











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink1. Introducción
El género Bison tiene una historia evolutiva corta y compleja. Evolucionó en Indochina durante el Plio-Pleistoceno, entre hace 3.4 y 2.6 Ma (Kurtén, 1968; McDonald, 1981; Khan et al., 2010; Castaños et al., 2012). Los bisontes se dispersaron por Eurasia hace aproximadamente 700 Ka, durante el Pleistoceno medio y arribaron a Beringia entre hace 300 y 130 Ka (Shapiro et al., 2004). Finalmente, invadieron América del Norte continental entre hace 160 y 75 Ka (McDonald, 1981; Haynes, 1985; Bell et al., 2004; Shapiro et al., 2004), donde evolucionaron dos especies autóctonas: Bison antiquus (syn= B. occidentalis, sensuMcDonald, 1981; Wilson et al., 2008) y Bison latifrons (Guthrie, 1970). Estas dos especies de bisontes se presentan de forma simpátrica con dos especies de origen eurasiático: B. priscus y B. alaskensis (Guthrie, 1970; McDonald, 1981). Todas estas especies coexistieron durante el Pleistoceno tardío de América del Norte, incluyendo México (McDonald, 1981; Ferrusquía-Villafranca et al., 2010), con excepción de B. bison, que se originó durante el Holoceno temprano (4 - 5 Ka) en el sur de Canadá (McDonald, 1981; Wilson et al., 2008).
El bisonte gigante (B. latifrons) es uno de los bisontes menos conocidos de México. Se tienen registros de la especie en San Juan de los Lagos (Dugès, 1894; Solorzano, 2002), Zacoalco (Solorzano, 2002; Lucas, 2008) y Chapala, Jalisco (McDonald, 1981; Solorzano, 2002); la Piedad-Santa Ana y la Cinta-Portalitos, Michoacán-Guanajuato (Díaz-Sibaja et al., 2012); Zumpango de Ocampo (Osborn, 1905) y Tequixquiac, Estado de México (Cope, 1884; Villada, 1903) y dos localidades sin nombrar en Zacatecas (Skinner y Kaisen, 1947). En Oaxaca sólo se tienen registros probables en la Mixteca alta, al norte del estado y en los Valles Centrales (Ferrusquía-Villafranca, 1976; Ferrusquía-Villafranca et al., 2010).
Los bisontes del Pleistoceno formaron parte de un ensamblaje mastofaunístico diverso, de cerca de 286 especies reportadas (Arroyo-Cabrales et al., 2002; Ferrusquía-Villafranca et al., 2010).
Esta riqueza de especies es comparable a la actual, aunque la composición y estructura de las comunidades es distinta (Ceballos et al., 2010; Ferrusquía-Villafranca et al., 2010; Ceballos y Arroyo-Cabrales, 2012). La mayoría de los registros de mamíferos cuaternarios en México corresponden a la NALMA (North American Land Mammal Age, edad de mamíferos terrestres norteamericanos) del Rancholabreano (Arroyo-Cabrales et al., 2002), la cual se caracteriza por la presencia del taxón índice Bison y corresponde al Pleistoceno tardío (160 - 9.5 Ka) (Savage, 1951; Bell et al., 2004). Durante el Rancholabreano se presentaron cambios climáticos importantes como por ejemplo, el Último Máximo Glaciar y el Dryas Reciente (Haynes, 2008; Clark et al., 2009). Estos cambios alteraron la composición de las comunidades mastofaunísticas, lo que se toma como referencia para establecer los inicios del Holoceno (Ceballos et al., 2010). El estudio de las faunas correspondientes al Rancholabreano contribuye al entendimiento de la fundación del Holoceno y de la respuesta de las comunidades de mamíferos al cambio climático (Arroyo-Cabrales et al., 2008).
Las faunas del Pleistoceno tardío en Oaxaca corresponden en su mayoría a la región de la Mixteca alta, localizada al noroeste del estado (Pérez-Crespo et al., 2008; Jiménez-Hidalgo et al., 2011) y a la región de los Valles Centrales (Pérez-Crespo et al., 2008). En este trabajo presentamos el registro más austral de Bison latifrons, su fauna asociada y el análisis de dieta de un ejemplar de mamut, procedente de una nueva localidad fosilífera del Rancholabreano en los Valles Centrales de Oaxaca.
2. Área de estudio
San Dionisio Ocotepec se localiza en el municipio del mismo nombre, al este de los Valles Centrales de Oaxaca. La localidad se encuentra entre las coordenadas 16°48’ - 16°47’ N y 96°24’ - 96°22’ O, con afloramientos fosilíferos en un par de arroyos de flujo intermitente denominados “El Pedregal” y “La Salina”. La fisiografía de la zona es heterogénea, con presencia de zonas planas utilizadas con fines agrícolas y ganaderos, así como una amplia representatividad de arroyos y cañadas (Figura 1).

Figura 1. Ubicación geográfica y mapa hipsográfico de los yacimientos que forman la fauna local de San Dionisio Ocotepec.
El yacimiento se localiza en una zona con aluvión Cuaternario que sobreyace a tobas andesíticas y andesitas del Neógeno y secuencias de areniscas y lutitas de la Formación Teposcolula-Ocotlán (Cenomaniano-Albiano) y sólo en el sureste del sitio se encuentran calizas y lutitas de la Formación Yucunama (Sánchez-Rojas et al., 2000).
