










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPolibotánica
versión impresa ISSN 1405-2768
Polibotánica no.30 México sep. 2010
La domesticación de plantas en México: comparación de la forma cultivada y silvestre de Byrsonima crassifolia (Malpighiaceae)
Plant domestication in Mexico: comparison of the cultivated and wild forms of Byrsonima crassifolia (Malpighiaceae)
J.C. Raya-Pérez1,2, C.L. Aguirre-Mancilla1, K. Gil-Vega3 y J. Simpson3
1 División de Estudios de Posgrado e Investigación, Instituto Tecnológico de Roque, Km 8 Carretera Celaya-Juventino Rosas, CP 38110, Celaya, Guanajuato.
2 Instituto Tecnológico Superior de Uruapan, Carretera Uruapan-Carapan Núm. 5555, Col. La Basilia, CP 60015, Uruapan, Michoacán.
3 Centro de Investigación y de Estudios Avanzados del Instituto Politécnico Nacional, Unidad Irapuato (CINVESTAV-IPN U. Irapuato), Km 9.6 Libramiento Norte Carretera Irapuato-León, AP 629, CP 36821, Irapuato, Guanajuato. Correo electrónico: juancarlos.raya@gmail.com.
Recibido: 25 mayo 2009.
Aceptado: 30 abril 2010.
Resumen
Las plantas domesticadas presentan una serie de características conocidas como síndrome de domesticación. Esto permite un mejor aprovechamiento de las mismas por parte de los humanos y ha sido estudiada principalmente en plantas anuales herbáceas. En este trabajo se hizo un muestreo de árboles silvestres (changunga) y cultivados (nanche) de Byrsonima crassifolia en la región de Uruapan, México. El objetivo fue buscar diferencias genéticas mediante marcadores moleculares por medio de la técnica de AFLP entre las dos formas de la especie, encontrándose que tanto los silvestres como los cultivados están genéticamente muy cercanos entre sí. También se compararon características de los frutos como pH, dulzor de los frutos, tamaño y contenido mineral encontrándose diferencias en las tres primeras. El análisis de AFLP's agrupa juntos a las dos formas del árbol con alto grado de similitud. Fenotípicamente los frutos se distinguen entre sí, pero no a nivel molecular. La presión de selección para surtir al mercado puede poner en riesgo la diversidad presente en estas poblaciones que se propone podrían ser un modelo para la búsqueda de genes que intervienen en características como dulzor y tamaño de fruto además del proceso de domesticación en sí mismo. Se recomienda hacer colectas para los bancos de germoplasma además de tratar de promover la conservación in situ.
Palabras clave: conservación, similitud genética, tamaño de fruto, composición elemental, dulzor.
Abstract
Domesticated plants show characteristics known as domestication syndrome, which enables better utilization of the plants by humans. Plant domestication has been studied mainly in herbaceous annuals. In this work, samples of Byrsonima crassifolia trees–changunga, the wild form, and nanche, the cultivated form–were taken from the region of Uruapan, Mexico. The objective of this work was to identify genetic differences by AFLP molecular markers between these two forms. Analysis showed that both wild and cultivated forms of B. crassifolia are genetically very close to each other. Fruit characteristics of human-interest were compared, and some differences were found between the two forms, e.g., in pH, sweetness, mineral content and size. The AFLP analysis groups together the two forms of the tree with a high degree of similarity. The fruits are distinguished phenotypically from one another, but not at the molecular level. Selection pressure to supply the market may threaten the diversity present in these populations of plants, which could be a model for the screening of genes involved in traits such as sweetness and fruit size in addition to the process of domestication itself. It is recommended that collections be made for germoplasm gene banks and also that in situ conservation be promoted.
Key words: domesticated plants, Byrsonima crassifolia, conservation, genetic similarity, fruit size, elemental composition, sweetness.
INTRODUCCIÓN
México es parte del centro mesoamericano del origen de la agricultura y diversificación de plantas domesticadas (Doebley et al., 2006). Nuestro país ha aportado a la humanidad cultivos como el maíz (Zea mays), el frijol (Phaseolus vulgaris) (Zizumbo-Villarreal et al., 2005), la calabaza (Cucurbita pepo), el chile (Capsicum annum), el amaranto (Amaranthus hypochondriacus), el maguey (Agave sp.) (Gentry, 1982), el nopal (Opuntia sp.) (Hughes et al., 2007) el camote (Ipomoea batatas) (Srisuwan et al., 2006), la chía (Hyptis suaveolens, Salvia hispanica) y el nanche (Byrsonima crassifolia). El proceso de domesticación de plantas y animales, en países como el nuestro, es un proceso continuo (Hughes et al., 2007).
