales se comportan como epífitas en el primer o último ciclo de vida.
Las epífitas tienen una importante función en los ecosistemas, ya que, según su distribución vertical pueden servir como alimento, hábitat y nicho para otros organismos como hormigas, artrópodos, insectos, aves y mamíferos (Cestari y Pizo, 2008; Hurtado-Alza et al., 2017; Aguilar-Rodríguez et al., 2019; Vargas-Castro y Newcombe, 2019; Reyes-García et al., 2022; Rocha, 2023).
Las características de los forófitos, como la estructura de la corteza, también podrían influir en la distribución vertical de las epífitas (Wyse y Burns, 2011; Sáyago et al., 2013; Wagner et al., 2015). Las cortezas ásperas y fisuradas tienen gran capacidad para acumular nutrientes en forma de detritos y capturar agua, utilizando estos micrositios para su fijación (Cascante-Marín et al., 2009; Reyes et al., 2010; Zytynska et al., 2011).
En general, las especies de epífitas vasculares asociadas a los árboles del manglar, son las mismas que se podrían encontrar en las comunidades terrestres adyacentes. No pueden tolerar niveles altos de sal y, por lo tanto, no entran profundamente en el hábitat de los manglares. Sin embargo, se pueden encontrar excepciones, por ejemplo, algunas bromelias como Tillansya dasyliriifolia, presentan hojas suculentas y parecen acumular sal dentro de sus tejidos. Esto sugiere que han desarrollado un grado de tolerancia a la sal en forma paralela con los árboles del manglar en los que se desarrollan (Gómez y Winkler, 1991).
El éxito en la supervivencia de las epífitas está directamente relacionado con su capacidad para adaptarse a un entorno aéreo, debido a sus modificaciones morfofisiológicas y con las condiciones de su hospedero, por lo que presentan preferencias por ciertas partes del tronco o rama del forófito (Benzing, 1990; Zimmerman y Olmsted, 1992; Krömer et al., 2007b). Entre los factores ambientales que determinan la distribución de las epífitas en los diferentes tipos de vegetación y dentro de su hospedero, el agua es el más importante. Sin embargo, se deben tomar en cuenta otros factores que pueden limitar el establecimiento y la supervivencia de las epífitas; como la baja disponibilidad de nutrientes, su capacidad fotosintética, el tamaño del sustrato y fenología del forófito, la presencia de otras epífitas, interacciones con otros organismos, vientos fuertes, la morfología y el tamaño de los individuos (Castro et al., 1999; Benzing, 2000; Graham y Andrade, 2004; Cervantes et al., 2005), así como su vulnerabilidad ante el cambio de uso de suelo, el cambio climático y las perturbaciones antropogénicas (Köster et al., 2009; Krömer et al., 2014; Males et al., 2023).
La distribución vertical y colonización de las epífitas se ha estudiado en diferentes tipos de vegetación y de condiciones ambientales, aportando importante información sobre su composición florística y su contribución hacia la flora total (Krömer et al., 2007a; Mora-Olivo et al., 2018), aunque esto último podría complicarse debido principalmente a la dificultad que implica su colecta (Ingram y Lowman, 1995; Krömer et al., 2007b).
En cuanto a los estudios sobre diversidad y abundancia de epífitas en los ecosistemas de manglar, se han hecho diferentes esfuerzos para generar información con inventarios y estudios ecológicos, sin embargo, estos siguen siendo escasos (Rahman et al., 2015; Sousa y Colpo, 2017; Kupec, 2018; Rohani et al., 2020; Noguera-Savelli et al., 2021).