La estratigrafía del sitio se compone principalmente por una alternancia de secuencias fluviales y volcanoclásticas (Figura 2). La columna tipo de la localidad se describe de base a cima en la Tabla 1.

Tabla 1. Estratigrafía generalizada de San Dionisio Ocotepec, elaborada a partir de cuatro columnas litológicas.
| Número de estrato | Descripción |
|---|---|
| 1 | 70 cm de arena volcánica andesítica fuertemente compactada. Contacto superior erosionado. |
| 2 | 109 cm de arcilla con arena de color verde y con presencia de concreciones de carbonato de calcio de hasta 20 cm de diámetro. Contacto superior erosionado. |
| 3 | 9 cm de arena volcánica andesítica de color rojizo fuertemente consolidada, con lentes de arcilla color café. Contacto superior neto. |
| 4 | 20 cm de arcilla con limo de color café. Contacto superior erosionado. |
| 5 | 9 cm de ortoconglomerados polimícticos con clastos de hasta 2 cm de diámetro. Contacto superior neto, con presencia de oxidación. |
| 6 | 10 cm de arcilla de color café. Contacto superior erosionado. |
| 7-11 | 40 cm de una intercalación de ortoconglomerados con brechas, ambos polimícticos y compuestos de clastos de riolita, basalto, granito y feldespatos, con contactos netos entre niveles y un contacto superior para el bloque de carácter fuertemente erosivo. |
| 12 | 112 cm de arcilla con limo, de color café obscuro, con algunos clastos riolíticos y basálticos embebidos en la matriz y presencia de carbonatación hacia la base. Presencia de restos fósiles de vertebrados. Los fósiles se presentan tanto en la base como en la cima. Contacto superior erosionado. |
| 13 | 28 cm de brecha de color amarillento, con clastos de hasta 10 cm de diámetro. Contacto superior erosivo. |
| 14 | 125 cm de arcilla con limo de color negro, con lentes de arcilla rojiza con oxidación y concreciones calcáreas de hasta 10 cm de diámetro máximo. Contacto superior neto, ligeramente oxidado. |
| 15 | 83 cm de toba retrabajada blanquecina con abundante oxidación. Contacto superior neto. |
| 16 | 30 cm de suelo (regosol eútrico) residual moderno, con evidencia de pedogénesis in situ. |
3. Material y método
Los fósiles se hallaron fortuitamente durante la construcción de un pozo de riego. En la zona del hallazgo se llevó a cabo el levantamiento de columnas litológicas para corroborar la ubicación estratigráfica de los restos fósiles. Adicionalmente, se llevaron a cabo salidas al campo para el levantamiento de la estratigrafía de la zona y la colecta de material fósil adicional en zonas aledañas al hallazgo original y en las barrancas y los arroyos de San Dionisio Ocotepec. El material colectado fue depositado en la Colección Paleontológica del Laboratorio de Paleontología de la Universidad Michoacana de San Nicolás de Hidalgo (con las iniciales UM) y en la colección del proyecto de Museo de Sitio de San Dionisio Ocotepec (con las iniciales MSDO), el cual se encuentra bajo resguardo de la presidencia municipal.
Para la identificación de los restos fósiles se empleó bibliografía especializada y comparación directa. La comparación directa se llevó a cabo con los materiales de la Colección Científica del Laboratorio de Paleobiología, Instituto de Recursos, Universidad del Mar; la Colección Paleontológica del Laboratorio de Paleontología, Universidad Michoacana de San Nicolás de Hidalgo (UMSNH); la colección virtual del Museo de Paleontología de la Universidad de Michigan (UMMP VP); el material del Museo Estatal de Indiana (ISM) y del Museo Nacional de Historia Natural del Instituto Smithsoniano (USNM).
También se realizó un análisis de microdesgaste dental de baja magnificación a las piezas dentales de Mammuthus columbi (UM 1340, UM 1338), siguiendo el protocolo de Solounias y Semprebon (2002). En total, se llevaron a cabo tres moldes por pieza dental (maxilar y mandibular), de tres zonas: mesial, media y distal. Los moldes se observaron en un microscopio esteroscópico a 35x, del que se obtuvieron fotografías que fueron procesadas en el software Microware 4.02 (Ungar, 2002). Posteriormente, se obtuvo un promedio de hoyuelos y estrías para cada fotografía, que fue vertido en una base de datos de ungulados modelo modernos, tomada de Solounias y Semprebon (2002). Se hizo un análisis bivariante de promedio de estrías contra promedio de hoyuelos, así como análisis de funciones discriminantes canónicas (AFDC). Para el AFDC se usó un modelo de discriminación lineal y covarianza común, con las variables: promedio de hoyuelos y promedio de estrías; el soporte estadístico fue evaluado mediante una prueba de Lambda de Wilk, para corroborar la separación de categorías dietarias (pacedor, mixto y ramoneador). Finalmente, se llevó a cabo un análisis de mesodesgaste para proboscídeos, según la metodología propuesta por Saarinen et al. (2015); donde se promediaron los ángulos de los valles de dentina entre láminas de esmalte de seis zonas en los dos dientes reportados. Estos análisis se llevaron a cabo para inferir el tipo de hábitat en el que vivió la fauna de esta nueva localidad.