La domesticación implica la selección de plantas con ciertas características deseables. Estas plantas bajo cultivo y presiones de selección llegan a presentar lo que se conoce como síndrome de domesticación (Doebley et al., 2006; Zizumbo-Villarreal et al., 2005). En general, el síndrome de domesticación incluye un aumento en tamaño de los rasgos deseados, mayor suculencia, mayor tamaño, insensibilidad al fotoperiodo, eliminación de latencia en la semilla y una disminución de la capacidad de dispersión y reproducción sexual de las plantas. Por ejemplo, en arroz la capacidad o incapacidad de dispersión de la semilla es una característica clave en el proceso de domesticación (Zhongwei et al., 2007). En la berenjena (Solanum melongena), se tiene documentado que se ha seleccionado para tamaño, forma y sabor del fruto (Wang et al., 2008), características que en general han sido modificadas en las diferentes especies de plantas cultivadas. El uso de nuevas técnicas y herramientas, tal como el estudio por microscopía electrónica de los granos de almidón, de polen o de los fitolitos nos han permitido ahondar y ampliar el conocimiento sobre la domesticación de plantas, confirmando o arrojando nueva luz sobre este proceso tan importante en la historia de la humanidad (Perry et al., 2006; Piperno et al., 2007). Además, el uso que se le puede dar a este conocimiento es diverso; se han desarrollado poblaciones de jitomate (Lycopersicon esculentum = Solanum lycopersicon) resultantes de la cruza de plantas silvestres con cultivadas, llevando a la introgresión de alelos nuevos de resistencia a enfermedades y otros que influyen en las características del fruto; las variedades de alto rendimiento usadas hoy en día por la industria y la agricultura son posibles gracias a la variabilidad genética presente en los cultivos tradicionales y en las poblaciones naturales (Kovach y McCouch, 2008; Van Deynze et al., 2007). Durante el proceso de domesticación ocurre una pérdida de diversidad genética de las plantas domesticadas respecto a sus parientes silvestres. Se estima que el arroz moderno tiene entre el 10 y el 20% de la diversidad genética presente en su ancestro silvestre Oriza rufipogon (Kovach y McCouch, 2008). O. rufipogon ha sido fuente de alelos para características como tamaño y peso de grano, rendimiento, calidad, tolerancia al frío, tolerancia al aluminio y tiempo de floración (Kovach y McCouch, 2008). En el caso de la soya se estima que la domesticación redujo la diversidad genética en un 50% y eliminó el 81% de los alelos raros presentes en Glycine soja (Hyten et al., 2006). Además es posible obtener genes interesantes o características deseables a partir de estos parientes silvestres; la probabilidad de obtener variación genética novedosa es mayor si se cruzan germoplasma mejorado con silvestre (Kovach y McCouch, 2008). Cambios pequeños en la expresión de genes reguladores pueden tener efectos profundos sobre la arquitectura y el desarrollo de las plantas, en tanto que aquellos involucrados en la resistencia a enfermedades y en la reproducción y complejidad morfológica evolucionan rápidamente (Ammiraju et al., 2008; Doebley et al., 2006; Gallavotti et al., 2004; Komatsuda et al., 2007). Eventos como la poliploidia parecen haber jugado un papel muy importante en la evolución y domesticación de muchos cultivos (Hughes et al., 2007; Dubcovsky y Dvorak, 2007; Srisuwan et al., 2006). La domesticación de árboles iría detrás de la domesticación de plantas anuales, debido a que tienen un ciclo de vida más largo y existe la necesidad de una agricultura sedentaria antes del cultivo de árboles (Hughes et al., 2007).
Muchos de los árboles cultivados son propagados vegetativamente y representan una muestra de la variación genética encontrada en las poblaciones silvestres (Miller y Schaal, 2005, 2006). En las regiones donde se conserva la agricultura tradicional, huertos de traspatio y recolección de plantas es posible encontrar un continuo desde la forma silvestre hasta la forma domesticada y las variedades de elite de hoy en día. Interesantemente, en el caso de Spondias purpurea se encontró variabilidad presente en las formas cultivadas pero ausente en las silvestres (Miller y Schaal, 2005, 2006) aun cuando en este árbol reproducido vegetativamente, existe mayor variabilidad en la población silvestre (Miller y Schaal, 2006). Para Leucaena leucocephala, se detectó una menor variabilidad para su subespecie glabrata, favorecida para la producción de alimento (Hughes et al., 2007).
Haciendo uso de los AFLPs se reporta un gen de manzana que contribuye a disminuir la acidez del fruto; ésta es una característica recesiva en esta especie pero es dominante en el melocotón (Yao et al., 2007). Otros factores influyendo esta característica son la variedad, clima e, incluso, método de determinación. Se cree que hay un gen principal que determina la acidez del fruto y, por lo tanto, susceptible de modificarse más fácilmente que otras características (Yao et al., 2007). El uso de marcadores moleculares es otra de las herramientas que nos permite obtener gran cantidad de información sobre los sitios mismos de origen de los cultivos, su relación con especies silvestres y la diversidad existente en unos y otros. El proceso de domesticación de plantas y animales, en países como el nuestro, es un proceso que continúa (Hughes et al., 2007).