Algunos manglares, como los de Asia, presentan una alta diversidad de epífitas vasculares (Giesen et al., 2007; Rahman et al., 2015). Aunque, también podemos encontrar que muchos ecosistemas de manglar pueden presentar alta densidad de epífitas vasculares pero baja diversidad de especies (Olmsted y Gómez-Juárez, 1996; Rico-Gray y Palacios-Ríos, 1996; Cach-Pérez et al., 2013). Esta composición y distribución espacial de las epífitas podrían estar relacionada con las características fisonómicas del forófito (e.g. corteza y arquitectura); con la altura, las zonas de un mismo forófito y las características microambientales que en los bosques de manglar pueden ser relativamente extremas, como luz, nutrientes, temperatura, sequía, exposición a aerosoles y salinidad (Gómez y Winkler, 1991; Wanek y Zotz, 2011; Cach-Pérez et al., 2013; Stein et al., 2014; Wagner et al., 2015; Mikolaev et al., 2016; Tomlinson, 2016; Jiménez-López et al., 2017; Rioja-Nieto et al., 2017; Sousa y Colpo, 2017; Rohani et al., 2020); así como con la producción de metabolitos secundarios derivados de los árboles que habitan en los bosques de manglar; como alcaloides, fenoles, esteroides, terpenoides y taninos (Boto, 1982; Bandaranayake, 1998; Marroquín y Cruz, 2016); entre otros factores y características de origen mecánico como la influencia de olas, mareas y corrientes (Gómez y Winkler, 1991; Zotz y Reuter, 2009; Rodríguez-Zúñiga et al., 2013).
La información que proporcionan los estudios sobre la riqueza y diversidad de especies es primordial para la conservación de áreas de excepcional valor biótico, así como para la mitigación de los problemas ambientales o de salud del ecosistema. El manglar ribereño de la comunidad de El Palmar, Quintana Roo, es un ecosistema de gran riqueza biológica y escénica que ha sido poco estudiado, y se encuentra en peligro debido a las actividades humanas como el uso recreativo para el turismo, la construcción de vialidades y la deforestación del manglar para actividades agrícolas como la caña de azúcar, las cuales han provocado la interrupción del flujo hidrológico (Magnon, 2002), además de poner en peligro a las especies que lo habitan como las epífitas, ya que son extraídas para fines ornamentales, principalmente las orquídeas (Ibarra et al., 2002). Así, el presente trabajo pretende contribuir a la identificación y conocimiento de la composición de epífitas vasculares del manglar ribereño, y promover la importancia de los manglares para su conservación en la comunidad de El Palmar, Quintana Roo, México.
OBJETIVOS
Determinar la composición florística y distribución vertical de las epífitas vasculares asociadas a los forófitos del manglar ribereño en la comunidad de Ramonal, Mpio. de Othón P. Blanco, del estado de Quintana Roo, México.
MATERIALES Y MÉTODOS
Área de estudio
Este trabajo se llevó a cabo en el bosque de manglar en la ribera del río Hondo, al suroeste del estado de Quintana Roo, México, en la comunidad de El Palmar en el ejido del Ramonal (N-88.531667, W18.419167, 30 m s.n.m.; Fig. 1) El río Hondo nace en Guatemala y desemboca en la bahía de Chetumal. Este sistema hidrológico es un importante corredor biológico entre el sur de la península de Yucatán con Belice y Guatemala (Arriaga et al., 2000); integrado por una amplia red de humedales compuesto de lagunas, manglares, pantanos, llanuras de inundación y cenotes (Magnon, 2002). Tiene una longitud de 145 km, un ancho promedio de 50 m, una profundidad media de 10 m y su cuenca tributaria tiene un área de 13 465 km2 (Herrera-Sansores y Heredia-Escobedo, 2011). Esta zona presenta un clima cálido subhúmedo con lluvias en verano, la temperatura promedio anual es de 26 °C y la precipitación total anual es de 1500 mm la cual ocurre principalmente entre mayo y noviembre (Sánchez-Sánchez e Islebe, 1999; Arriaga et al., 2000).

Figura 1 Mapa del sitio de muestreo en el manglar ribereño de la comunidad de Ramonal, en el Río Hondo, Mpio. de Othón P. Blanco, en el estado de Quintana Roo, México.
La vegetación ribereña del río Hondo incluye manglares compuestos por Rhizophora mangle (orilla del río), Avicennia germinans, Conocarpus erectus y Laguncularia racemosa (detrás de la franja de R. mangle) (Torrescano e Islebe, 2006), así como elementos de la vegetación aledaña como la selva mediana subcaducifolia, selva baja perennifolia, selva baja inundable, sabana, cañaveral y pastizal cultivado (Arriaga-Cabrera et al., 1998; Sánchez-Sánchez e Islebe, 2002).