4. Resultados
En total se estudiaron más de 50 restos fósiles de los cuales, sólo 15 resultaron ser diagnósticos. La mayor parte de los restos procede del arroyo El Pedregal y sólo tres proceden de La Salina. Se reportan cuatro taxones en total: Mammuthus columbi, Equus cf. E. conversidens, Bison sp. y Bison latifrons.
4.1. PALEONTOLOGÍA SISTEMÁTICA
Orden Proboscidea Illigeri, 1811
Familia Elephantidae Gray, 1821
MammuthusBrookes, 1828
Mammuthus columbi (Falconer, 1858)
Material referido. UM 1340, M2 derecho completo, con fragmentos del maxilar; UM 1338, m2 derecho completo; UM 1339, vértebra lumbar (L1); UM 1341 unciforme derecho completo; UM 1342, costilla izquierda; MSDO 001, fragmento distal de defensa izquierda; MSDO 002, MSDO 003 (Figura 3) y MSDO 004, fragmentos de escápulas y MSDO 005, epífisis parcial de un húmero derecho.

Figuras 3. Restos fósiles de Mammuthus columbi. a, b) UM 1340 en vistas oclusal y labial; c, d) UM 1338 en vistas oclusal y labial; e, f, g) UM 1339 en vistas craneal, lateral izquierda y dorsal; h, i) UM 1341 en vistas medial y anterior; j) UM 1342 en vista especular posterior; k, l) MSDO 001 en vistas posterior y medial. Abreviaturas: MX, maxilar; PE, proceso espinoso; PT, proceso transverso; CV, centro vertebral; PAPE, porción anterior del proceso espinoso; FAPP, faceta articular de los procesos articulares caudales (posteriores); PAP, proceso articular caudal; PAC, proceso articular craneal; FAPC, faceta articular del proceso articular craneal; SAOMD, superficies articulares dorsales del os magnum; SAOMV, superficie articular ventral del os magnum; CP, cavidad pulpar; DE, dentina; FACD, faceta articular caudal; CA, cabeza; CU, cuello; TC, tubérculo costal; TIC, tuberosidad íleo-costal; SC, surco costal. Barra de escala: 10 cm.
Descripción y asignación taxonómica. Los molares (Figura 3a-d) presentan la morfología típica de un elefántido elefantino, con dientes hipsodontes y loxodontes de láminas compactas (Agenbroad y Brunelle, 1992; Osborn, 1942). Las piezas presentan 6.5 placas por cada 100 mm, lo que concuerda con el número diagnóstico para Mammuthus columbi de 6 - 7/100 mm en M2 y 5.5 - 6.5/100 mm en m2 (Osborn, 1942). La edad biológica del ejemplar corresponde con 26 ± 1 años, según el patrón de desgaste en las láminas que conforman los molares (Agenbroad y Brunelle, 1992).
La vértebra lumbar (Figura 3e-g) presenta la morfología típica de un mamut, con procesos transversales delgados y no incorporados al centro vertebral, el cual posee una forma ovalada y presenta un margen inferior en forma de U (Hodgson et al., 2008; Olsen, 1972). Su comparación con el ejemplar UMMP VP 116967, permitió identificarla como una lumbar 1 (L1), ya que en ambas el centro vertebral es más alto que ancho, la faceta articular de los procesos articulares craneales tienen forma de V en vista dorsal, los procesos transversos están orientados de forma caudal y son delgados, el margen dorsal de los procesos articulares caudales es redondeado y finalmente, la porción anterior del proceso espinoso inicia alrededor del 50 % del ancho del centro vertebral.
Por su parte, en vistas ventral y posterior el unciforme (Figura 3h-i) presenta una forma subpiramidal, en vista anterior presenta una forma cuadrangular (Olsen, 1972); en vista medial, las superficies articulares del os magnum son anchas y se encuentran separadas por un istmo amplio hacia la zona volar; finalmente, la superficie articular inferior tiene un margen recto poco prominente (Hodgson et al., 2008).
Asumiendo que el dimorfismo sexual de defensas de mamut es similar al de elefántidos modernos, el fragmento de defensa (Figura 3k-l) es referido como perteneciente a un macho, debido a la forma fuertemente triangular de la sección transversal de la misma, en contraposición con la sección oval de las hembras (Elder, 1970).
Al comparar la costilla de mamut asociada a los restos diagnósticos con costillas de Mammut americanum (ISM 71.3.261) y Mammuthus primigenius (USNM 23792), el elemento UM 1342 (Figura 3j) se asigna al género Mammuthus. La costilla de M. columbi referida es larga, delgada (Hodgson et al., 2008) y en sección transversal, presenta una marcada forma de S, con prominentes cabeza y faceta articular caudal, un cuello relativamente largo, un tubérculo costal prominente y una tuberosidad íleo-costal con zonas de inserción muscular para el músculo íleo-costal torácico prominentes, además de un surco costal profundo. Es probable que la costilla proceda de la región media del cuerpo por la curvatura del ángulo (Hodgson et al., 2008). El resto de las piezas se encontraron asociadas a las piezas diagnósticas, y presentan la morfología típica de Mammuthus (Olsen, 1972) por lo que se infiere que pertenecieron al mismo individuo.