Las semillas de 13 especies de Leucaena se usan en México como alimento; el uso va desde la recolección para el autoconsumo en poblaciones silvestres a la cosecha intensiva para venta en los mercados regionales. Recientemente se ha reportado la existencia de formas de árboles de guaje (Leucaena sp.), en Morelos, Puebla, Veracruz, Guerrero, Oaxaca que no existen en la naturaleza. Estas formas se han originado al entrecruzarse en los pueblos, en los solares, los árboles seleccionados y plantados allí por los habitantes. Son formas de árboles que han aparecido gracias a la intervención humana. De hecho, no se han encontrado poblaciones naturales de Leucaena leucocephala (Hughes et al., 2007). Para el caso de Byrsonima crassifolia se han reportado colores de frutos rojo, amarillo, anaranjado, con un incremento en los grados brix (de 3.2 a 7.9), que en general indican mayor dulzor del fruto, a medida que se avanza en la domesticación (Martínez Moreno et al., 2006). En la región de estudio, los frutos de Byrsonima crassifolia se conocen como nanche para la forma domesticada y changunga para la forma silvestre. La fruta se vende principalmente fresca, aunque se elaboran paletas y nieves de ambas, licor de changunga y también preparada con alcohol a manera de conserva y para preparar atole. La changunga es la preferida para elaborar nieves y paletas, atole y licor, lo que resulta contrario al consumo en fresco, caso en el que se prefiere al nanche. Así pues, la siembra, cosecha, recolección, elaboración (con algún grado de industrialización) y venta de nanche y changunga resulta muy importante en la región.
Para el análisis de la diversidad genética se han desarrollado muchas técnicas, una de las más poderosas son los AFLP, que usan enzimas de restricción para fragmentar el DNA genómico. Luego se preamplifican los fragmentos y a través de amplificaciones selectivas se selecciona un subgrupo de los fragmentos; esta técnica ya ha sido utilizada para explorar la diversidad genética en plantas cultivadas (Gil-Vega et al., 2006; Yao et al., 2007). Este método es más reproducible que otros y puede amplificar transcritos de baja abundancia, además de distinguir entre genomas altamente homólogos, sin necesidad de conocer previamente la secuencia. Los marcadores se distribuyen por todo el genoma, aunque principalmente en regiones no codificantes (Meudt y Clarke, 2006). En Oryza se ha observado un reemplazo masivo de regiones intergénicas (Ammiraju et al., 2008). Usando cDNA-AFLP se aisló un gen de manzana que contribuye a disminuir la acidez del fruto (Yao et al., 2007).
MATERIAL Y MÉTODOS
Material vegetal. Se colectaron retoños de 27 árboles (16 changungas y 11 nanches) de las cercanías de Uruapan, México (cuadro 1). Los de changunga fueron colectados de El Sabino, camino a Taretan y Charapendo, las dos primeras locaciones son bosque de pino; la última es un agostadero con presencia de ganado vacuno y escasos árboles de changunga. Los de nanche se colectaron en la propia ciudad de Uruapan, en el camino a Taretan de un huerto familiar y de Charapendo de huertas dedicadas al cultivo del árbol.

AFLP. Las muestras se congelaron en nitrógeno líquido para su procesamiento en laboratorio. El ADN se obtuvo tomando 0.5 g de tejido y siguiendo el procedimiento de Edwards (2001) con modificaciones, se usó CTAB al 2.5% en el caso del nanche y al 3.5% en changunga. Los AFLP's se generaron usando cuatro combinaciones de oligos (ACT, CAC; ACT, CTC; ACT, CAT; ACT, CTA) marcados para detección con fluorescencia. Las bandas se detectaron con un secuenciador automatizado LI-COR modelo 4200. Para el análisis de relación de similitud se uso el software FreeTree. Para ello la matriz de datos de presencia/ausencia se utilizó para estimar la matriz de distancias (similitud) con el coeficiente de Sokal y Sneath 3 (S= [(a)(d)]/ [(a+b)(c+d)(a+c)(b+d)]0.5), el método de agrupamiento con promedios aritméticos no ponderados (UPGMA) para producir el dendrograma. Para evaluar su robustez, se hicieron 1000 réplicas de muestreo con reemplazo (Bootstrap) (Gil-Vega et al., 2006; Vos et al., 1995).
Análisis de frutos. Los frutos (100 de cada uno) se compraron en el mercado local de Uruapan o en Charapendo, México, los nanches que se venden en Uruapan proceden en su mayoría de Charapendo. La composición elemental se determinó mediante una sonda de rayos X (INCAxsight Oxford Instruments) acoplada a un microscopio electrónico de barrido (JEOL JMS-6480LV).
La determinación de cenizas se realizó en mufla a 600°C por 30 min; la humedad se determinó incubando las muestras en un horno a 50°C por 72 h y llevándolas a peso constante (Onder y Babaoglu, 2001), el pH se determinó con potenciómetro.
Extracción de grasas. Se utilizaron 8 g de pulpa seca de fruta extraída con hexanos en Soxhlet durante cinco horas, la grasa extraída se utilizó para correr en placa fina (TLC) usando el sistema de solventes cloroformo-metanol-agua 65-25-4, revelado con sulfato de cobre, éste se preparó usando 15.6 g de sulfato de cobre penta hidratado y se le agregó 9.4 ml de ácido fosforito al 85% en agua (p/v) y se aforó a 100 ml con agua destilada (Loyola-Martínez, 2006).
Extracción de pectina. Se utilizaron 18 g de pulpa seca, y se colocó en agua acidificada con ácido clorhídrico (pH 3), se calentó durante una hora y se dejó enfriar. Para precipitar la pectina se le agregó un volumen igual de etanol y se dejó reposar por 24 h. La pectina se obtuvo filtrando a través de papel whatman 3mm (Camejo de Aparicio et al., 1996).