Diseño de muestreo y medición de variables
El trabajo de campo se llevó a cabo entre febrero a noviembre de 2020. Se eligieron cinco sitios de muestreo en el manglar ribereño, y debido a que el bosque de manglar es angosto en esta zona (aprox. 4 m - 16 m de ancho) en cada sitio se hizo un transecto de 100 m de longitud, paralelo al río, con cuatro parcelas circulares de 7 m de diámetro cada una, a una distancia de 20 m entre ellas, cubriendo una superficie total muestreada de 769.3 m2. La ubicación de cada transecto se registró con un GPS.
En cada parcela se identificó cada uno de los forófitos y se registró la altura con una vara telescópica de 15 m de alcance, así como el diámetro del tronco a la altura del pecho (DAP), se registraron dos mediciones del perímetro del tronco principal a 30 cm sobre las raíces aéreas con una cinta métrica de 1 m de alcance (Hechavarria y Ferro, 2017; Rodríguez et al., 2018) y se calculó el diámetro normalizado (diámetro = perímetro/π). Cada forófito fue etiquetado con una placa de aluminio representando su parcela, transecto y el número de forófito muestreado, para corroborar datos si se requiere. Por otro lado, para determinar el índice de valor de importancia (IVI) de los forófitos se utilizaron las fórmulas propuestas por Aguirre y Aguirre (1999).
Para la identificación, localización, medición y conteo de las especies epífitas y hemiepífitas (Bautista-Bello et al., 2021; Zotz y Andrade, 2002) se utilizó el método de observación directa y a distancia (Johansson, 1974; Ferro, 2015), y la técnica de arbolero (Wendt, 1986), en donde para el ascenso al dosel se contó con personas con habilidad para trepar. Para localizar la estratificación vertical se utilizó la zonación propuesta por Johansson (1974). De cada individuo se midió el tamaño y la altura con un flexómetro, y con una vara telescópica se registró la altura a la que se encontró cada epífita sobre el forófito desde la base del tronco hasta la parte superior del dosel para cada uno de los forófitos por parcela, incluyendo las raíces aéreas de Rhizophora mangle. En el muestreo se incluyeron los adultos y juveniles (>5 cm). Para estimar la abundancia de las epífitas, se tomaron como individuos las rosetas (bromelias), laminas (aráceas) y grandes agregados (orquídeas). Se tomaron fotografías digitales de las epífitas vasculares agregando información de su ubicación y un número de inventario. Para la identificación taxonómica se utilizaron guías, artículos y catálogos de la Flora de Quintana Roo y la Península de Yucatán (Ramírez et al., 2004; Mondragón-Chaparro et al., 2011; Campos et al., 2020; Noguera-Savelli et al., 2021).
Para determinar la diversidad se calculó con el índice de Shannon-Wiener y el índice de Simpson (Aguirre-Mendoza, 2013). El índice H´ tiene tres rangos de valores: 0-1.35, 1.36-3.5 y >3.5, indican baja, media y alta diversidad, respectivamente. En el caso del índice de Simpson el rango es: 0-0.33, 0.34-0.66 y >0.67, indican media, baja y alta, respectivamente.
El área basal de los forófitos se obtuvo en función del diámetro a la altura del pecho (DAP) del árbol, ésta se calcula mediante las fórmulas propuestas en Aguirre-Mendoza (2013) y Rodríguez et al. (2018).
La altura y el DAP de los forófitos son las variables más utilizadas en estudios de epífitas, y se ha encontrado que a mayor altura y DAP, hay mayor riqueza de epífitas vasculares (Flores-Palacios y García-Franco, 2006; Haro-Carrión et al., 2009; Wolf et al., 2009), de esta forma para determinar si existía una relación entre el número de especies y la estructura del forófito (altura y DAP) se hizo un análisis de regresión lineal simple utilizando el software Origin 8. Para jerarquizar la dominancia de cada especie arbórea presente en el sitio de estudio, se obtuvieron los datos de las variables estructurales como abundancia, dominancia, cobertura y frecuencia, para sintetizar la jerarquía de cada especie en un número adimensional y refleja su importancia en la comunidad. Finalmente, se hizo una prueba de Kruskal-Wallis para determinar diferencias significativas entre las alturas a las cuales se encontraron las especies de epífitas con el software STATISTICA 8.