Orden Perissodactyla Owen, 1848
Familia Equidae Gray, 1821
EquusLinneo, 1758
Equus cf. E. conversidensOwen, 1869
Material referido. UM 1343, una vértebra cervical (C4-5) y UM 1344, una vértebra torácica de la región anterior (T5-7) (Figura 4).

Figura 4. Restos fósiles de Equus cf. E. conversidens. a-c) UM 1343, en vistas craneal, lateral izquierda y dorsal. d-f) UM 1344, en vistas craneal, lateral derecha y ventral. Abreviaturas: PN, proceso neural; CV, centro vertebral; CV, cresta ventral; FL, foramen lateral; PT, proceso transversal; BPN, bifurcaciones del proceso espinoso; PAC, proceso articular caudal; FAC, faceta articular craneal; FAD, faceta articular caudal (distal). Barra de escala: 5 cm.
Descripción y asignación taxonómica. La vértebra cervical (Figura 4) presenta las características típicas del género. Posee un centro vertebral inclinado en plano sagital anteroposteriormente, con forma acorazonada en las extremidades craneal y caudal; el proceso transverso sigue el eje del centro vertebral y se encuentra roto en su base; el proceso neural es corto y poco pronunciado, único en la región anterior y bifurcado en la posterior (formando una W); posee una cresta ventral bien desarrollada hacia la región posterior y presenta forámenes laterales bien definidos (Barone, 1995; Klaus-Dieter et al., 2009; Pales y Garcia, 1981). Sólo se conserva el proceso articular caudal derecho, el cual rebasa el margen distal del centro vertebral en vista dorsal y posee una forma oval e inclinada medialmente.
La vértebra torácica (Figura 4) no posee el arco neural; presenta un centro vertebral inclinado en plano sagital anteroposteriormente y comprimido lateralmente en la región central; el centro posee forma semicircular, el margen dorsal es recto. Se reconoce como vértebra torácica por la posesión de las facetas costales craneal y caudal derechas y como séptima a quinta dorsal por la ausencia del foramen vascular, típico de vértebras torácicas posteriores (Barone, 1995; Klaus-Dieter et al., 2009). Se comparó la altura y la longitud del centro vertebral del elemento UM 1344 con Equus conversidens y Equus cf. E. cedralensis de la Colección Paleontológica de la UMSNH y el elemento UM 1344 es similar en dimensiones a E. conversidens (Tabla 2).
Tabla 2. Medidas de vértebras torácicas seleccionadas de Equus de México. Abreviaturas: LCDe, longitud máxima del cuerpo vertebral, incluyendo el diente; BFcr, ancho máximo en la facies articularis cranialis; BFcd, ancho máximo en la facies articularis caudalis. Medidas en mm.
| Elemento | Taxón | LCDe | BFcr | BFcd | Altura del centro | Largo del centro |
|---|---|---|---|---|---|---|
| UM 1344 | Equus cf. E. conversidens | 64.9 | 48.2 | 51.2 | 48.1 | 70.6 |
| UM 1346 | E. cedralensis | 33.6 | 32.1 | 35.1 | 27.5 | 39.9 |
| UM 1347 | E. cedralensis | 37.8 | 28.8 | 31 | 25.2 | 36.3 |
| UM 1348 | E. cedralensis | 40.9 | 30.6 | 32.5 | 29.6 | 44.4 |
| UM 1349 | E. cedralensis | 40.9 | 33.8 | 32.2 | 28.7 | 40.6 |
| UM 1350 | E. cedralensis | 40.1 | 29.8 | 29.2 | 26.8 | 42.3 |
| UM 1351 | E. conversidens | 63.9 | 37.5 | 45.6 | 43.3 | 68.4 |
| UM 1352 | E. conversidens | 58.2 | 45.3 | 49.2 | 50.8 | 68.2 |
| UM 1353 | E. conversidens | 51.6 | 42.3 | 47.4 | 48.8 | 61.8 |
Debido a la similitud de talla entre los elementos UM 1343 y UM 1344 y debido a que Equus conversidens es el équido de talla media reportado para el Rancholabreano de México (Priego-Vargas et al., 2016), ambos elementos se asignan a este taxón.
Orden Artiodactyla Owen, 1848
Familia Bovidae Gray, 1821
Bison (Hamilton-Smith, 1827)
Bison sp.
Material referido. UM 1345, un acetábulo derecho.
Descripción y asignación taxonómica. El acetábulo (Figura 5) muestra la morfología típica de un bovino de gran talla, que se corresponde con Bison (McCuaig-Balkwill y Cumbaa, 1992). Posee un borde acetabular semicircular, la superficie semilunar es amplia y se encuentra casi cerrada, con una incisura acetabular estrecha y una fosa acetabular profunda y notoria. La superficie semilunar del pubis emerge de una eminencia íleo-púbica notoria y amplia; en vista cenital, el borde acetabular se encuentra hendido en su porción dorsal, lo que permite apreciar parte de la superficie semilunar inferior (Barone, 1995; Schmid, 1972).