Determinación de grados Brix. Se prensaron 5 g de pulpa fresca y del jugo obtenido se utilizó una gota para medir los grados Brix en un refractómetro modelo Atago 500.
Determinación de taninos. En un tubo de ensaye se colocaron 10 ml de jugo de la fruta con 2 ml de agua destilada (tubo A). En otro tubo se colocaron 10 ml de jugo de la fruta y 2 ml de solución de gelatina preparada con 3.5 g de gelatina y 10 g de NaCl en 100 ml de Agua (tubo B). Los tubos se mantuvieron a 4°C durante tres días. Posteriormente se centrifugaron y se determinaron los taninos del sobrenadante de cada uno.
Para la determinación de los taninos los sobrenadantes de los tubos A y B se diluyeron 50 veces con agua destilada. Se prepararon dos tubos y se colocaron 2 ml de sobrenadante diluido, 1 ml de agua destilada y 6 ml de HCl 12N (tubos A1, A2 y tubos B1, B2). Los tubos A1 y B1 se taparon herméticamente y se protegieron de la luz con papel de aluminio. A continuación se metieron en baño maría a 100°C durante 30 minutos. Al cabo de 30 minutos de ebullición se sacaron los tubos del baño maría y se enfriaron en agua con hielo. Los tubos A2 y B2 se dejaron a temperatura ambiente. Posteriormente, a los cuatro tubos se les añadió 1 mL de etanol y después de agitarlos se leyó la absorbencia a 550 nm (Glories, 1978). La concentración de taninos viene dada por la expresión:
Taninos condensados totales (g/L) = (D1 – D2) X 19.33
donde:
D1 = DO del tubo A1
D2 = DO del tubo A2
El valor 19.33 corresponde al coeficiente de extinción molar de la cianidina obtenida por la hidrólisis ácida de los taninos condensados, corregido para dar directamente el resultado en g/L.
El índice de gelatina fue calculado con la siguiente fórmula:
Índice de gelatina (%) = 100 x (TA -TB)/TA
donde:
TA = Taninos del sobrenadante del tubo A
TB = Taninos del sobrenadante del tubo B
TB= (D1 – D2) X 19.33
D1 = DO del tubo B1
D2 = DO del tubo B2
Acidez titulable
A una muestra de 10 g de pulpa se le agregaron 100 ml de agua destilada, se homogenizó y se filtró. Posteriormente, se tituló una muestra de 25 ml con una solución de hidróxido de sodio 0.05 N hasta alcanzar un pH de 8.3, la acidez se calculó con la siguiente fórmula:
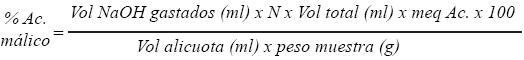
Análisis de granos de polen. Las imágenes de los granos de polen también fueron obtenidas con este microscopio y obtenidas de al menos tres plantas, cuando las anteras aún no abrían, pues una vez abiertas la dispersión es muy rápida y ya no se observan granos.
Se aplicaron 158 encuestas en algunas colonias populares de Uruapan y con personal y alumnos del Instituto Tecnológico Superior de Uruapan para conocer la preferencia de las personas por el nanche o la changunga.
RESULTADOS
El análisis de AFLP, con 172 bandas generadas, muestra un alto grado de similitud entre 25 de los 27 árboles muestreados, excepto dos changungas de Charapendo (CH24 y CH26) (Fig. 1). Además, pareciera presentarse cierta tendencia a que los individuos sean subagrupados de acuerdo al sitio de muestreo; los del Sabino se mantienen juntos en algunas de las ramas superiores que muestra el dendrograma, con algunas excepciones de individuos de otros sitios (CH15C, CH10C) y con dos individuos de Sabino que están fuera (CH12S y CH16S); los de Charapendo también forman al menos un subgrupo en la parte inferior del dendrograma y así los de la ciudad de Uruapan, también con algunas excepciones (CH8U y N1U). Lo más importante y relevante a destacar es que no existe una diferencia genética importante entre los nanches y changungas que se hubiera visto refl ejada en dos grupos fuertemente separados, sino que a pesar de que sí hay pequeños subgrupos de changungas y de nanches, en lo general se encuentran mezclados y esto habla de que son más las similitudes genéticas que comparten ambos tipos de árboles, silvestres y cultivados de Byrsonima crassifolia.
El análisis de frutos mostró que el fruto de nanche es más grande que el de changunga (Fig. 2), sin embargo, en el mercado se venden frutos de changunga injertada sobre nanche, que resultan más grandes que los de nanche. La semilla de nanche pesó en promedio 0.21 g, la de changunga injertada 0.26 g y la de changunga silvestre 0.17 g (Fig. 3). Una diferencia notable se encontró en el pH de la pulpa en nanche, fue de 4.5 y changunga de 3.5; los grados Brix en ambas formas resultaron muy similares: 10.1 para el nanche y 9.1 para la changunga. En cuanto a otras determinaciones, se encontró diferencia estadísticamente significativa en el contenido de cenizas: 2.6% para changunga y 3.04% para nanche. La acidez titulable (como % de ácido málico) fue de 0.0735 para nanche y 0.569 para changunga. Los taninos condensados totales fueron de 0.4478 g para changunga y 0.4978 g para nanche; los taninos astringentes 51.35 para changunga y 46.73 para nanche, como porcentaje de los taninos totales. No se encontraron diferencias en el perfil de grasas (separadas por TLC) entre el nanche y la changunga (dato no mostrado). Se extrajo considerablemente más pectina de la changunga (2.25 g) que del nanche (0.95 g).