RESULTADOS
En total se registraron 60 individuos de árboles, pertenecientes a seis especies distribuidas en seis familias: Rhizophoraceae, Combretaceae, Sapotaceae, Bombacaceae, Arecaceae y Anacardiaceae; de estas, sólo en cuatro especies se registraron epífitas: Rizophora mangle, Bucida buceras, Pachira acuatica y Chrysophyllum cainito. Las especies arbóreas se ordenaron en función de mayor a menor IVI, y se calculó la densidad, frecuencia y dominancia relativa (Tabla 1), así R. mangle y Bucida buceras fueron los forófitos con mayor IVI. De las especies arbóreas, R. mangle fue la especie con mayor abundancia de epífitas (77%), seguido de Chrysophyllum cainito (12%, Fig. 2).
Tabla 1 Índice de valor de importancia (IVI), de las especies arbóreas que componen el manglar ribereño en la comunidad de El Palmar, en el río Hondo.
| Familia | Especie | Forófito | Área basal (m2) | Drel (%) | Frel (%) | Domrel (%) | IVI |
| Rhizophoraceae | Rizophora mangle (L.) C.DC. | X | 131.65 | 50 | 42.5 | 58.92 | 50.47 |
| Combretaceae | Bucida bucerasL. | X | 64.43 | 21.67 | 27.5 | 28.83 | 26.00 |
| Sapotaceae | Chrysophyllum cainito L. | X | 15.9 | 11.67 | 15 | 3.69 | 10.12 |
| Bombacaceae | Pachira acuatica Aubl. | X | 8.25 | 10.00 | 7.5 | 7.12 | 8.21 |
| Arecaceae | Acoelorraphe wrightii (Griseb. & H. Wendl.) H. Wendl. ex Becc. | 2.52 | 5.00 | 5.00 | 1.13 | 3.71 | |
| Anacardiaceae | Metopium brownei (Jacq.) Urb. | 0.68 | 1.67 | 2.50 | 0.31 | 1.49 | |
| Total | 223.44 | 100 | 100 | 100 | 100 |
Densidad relativa (Drel), frecuencia relativa (Frel), dominancia relativa (Domrel)
Las especies con función de forófito se indican con una X.

Figura 2 Abundancia absoluta (%) de epífitas vasculares encontradas sobre los forófitos del sitio de estudio.
Se registraron nueve especies de epífitas y dos hemiepífitas, distribuidas en cuatro familias y ocho géneros. Las tres especies de la familia Bromeliaceae representan 76% del total de individuos encontrados, seguida de Orchidaceae (5 spp.; 18%), Cactaceae (1 spp.; 5%) y, por último, Araceae (2 spp.; 1%). En total se registraron 335 individuos de epífitas, correspondientes principalmente a tres especies que presentaron la mayor abundancia: Tillandsia dasyliriifolia (Bromeliaceae, 59%), T. streptophilla (Bromeliaceae, 16%) y Cohniella ascendens (Orchidaceae, 10%, Fig. 3).
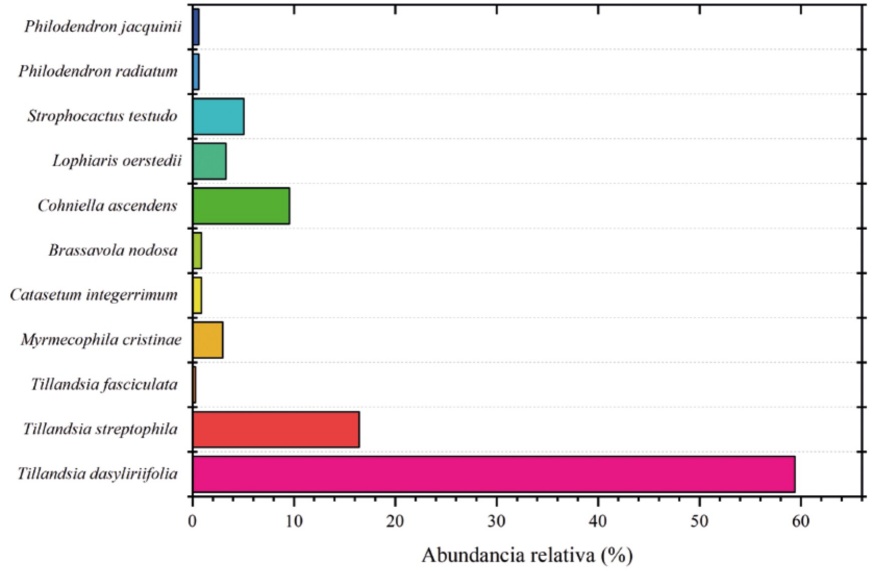
Figura 3 Abundancia relativa (% de individuos) de las especies epífitas registradas en el sitio de estudio.