Figura 5. Acetábulo de Bison sp. UM 1345. a) Vista lateral, b) vista caudal. Abreviaturas: BA, borde acetabular; SS, superficie semilunar; IA, incisura acetabular; FA, fosa acetabular; EIP, eminencia íleo-púbica; PU, pubis. Barra de escala: 5 cm.
Bison latifrons (Harlan, 1825)
Material referido. UM 1346, parte proximal del radio derecho de un macho adulto, preservado desde el sulco tendíneo y hasta la zona articular proximal y con la cabeza completa (Figura 6).
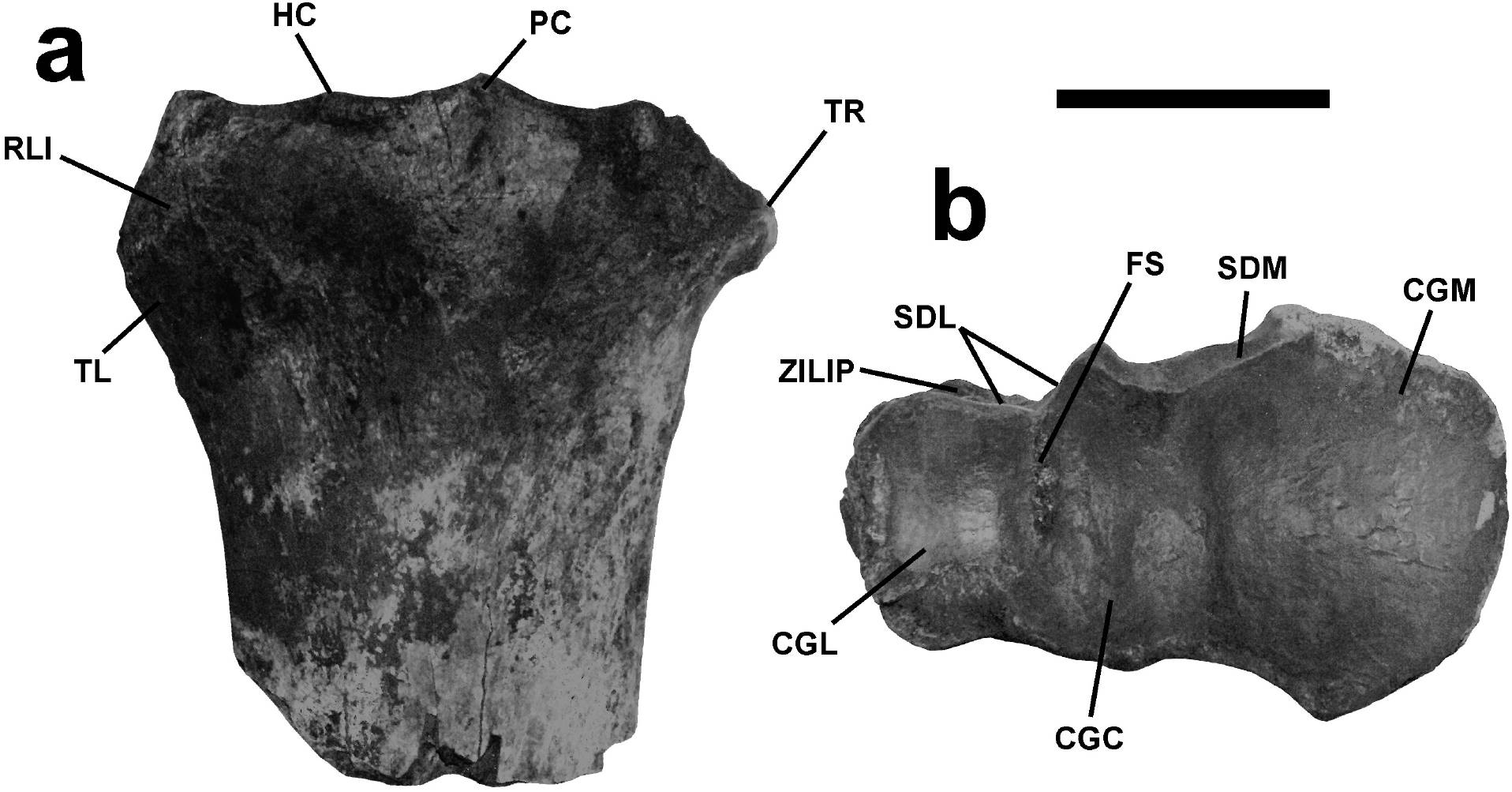
Figura 6. Radio de Bison latifrons, UM 1346. a) Vista anterior, b) vista dorsal. Abreviaturas: RLI, relieve lateral de inserción; TL, tuberosidad lateral; HC, hendidura central; PC, proceso coronoides; TR, repisa de la tuberosidad radial; ZILIP, zona de inserción del ligamento interóseo proximal; SDL, superficies diartrodiales laterales; SDM, superficie diartrodial medial; FS, foseta sinovial; CGM, cavidad glenoidea medial; CGC, cavidad glenoidea medial (central); CGL, cavidad glenoidea lateral. Barra de escala: 5 cm.