Al preguntarles a las personas sobre su preferencia por nanche o changunga se encontró que 27% prefieren el nanche, 18% la changunga y 42% ambos tipos de fruto, con 13% de los individuos contestando que no les agradan, de un total de 158 encuestados.
Al comparar el polen de changunga (Fig. 4) con el de nanche (Fig. 5) no se encontraron diferencias aparentes entre los granos de uno y otro, con el microscopio electrónico de barrido. En algunos casos el polen permite diferenciar al menos a nivel de género y en el caso del maíz, a separarlo del teocintle con base en el tamaño (Perry et al., 2006; Piperno et al., 2007; Pohl et al., 2007). En cuanto a la composición elemental, no se observan diferencias notables entre ambos frutos (cuadro 2).



En otros lugares, como en Tumbiscatio (900 m.s.n.m.), Michoacán, sólo hay changunga (no existe la forma domesticada o nanche) y en Colima, Colima (500 m.s.n.m.) sólo se le conoce como nance, sin diferenciar entre las dos formas, aunque no sabemos si las hay. En este caso, somos los primeros en separar a Byrsonima crassifolia en la forma cultivada (nanche) y la forma silvestre (changunga) y en aislar ADN de ambas.
DISCUSIÓN
Aunque los AFLP parecen agrupar un subconjunto de nanches, y a algunas changungas por otro, en general el grado de similitud entre ambas formas es alto y esto se muestra en que están distribuidos en los diferentes subgrupos formados. Esto es interesante, ya que a pesar de las diferencias morfológicas que presentan los árboles silvestres y cultivados de Byrsonima crassifolia, su información genética es similar. Esto podría indicar que los pobladores han tomado el recurso que tenían más cercano en la naturaleza para llevarlo a sus huertos o solares y por ello los árboles de un mismo sitio tienden a agruparse. La diferencia entre las dos formas se debe a que el proceso de domesticación es reciente y aun no hay una divergencia genética fuerte. En el caso de ciruela (Spondias purpurea) con análisis de secuencias de regiones espaciadoras de cloroplasto se encontró que individuos genéticamente distintos, de diferentes regiones geográfi cas, fueron tomados por los humanos para llevarlos al cultivo (Miller y Schaal, 2005). Así, en el caso de Byrsonima hay una selección de árboles frutales con ciertas características deseables y aunque éstos y sus frutos son reconocidos como diferentes por los agricultores y consumidores, a nivel molecular existe una gran similitud entre ambos. La presencia de sodio y cloro en la changunga acaso podría explicarse por el tipo de suelo en que se encuentra y los menores o nulos cuidados humanos (riego, fertilización). El contenido de magnesio, potasio y fósforo resulta interesante pues sin duda representa un aporte a la dieta en estos elementos. Es de notarse que en la ciudad de Uruapan se plantan árboles de nanches en las calles (aceras) con la finalidad de arbolar y tener frutos, pero hasta ahora no parece haber árboles de changunga plantados en la calle. Según una informante, se debe a que la changunga es de tierra caliente y no se da en Uruapan. Así, para la forma domesticada del árbol, nanche, se está ampliando el área de distribución, cosa que no ocurre con la changunga. Debe señalarse que las huertas de árboles frutales, sobre todo nanche, han desplazado el cultivo tradicional del maíz por la mayor rentabilidad del cultivo, no explicándose que no existan estadísticas oficiales por ejemplo, producción y venta (que podría impedir la obtención de apoyos gubernamentales para la siembra del cultivo). En cambio, las estadísticas se centran en otros cultivos como el maíz, que, como se señaló, ha sido reemplazado en algunos lugares, como en Charapendo, por el nanche y la changunga.
Faltaría un análisis más detallado para establecer diferencias entre ambas, relacionadas con la mayor firmeza del nanche (Rose et al., 1998). Cabe destacar que las diferencias se extienden al modo de cosechar los frutos. Las changungas se arrancan del árbol, mientras que los nanches se recogen del suelo una vez que caen, haciéndose la recolección diaria. Para transportarla y almacenar la changunga se lleva en botes con agua, lo que evita que se maltrate, en cambio, el nanche puede ser transportado y almacenado sin agua. También se refiere que la corteza de la changunga es utilizada para tratar problemas estomacales, no así la del nanche, quizá por la diferencia en el contenido de taninos entre ambas, presumiblemente mayor en la corteza de changunga. La necesidad de usar una concentración mayor de CTAB para la extracción del ADN de la changunga podría estar relacionada con esto ya que el CTAB permite una mayor limpieza del ADN (Nordagard et al., 2005). Una de las características de las plantas domesticadas es que, en general, presentan un menor contenido de metabolitos secundarios.