La zona con mayor colonización de epífitas fueron la zona I y II, que corresponde a los tallos y raíces a una altura promedio de 2.36 m ± 0.054 m, y se encontraron diferencias significativas en la altura de colonización entre especies (Kruskal-Wallis: H (10, N = 335) = 21.30; p = 0.019, Fig. 4); en donde T. fasciculata fue la especie que se encontró a mayor altura (6.005 m ± 0.005 m) y Catasetum integerrimum a menor altura (0.833 m ± 0.283 m).

El modelo de regresión lineal simple sugiere que no existe relación entre la altura y DAP de los forófitos con la riqueza de epífitas (altura: a = 0.977, b = 0.0403, r2 = 0.0177, p > 0.05; DAP: a = 0.760, b = 0.0321, r2 = 0.0478, p > 0.05).
En la tabla 2 se muestra la abundancia, el índice de diversidad de Shannon-Wiener (H´) y el índice de diversidad de Simpson (D) de las especies de epífitas encontradas en el sitio de estudio, en donde los valores de los índices de H´ = 1.36 y D = 0.61 sugieren una diversidad media de epífitas vasculares.
Tabla 2 Abundancia, índice de diversidad de Shannon-Wiener (H´) e índice de diversidad de Simpson (D).
| Familia | Especies epífitas | Abundancia | Índice de Shannon-Wiener | Índice de Simpson (D) | ||
| (H´) | pi (n/N) | (pi)2 | ||||
| Bromeliaceae | Tillandsia dasyliriifolia Baker. | 199 | 0.31 | 0.594 | 0.353 | |
| Tillandsia streptophilla Scheidw. ex E. Morren. | 55 | 0.3 | 0.164 | 0.027 | ||
| Tillandsia fasciculata S.w. | 1 | 0.02 | 0.003 | 0 | ||
| Orchidaceae | Myrmecophila christinae Carnevali & Gómez-Juárez var. christinae | 10 | 0.1 | 0.03 | 0.001 | |
| Catasetum integerrimum Hook. | 3 | 0.04 | 0.009 | 0 | ||
| Brassavola nodosa (L.) Lindl. | 3 | 0.04 | 0.009 | 0 | ||
| Cohniella ascendens (Lindl.) Christenson. | 32 | 0.22 | 0.096 | 0.009 | ||
| Lophiaris oerstedii (Rchb. f.) R. Jiménez, Carnevali & Dressler | 11 | 0.11 | 0.033 | 0.001 | ||
| Cactaceae | Strophocactus testudo (Karw. ex Zucc.) Bauer | 17 | 0.15 | 0.051 | 0.003 | |
| Araceae | Philodendron radiatum Schott.* | 2 | 0.03 | 0.006 | 0 | |
| Philodendron jacquini Schott.* | 2 | 0.03 | 0.006 | 0 | ||
| 335 | 1.36 | 0.394 | ||||
| 0.606 | ||||||
Las proporciones de las especies se indican como pi, en donde pi indica la abundancia de la especie i con respecto a la abundancia total de las especies epífitas en el sitio de estudio. *hemiepífitas
DISCUSIÓN
Las especies arbóreas del bosque de manglar ribereño de El Palmar, Quintana Roo, están representadas principalmente por Rhizophora mangle, y por cinco especies que provienen de las formaciones vegetales vecinas, que manifiestan un comportamiento facultativo en su asociación con el manglar (Sutton et al., 1983); y de las cuales cuatro especies funcionan como forófitos (R. mangle, Bucida buceras, Pachira acuática y Chrysophyllum cainito). El éxito de las epífitas está relacionado con las características y condiciones de su forófito, se han sugerido zonas particulares de establecimiento, con tendencias por ciertas zonas del tronco o ramas del forófito (Benzing, 1990; Zimmerman y Olmsted, 1992; Krömer et al., 2007b), al tamaño, edad, calidad de las ramas y textura de la corteza (Andersohn, 2004; González y Ceballos, 2021), así como con la estructura del dosel, el cual puede modificar el microclima (Susan-Tepetlan et al., 2015).