Descripción y asignación taxonómica. El radio presenta las características típicas de un bovino de gran talla y se identifica como Bison por su morfología general (McCuaig-Balkwill y Cumbaa, 1992). Por debajo de la repisa de la tuberosidad radial y hacia la zona medial, se presenta una cicatriz para el músculo bíceps braquial bien desarrollada. Ésta posee forma cuadrada y se extiende hacia la zona posterior. La repisa de la tuberosidad radial es amplia y se encuentra expandida medialmente. La tuberosidad lateral o externa es amplia y posee un relieve lateral de inserción en un ángulo aproximado de 45° con respecto al eje articular. Este relieve es amplio y la zona de inserción del extensor dorsal del dedo (extensor digiti quarti propius) es mayor y dispuesta de forma más anterior que la zona de inserción del extensor lateral del dedo (digitorum communis). El proceso coronoide es redondeado y poco prominente; la hendidura central es amplia y presenta una curva suave; en vista cenital, la faceta articular proximal presenta una cavidad glenoidea medial (o fosa capitular) amplia y separada de la cavidad glenoidea media y lateral, que son más pequeñas y cuadrangulares. Finalmente, la hendidura articular de la ulna es amplia y presenta un ángulo obtuso de aproximadamente 135° (Olsen, 1960; McCuaig-Balkwill y Cumbaa, 1992; Barone, 1995; Pales y Garcia, 1981; France, 2009). Para el Rancholabreano de Norteamérica y México se reportan cuatro especies de bisontes (McDonald, 1981; Ferrusquía-Villafranca et al., 2010), de los cuales dos eran gigantes: el eurasiático Bison alaskensis y el autóctono Bison latifrons. Sin embargo, consideramos que este radio perteneció a B. latifrons por sus rasgos morfológicos. Comparamos la morfología de radios descrita para B. priscus debido a que B. alaskensis es conocido únicamente de cráneos y algunos autores lo refieren como una subespecie de B. priscus (e.g.Guthrie, 1966), además de que es parte del grupo de bisontes de origen eurasiático (McDonald, 1981).
Los radios de B. priscus son largos y gráciles, pero con epífisis gruesas, lo que les da un aspecto de tener una cabeza amplia (Shpansky et al., 2016), en contraposición, el elemento UM 1346 no presenta una región epifisiaria notablemente expandida (Figura 6). En B. priscus la ulna se encuentra fuertemente anquilosada al radio (Prat et al., 2010; Shpansky et al., 2016), mientras que en UM 1346, se observan claramente la superficie de sinostosis radio-ulnar y la zona de inserción del ligamento interóseo proximal y no existe evidencia de anquilosis, lo que sugiere la presencia de un espacio interóseo proximal bien desarrollado. Además, en B. priscus (Figura 7) el relieve lateral de inserción se presenta en un ángulo cercano a 90°, presenta una tuberosidad lateral muy grande, alta y ligeramente hendida, con un borde dorsal que se encuentra a la misma altura del proceso coronoide y en algunos ejemplares, incluso lo excede en altura (Prat et al., 2010). Ninguna de estas características está presente en UM 1346.
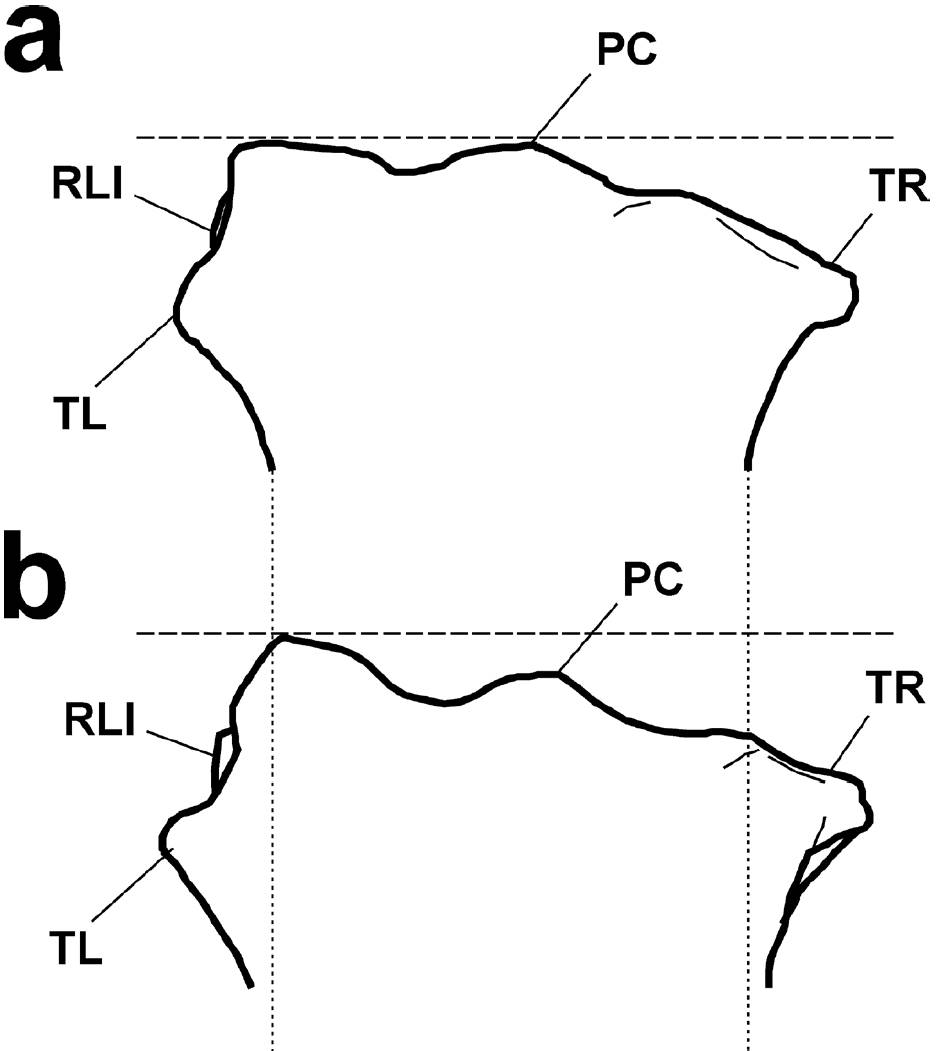
Figura 7. Comparación de la región proximal en radios de a) Bison latifrons y b) B. priscus en vista anterior. La línea punteada gruesa representa el punto más alto del tubérculo lateral. La línea punteada delgada indica el ancho relativo de la diáfisis en B. latifrons. Abreviaturas igual que en la figura 6. Los diagramas no están a escala.