En las huertas existe cierta variabilidad, por ejemplo, en la época de fl oración, los árboles tardíos podrían ser ventajosos para el fruticultor que podría vender la fruta una vez que pase la temporada principal de producción. Sin embargo, los agricultores no parecen muy interesados por ahora en seleccionar esta característica. Les interesa más tener árboles con fruto grande. Esto también podría deberse a que si son pocos árboles no les convendría transportar la fruta por ser poca, dado que la llevan hasta Pátzcuaro y Morelia, principalmente.
Con la necesidad de llevar la fruta al mercado, para su venta, es posible que la selección de algunas características se intensifique (tamaño, dulzor y apariencia del fruto) y se acelere la pérdida de otras (color rojo, frutos ácidos), como se ha encontrado para ciruela (Miller y Schaal, 2005). A priori, se podría proponer proteger los árboles silvestres de las diferentes localidades a fin de preservar la diversidad genética de esta especie. Es también de tomarse en consideración las presiones del mercado, que han desplazado al maíz, un cultivo ancestral, por otro más rentable económicamente, y que también está ocasionando pérdida de diversidad pues los agricultores no conservan aquellos árboles que no resultan valiosos desde el punto de vista de la venta del fruto. No fue fácil conseguir changunga del cerro (forma silvestre); en Uruapan la venden revuelta con la injertada. En Charapendo debe conseguirse el día que la llevan los recolectores, antes que se agote. En general no se recolecta cuando hace mal tiempo.
Como trabajo a futuro se podría tratar de establecer el número de cromosomas de la especie, para dilucidar si la domesticación ha favorecido o propiciado la poliploidia, como se reporta para otros árboles y plantas cultivadas (Dubcovsky y Dvorak, 2007; Hughes et al., 2007; Srisuwan et al., 2006).
Sin duda es deseable llevar a cabo una exploración más amplia sobre este árbol, su conservación y aprovechamiento. La zona de muestreo es de transición, entre la parte templada (Uruapan) y la denominada "tierra caliente" (Charapendo, Taretan), lo que le añade interés adicional pues en este tipo de regiones se han encontrado evidencias de los inicios de la agricultura y la domesticación de plantas al poderse acceder a los recursos de ambas zonas (Perry et al., 2006). En la región existen ruinas prehispánicas (Tingambato), al parecer, poco exploradas al respecto.
Los resultados aquí expuestos indican que en la región de Uruapan se ha hecho una selección para ciertas características del fruto de Byrsonima crassifolia, como son el tamaño (frutos más grandes) y para mayor dulzor, como se reporta para ciruela (Miller y Schaal, 2005). La semilla también ha sufrido una reducción en tamaño bajo domesticación, de acuerdo con nuestros datos, aunque se traslapa con el de la changunga y quizá sea una característica que se selecciona secundariamente en relación al tamaño del fruto. Otros autores reportan pesos que van de 0.17 a 0.42 g, intervalo dentro del cual caen los pesos hallados en este estudio incluso las medias están por debajo del valor mayor reportado (0.42 g). El peso promedio indicaría una selección para tamaño de semilla pequeña. La reducción en el peso de la semilla, una parte que no interesa al humano, es acorde con los efectos de la domesticación. La changunga tiene ocho veces más ácido que el nanche, lo que sin duda influye en la percepción de los consumidores. En el caso del pH de la pulpa se reportan valores de 2.6 a 4.38, también acordes con lo encontrado (Martínez-Moreno et al., 2006). Los grados Brix referidos por Martínez-Moreno et al. (2006) para Byrsonima van de 7.6 a 12.2, un poco por debajo de los valores encontrados por nosotros, que podría deberse al estado de madurez del fruto al momento de hacer la determinación y también al grado de domesticación (Martínez Moreno et al., 2006). El color del fruto también es importante, las exigencias del mercado impiden que se seleccionen árboles con fruto de color verde pues las personas no los quieren por considerar que aún no maduran. Encontramos árboles de forma intermedia que la gente llama "nanche-changungo" o "nanche agrio" pues no son tan dulces como el nanche ni tan ácidos como la changunga. Los propios fruticultores se deshacen de los árboles que dan fruta roja, a pesar de su color vistoso y llamativo, debido a que la fruta se maltrata fácilmente y no llega en buenas condiciones al mercado. En Tabasco otros investigadores reportan colores rojo, amarillo, verde, anaranjado, haciendo resaltar el rojo como deseable para seleccionar, quizá por no tener en consideración la exigencia del mercado (Martínez Moreno et al., 2006). Existen árboles de changunga silvestres que dan la fruta de color rojo, pero es de esperarse que no sean tomadas para cultivarse en los huertos por esa desventaja. La característica de mayor dulzor es, al parecer, determinante para la mayor preferencia por parte de los consumidores, aunque la búsqueda de changunga por aquellos que la prefieren ha llevado a los agricultores a practicar el injerto de changunga sobre nanche. Estas changungas injertadas dan fruto más grande y durante una temporada más larga del año, pues la changunga silvestre es estacional y da frutos de tamaño pequeño. Cabe la pena señalar que también existen consumidores que bus-can en específico la changunga del cerro.