Bajo ciertos ambientes se ha registrado una relación entre el DAP y la altura de los forófitos con la riqueza de especies epífitas (Flores-Palacios y García-Franco, 2006; 2008). Sin embargo, Cach-Pérez et al. (2013) no encontraron una relación entre la altura y la abundancia de bromelias en manglares de Yucatán; esto podría deberse a la baja estatura de los individuos de R. mangle, los cuales miden 1.5m. De la misma forma, en el manglar ribereño de El Palmar no se encontró una relación entre la altura y el DAP de los forófitos con la riqueza de epífitas. Estos datos sugieren que, a pesar de que los árboles son altos (hasta 9 m), las epífitas y hemiepífitas colonizan preferentemente las zonas I y II de los forófitos (2.5 m; Johansson, 1974) que corresponden a los troncos y raíces de los árboles de R. mangle, que además de presentar el mayor IVI, su corteza rugosa podría facilitar el establecimiento de las epífitas; además de disponer de mayor humedad debido a su cercanía al suelo inundado proporcionando un microclima adecuado para la germinación y anclaje de las semillas. También, la distribución y abundancia de las epífitas podría estar influenciada por las características de los forófitos (Krömer et al., 2007b), y por las raíces aéreas, como un nicho adicional para la colonización de epífitas (Jiménez-López et al., 2017).
Esta distribución se ha observado en diferentes ecosistemas, en donde las epífitas frecuentemente colonizan la altura baja y media de los forófitos, y la mayor abundancia de los individuos podría estar relacionada con menor intensidad de luz y temperatura, así como mayor humedad, más que del forófito o la ubicación de su hábitat (Benzing, 1990; Castro et al., 1999; Zotz y Hietz, 2001; Krömer et al., 2007b; Noguera-Savelli et al., 2021; Flores-Argüelles et al., 2022), respondiendo a una estrategia de máximo aprovechamiento de recursos como agua, nutrientes y materia orgánica.
Diferentes estudios han sugerido que las epífitas vasculares son poco comunes en los bosques de manglar (Valdez-Hernández, 2002; Cach-Pérez et al., 2013; Rahman et al., 2015; Jiménez-López et al., 2017; 2018; Sousa y Colpo, 2017; Kupec, 2018; Rohani et al., 2020; Noguera-Savelli et al., 2021); sin embargo, también se ha encontrado que estas plantas pueden ser frecuentes y abundantes (Carmona-Díaz et al., 2014; Jiménez-López et al., 2017; 2018); registrando especies de bromelias, orquídeas y algunas aráceas (Gentry y Dodson, 1987; Moreno-Casasola e Infante Mata, 2009; Tomlinson, 2016).
Para el manglar ribereño de El Palmar, la familia Orchidaceae está representada por cinco especies, seguida de la familia Bromeliaceae con tres especies. En este caso el número de especies coincide con las familias de epífitas vasculares de mayor importancia en el Neotrópico: Orchidaceae, Bromeliaceae y Araceae (Gentry y Dodson, 1987; Küper et al., 2004; material complementario 1); y fue similar a otros manglares de México y Centro América (Jiménez-López et al., 2017; 2018; Noguera-Savelli et al., 2021; ver material complementario 1).
La familia Orchidaceae, aunque presenta mayor número de especies (cinco), la abundancia de individuos es baja. Catasetum integerrimum y Brassavola nodosa son especies de orquídeas con una baja representación en su género en comparación con el grupo de las bromelias. La abundancia de la familia Orchidaceae es consistente con la literatura, en donde se señala que, si bien las orquídeas presentan mayor diversidad de especies, su principal característica es la escasez de individuos (Nieder et al., 1999).
El género Tillandsia presentó el mayor número de especies (n = 3), similar a otros estudios llevados a cabo en manglares de México y Guatemala (material complementario 1; Gómez y Winkler, 1991; Cach-Pérez et al., 2013; Jiménez-López et al., 2017). Una ventaja del género Tillandsia es que sus semillas son plumosas, permitiendo su dispersión por el viento, además algunas características de los forófitos (e.g. altura, tipo de corteza) podrían facilitar el anclaje de las semillas y proporcionar las condiciones microambientales adecuadas (García-Franco y Rico-Gray, 1988). La alta abundancia de T. dasyliriifolia podría estar relacionada con sus estrategias reproductivas (presenta sistemas de polinización como xenogamia y autogamia) y de dispersión en comparación con T. streptophilla (Mondragón-Chaparro et al., 2011) que es auto incompatible, es decir rechazan al auto-polen e impiden el cruce entre las flores de una misma planta, es solitaria y no forma grandes masas (Ramírez, 2009).