En Oaxaca, se ha reportado la presencia de Bison antiquus (Jiménez-Hidalgo et al., 2011, 2013). El espécimen UM 1346 se asigna a un ejemplar macho de B. latifrons por su talla (Figura 6) y por comparación con radios de B. latifrons (UM 1264) y B. antiquus (UM 1347 y UMPE 0004). B. antiquus presenta una foseta sinovial en forma de moño, que ingresa a las cavidades glenoideas (medial y media) desde el punto intermedio de las superficies diartrodiales, mientras que la foseta de B. latifrons es recta, está dirigida hacia la parte anterior e ingresa únicamente a la cavidad glenoidea media desde el punto central de las superficies diartrodiales laterales. En B. antiquus el margen anterior de la cavidad glenoidea medial es ligeramente subtriangular, mientras que en B. latifrons forma un ángulo bastante prominente. En B. latifrons la repisa de la tuberosidad radial está más acentuada y el proceso coronoide es más redondeado. En la región posterior, la superficie diartrodial medial de B. latifrons es más grande y larga, rebasando el margen ventral de las superficies diartrodiales laterales. Además, la superficie diartrodial medial de B. latifrons es menos pronunciada que en B. antiquus. Adicionalmente, la inserción del ligamento interóseo proximal es mucho más amplia y expandida posteriormente en B. antiquus. Finalmente, el ancho y largo de las superficies articulares proximales del radio UM 1346 se corresponden en sus dimensiones con las reportadas de B. latifrons machos (Figura 8).
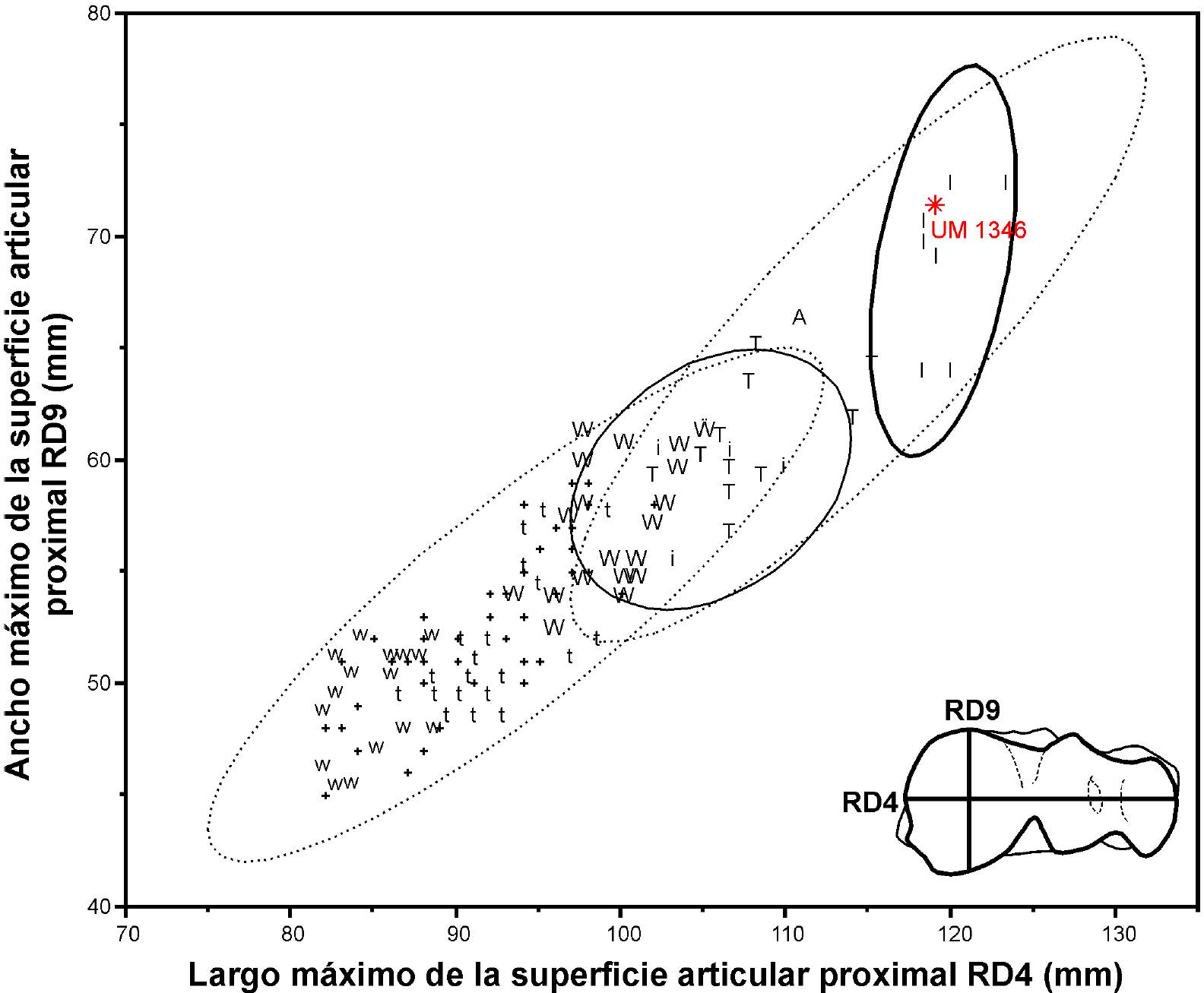
Figura 8. Análisis bivariante de radios de bisontes. Las elipses representan el 95 % de la confianza. Elipse punteada inferior de B. antiquus, elipse punteada superior de B. latifrons. Elipse sólida delgada de hembras de B. latifrons. Elipse sólida gruesa de machos de B. latifrons. Simbología i, I: hembras y machos (respectivamente) de Reserva de American Falls, Idaho; t, T: hembras y machos (respectivamente) de Lipscomb, Texas; w, W: hembras y machos (respectivamente) de Horner II, Wyoming; A: macho de Gallelli Gravel Pit, Alberta; +: Horner II, Wyoming (datos sin sexo especificado). I, i: Bison laifrons. T, t, W, w, A, +: Bison antiquus. Datos tomados de (Stevens, 1978; Todd, 1987; Todd et al., 1992; Wilson et al., 2008).
4.2. INFERENCIA ALIMENTARIA