La preferencia de la changunga para elaborar atoles, paletas de hielo, nieves y licor puede influir favorablemente para lograr la conservación de estos árboles. Lo anterior no es una garantía para la conservación de diversidad genética, además de que podría darse el caso de que se propagaran clones mediante el injerto. En Charapendo se están usando los terrenos como agostadero para el ganado, con la consecuente destrucción de árboles, sobre todo changungos. Quizá la gente prefiera comercializar nanche en fresco que procesar changunga pues no todos tienen las facilidades y el conocimiento. Nosotros elaboramos mermelada de nanche o changunga y ambas resultaron con buena aceptación entre las personas a quienes se les pidió que la probaran. En el caso del nanche, debido a su olor peculiar que desagrada a algunos consumidores, le agregamos esencia de vainilla a la mermelada, logrando enmascarar el olor. Ésta podría ser una alternativa de comercialización. Estos frutos podrían servir como modelo para el estudio de los componentes de la pared celular que intervienen en la firmeza o ablandamiento del fruto (Rose et al., 1998), la búsqueda de genes que intervienen en la dulzura o acidez del fruto, dado que son características contrastantes entre el nanche y la changunga, que podrían también dar mayor información sobre los cambios a nivel genético que han ocurrido entre el nanche y la changunga (Ammiraju et al., 2008; Komatsuda et al., 2007; Yao et al., 2007; Zhongwei et al., 2007).
A nivel molecular hay gran similitud entre los árboles de nanche y changunga pero el proceso de selección y domesticación es muy activo y aunque parecen formar un conjunto como un todo, también se subagrupan como nanches y changungas, por lo tanto serían objeto de gran interés de estudio.
Las diferencias entre los frutos de nanche changunga y las diferencias fenotípicas entre ambos tipos de árboles hacen este sistema un excelente modelo para el estudio de la domesticación de árboles.
AGRADECIMIENTOS
Al M.C. Atahualpa Guzmán de Casa por mostrarnos los sitios de muestreo, a la técnico Emigdia Alfaro por la asistencia en los geles de AFLPs, a los ingenieros Rafael Tapia Aparicio y Rosa Santoyo por la ayuda técnica y al COECYT-Michoacán e Instituto Tecnológico Superior de Uruapan por el financiamiento para la realización de este proyecto. A nuestros informantes Mario Quintana y Camilo Ortiz. A los revisores por sus valiosas sugerencias. Al CONCYTEG convenio 09-11-K119-044.
LITERATURA CITADA
Ammiraju, J.S.S., F. Lu, A. Sanyal, Y. Yu,. X. Song, N. Jiang, A.C. Pontaroli, T. Rambo, J. Currie, K. Collura, J. Talag, C. Fan, J.L. Goicochea, A. Zuccolo, J. Chen, J.L. Bennetzen, M. Chen, S. Jackson, R.A. Wing, 2008. "Dynamic evolution of Oryza genomes is revealed by comparative genomic analysis of a genus-wide vertical data set". Plant Cell, 20: 3191-3209 [ Links ]
Camejo de Aparicio, C., A. Ferrer, B. de Ferrer, J. Peña, M. Cedeño, 1996. "Extracción y caracterización de pectina en limones injertados de la región Zuliana". Rev. Fac. Agron., (LUZ) 13: 641-645. [ Links ]
Dubcovsky, J., J. Dvorak, 2007. "Genome plasticity a key factor in the success of polyploid wheat under domestication". Science, 316: 1862-1866. [ Links ]
Doebley, J.F., B.S. Gaut, B.D. Smith, 2006. "The molecular genetics of crop domestication". Cell, 127: 1309-1321. [ Links ]
Edwards, K.J., 2001. "Miniprep procedures for the isolation of plant DNA". In: Molecular tools for screening biodiversity. Ed. For A. Karp, P.G. Isaac and D.S. Ingram. Kluwer Academic Publishers pp 22-23. [ Links ]
Gallavotti, A., Q. Zhao, J. Kyozuka, R.B. Meeley, M. K. Ritter, J.F. Doebley, M.E. Pè, R.J. Schmidt, 2004. "The role of barren stalk1 in the architecture of maize". Nature, 432: 630-635. [ Links ]
Gentry, H.S., 1982. Agaves of continental North America. The University Arizona Press. Tucson, Arizona U.S.A. [ Links ]
Gil-Vega, K., C., Díaz, A., Nava-Cedillo, J., Simpson, 2006. "AFLP analysis of Agave tequilana varieties". Plant Sci., 170: 904-909. [ Links ]
Glories, Y., 1978. "Recherches sur la matière colorante des vins rousg". Thése a L'Université de Bordeaux II. [ Links ]
Hughes C.E., R., Govindarajulu, A., Robertson, D.L., Filer, S.A., Harris and C.D., Bailey, 2007. "Serendipitous backyard hybridization and the origin of crops". Proc. Nat. Acad. Sci., 104: 14389-14394. [ Links ]