Las epífitas de cactáceas y aráceas, están representadas por una y dos especies, respectivamente; este resultado es similar a otros estudios en manglares (Valdez-Hernández, 2002; Díaz-Jiménez, 2007; Jiménez-López et al., 2017; 2018; Kupec, 2018).
En cuanto a la diversidad, Olmsted y Gómez-Juárez (1996), mencionan que las epífitas de la Península de Yucatán no son muy diversas, si se comparan con otras comunidades de epífitas de México y otros países (material complementario 1); algunos estudios han encontrado una riqueza alta, sobre todo para Tabasco y Campeche (Díaz-Jiménez, 2007; Jiménez-López et al., 2018; Noguera-Savelli et al., 2021).
Tanto el índice de Shannon-Wiener (H´ = 1.36) como el de Simpson (D = 0.61) indican una diversidad media de especies epífitas para este manglar ribereño, similar a lo reportado por Kupec (2018) y Rohani et al. (2020) con un H´ = 1.63 y 1.43, respectivamente. El total de especies epífitas registradas fue mayor, comparado con las observaciones en estudios anteriores en el sitio de estudio, en donde registraron cuatro especies de epífitas sobre R. mangle y Ficus sp. (Tovilla-Hernández et al., 2009) y, para Celestún, Yucatán en un manglar arbustivo (R. mangle y Avicennia germinans) con dos especies de epífitas (Cach-Pérez et al., 2013) y en un Petén (Cach-Pérez et al., 2018) con una especie de epífita. El haber registrado un mayor número de especies podría estar relacionado con los factores climáticos e hidrológicos y con el tipo de manglar; en donde el río Hondo tiene una fuerte influencia de agua dulce, además de un mayor esfuerzo de muestreo (Fig. 5). El mayor número de epífitas vasculares registrado para manglares en México es en los Pantanos de Centla, Tabasco, con 21 especies de epífitas vasculares sobre R. mangle (Jiménez-López et al., 2017).

Figura 5 Curva acumulada de especies de epifitas vasculares registradas para el manglar ribereño de la comunidad del Palmar, Quintana Roo.
Este trabajo es una primera línea base sobre la riqueza y abundancia de las epífitas que habitan el ecosistema de manglar ribereño en la comunidad de El Palmar en el río Hondo, Quintana Roo, por lo que se requiere continuar con proyectos que aporten más información de las interrelaciones de los hospederos y las epífitas y el ambiente que los rodea. Si bien no se registró una relación del DAP y la altura de los forófitos con la riqueza de las epífitas, los resultados apuntan a la necesidad de un muestreo más exhaustivo (Zotz y Hietz, 2001; Zotz, 2013). Al mismo tiempo se requiere de estudios del impacto antropogénico sobre la diversidad de epífitas para determinar su estado de conservación.
CONCLUSIONES
El manglar ribereño de la comunidad de El Palmar de Río Hondo presenta una diversidad media de epífitas vasculares. Las familias de epífitas vasculares mejor representadas son Orchidaceae y Bromeliaceae, similar a otros estudios para este tipo de ecosistema en el Neotrópico. Los árboles de Rizophora mangle son un refugio importante para las especies de epífitas vasculares, debido a su abundancia y sobre todo a su arquitectura en donde las zonas I y II, que corresponden a los troncos y raíces, y que por su cercanía al suelo siempre húmedo, y corteza rugosa proporcionan un microclima adecuado para su establecimiento. De esta forma, la disminución de la cobertura del manglar tendrá un impacto sobre la supervivencia y diversidad de las epífitas. La extracción con fines comerciales y ornamentales de las epífitas, así como las diversas acciones antrópicas que se llevan a cabo en la comunidad, inciden negativamente sobre la salud del manglar del río Hondo, provocando el deterioro y fragmentación del hábitat, por lo que los estudios sobre la biodiversidad del ecosistema son necesarios para establecer estrategias de conservación.