Hyten, D.L., Q. Song, Y. Zhu, I.y Chou, R.L. Nelson, J.M. Costa, J.E.Spech, R.C. Shoemaker, P.B. Cregan, 2006. "Impacts of genetic bottlenecks on soybean genome diversity". Proc. Nat. Acad. Sci., 103: 16666-16671. [ Links ]
Komatsuda, T., M. Pourkheirandish, C. He, P. Azhaguvel, H. Kanamori, D. Perovic, N. Stein, A., Graner, T. Wicker, A. Tagiri, U. Lundqvist, T. Fujimura, M. Matsuoka, M. Yano, 2007. "Six-rowed barley originated from a mutation in a homeodomain-leucine zipper I-class homeobox gene". Proc. Nat. Acad. Sci., 104: 1424-1429. [ Links ]
Kovach, M.J., S.R. McCouch, 2008. "Leveraging natural diversity: back through the bottleneck". Curr Opin. Plant Biol., 11: 193-200. [ Links ]
Loyola-Martínez Liliana, 2006. "Determinación de tres esfingoides y ceramidas por TLC tras la adición de la micotoxina FB, a tejidos vegetales". Tesis de Licenciatura. Química en alimentos, Facultad de Química. UNAM. [ Links ]
Martínez Moreno, E., T. Corona Torres, E. Avitia Garcia, A.M. Castillo Gonzalez, T. Terrazas Salgado, M.T. Colinas León, 2006. "Caracterización morfométrica de frutos y semillas de nanche (Byrsonima crassifolia (L) H.B.K.)". Rev. Chapingo serie horticultura, 12: 11-17. [ Links ]
Meudt, H.M., A.C. Clarke, 2007. "Almost forgotten or latest practice? AFLP applications, analyses and advances". Trends Plant Sci., 12: 106-116. [ Links ]
Miller, A.J., B.A., Schaal, 2005. "Domestication of a Mesoamerican cultivated fruit tree, Spondias purpurea". Proc. Nat. Acad. Sci., 102: 12801-12806. [ Links ]
Miller, A.J., B.A., Schaal, 2006. "Domestication and distribution of genetic variation in wild and cultivated populations of the Mesoamerican fruit tree Spondias purpurea L. (Anacardiaceae)". Mol. Ecol., 15: 1467-1480. [ Links ]
Nordagard, L., T. Traavik, K.M. Nielsen, 2005. "Nucleic acids isolation from ecological samples-vertebrate gut flora". Meth. Enzymol., 395: 38-48. [ Links ]
Onder, M., and M. Babaoglu, 2001. "Interactions amongst Grain Variables in Various Dwarf Dry Beans (Phaseolus vulgaris L.) Cultivars". J. Agronomy & Crop Science, 187: 19-23. [ Links ]
Perry, L., D.H.Sandweiss, D.R. Piperno, K.Rademaker, M.A. Malpass, A. Umire, P. de la Vera, 2006. "Early maize agriculture and interzonal interaction in southern Peru". Nature, 440: 76-79. [ Links ]
Piperno, D.R., J.E. Moreno, J. Iriarte, I. Holst, M. Lachniet, J.G. Jones, A.J. Ranere, R. Constanzo, 2007. "Late Pleistocene and Holocene environmental history of the Iguala valley, Central Balsas watershed of Mexico". Proc. Nat. Acad. Sci., 104: 11874-1881. [ Links ]
Pohl, M.E.D., D.R. Piperno, K.O. Pope, J.G. Jones, 2007. "Microfossil evidence for pre-columbian maize dispersals in the Neotropics from San Andres, Tabasco, Mexico". Proc. Nat. Acad. Sci., 104: 6870-6875. [ Links ]
Rose, J.K.C., K.A. Hadfield, J.M. Labavitch, A.B. Bennett, 1998. "Temporal sequence of cell wall disassembly in rapidly ripening melon fruit". Plant Physiol., 117: 345-361. [ Links ]
Srisuwan, S., D. Sihachakr, S. Siljak-Yakovlev, 2006. "The origin and evolution of sweet potato (Ipomoea batatas Lam.) and its wild relatives through the cytogenetic approaches". Plant Sci., 171: 424-433. [ Links ]
Van Deynze, A., K. Stoffel, C. Robin Buell, A. Kozik, J. Liu, E. van der Knaap, D. Francis, 2007. "Diversity in conserved genes in tomato". BMC Genomics, 8: 465-474. [ Links ]
Vos, P., R. Hogers, M. Bleeker, M. Reijans, T. Van de Lee, M. Hornes, A. Fritjers, J. Pot, J. Peleman, M. Kuiper, M. Zabeau, 1995. "AFLP: a new technique for DNAfingerprinting". Nucleic Acids Res., 23: 4407-4414. [ Links ]
Wang, J.X., T.G. Gao, S. Knapp, 2008. "Ancient literature reveals pathways of eggplant domestication". Ann. Botany, 102: 891-897. [ Links ]
Yao, Y.X., Li M., Liu, Z., Hao Y.J., H., Zhai, 2007. "A novel gene, screened by cDNA-AFLP approach, contributes to lowering the acidity of fruit in apple". Plant Physiol. Biochem., 45: 139-145. [ Links ]
Zhongwei Li, M.E. Griffith, X.Li, Z. Zhu, L. Tan, Y. Fu, W. Zhang, X. Wang, D. Xie, C. Sun, 2007. "Origin of seed shattering in rice (Oryza sativa L.) ". Planta, 226: 11-20. [ Links ]
Zizumbo-Villarreal, D., P.C., Garcia-Marin, E., Payro de la Cruz, P., Delgado-Valerio, P., Gepts, 2005. "Population structure and evolutionary dynamics of wild-weedy-domesticated complexes of common bean in Mesoamerica region". Crop Sci., 45: 1073-1083. [ Links ]