El único taxón analizado fue Mammuthus columbi, dado que fue el único con piezas dentales conservadas. El análisis de microdesgaste dental (Figura 9) sugiere que, en los últimos días de su vida, el individuo de mamut fue pacedor. Esta asignación dietaria se soporta con el análisis de funciones discriminantes, donde la pieza mandibular se asigna a dicho gremio con una probabilidad de 61 % y la maxilar con una probabilidad de 45.3 %. Además, la prueba de Lambda de Wilk muestra diferencias estadísticamente significativas entre gremios (ramoneadores, mixtos, pacedores), reforzando así esta asignación.

Figura 9. Análisis bivariante de microdesgaste dental de baja magnificación (35x), datos de Solounias y Semprebon (2002). Simbología: triángulos con línea gruesa: ramoneadores, triángulos con línea delgada: frugívoros, círculos con líneas delgadas: mixtos estacionales, círculos con líneas gruesas: mixtos no estacionales, rombos:pacedores. Ecoespacios principales delimitados por polígonos punteados.
A diferencia del análisis de microdesgaste dental, el análisis de mesodesgaste representa la preferencia dietaria a lo largo de la vida de un individuo (Fortelius y Solounias, 2000; Saarinen et al., 2015). Los fósiles analizados poseen un promedio de ángulos de 125.7° (m2) y de 125.5° (M2). Estos valores (Figura 10) sugieren que el ejemplar de M. columbi de este estudio tenía una dieta dominada por plantas C4 (>70 %), lo cual es consistente con un hábito pacedor no estricto (Saarinen et al., 2015).

Figura 10. Mesodesgaste de proboscídeos y su relación con valores isotópicos de carbono y gremios conocidos. Modificado de Saarinen et al. (2015).
5. Discusión
5.1. ORIGEN DE BISON LATIFRONS Y EDAD RELATIVA DE SAN DIONISIO OCOTEPEC
Se considera que el Rancholabreano inició hace 160 Ka antes del presente, usando como taxón índice al género Bison y en particular a B. latifrons (Bell et al., 2004). Sin embargo, los registros tempranos usados para establecer esta edad son dudosos (Scott y Lindvall, 1970; Scott et al., 1982; Haynes 1985). Por otro lado, la evidencia molecular apunta a que los bisontes invadieron América del Norte en dos oleadas, la más temprana situada entre 195 y 135 Ka, pero representada únicamente en Beringia por formas eurasiáticas como Bison priscus (Froese et al., 2017). Los autóctonos americanos como Bison latifrons aparecieron posteriormente en América continental (McDonald, 1981). Esta especie se conoce de un intervalo de entre 120 Ka (Froese et al., 2017; Miller et al., 2014) y 6.4 a 7.7 Ka antes del presente (Dillehay, 1974; Gagliano, 1967). Por consiguiente, asignamos la fauna local de San Dionisio Ocotepec a la edad de mamíferos terrestres norteamericanos del Rancholabreano (Savage, 1951; Bell et al., 2004).
5.2. DISTRIBUCIÓN GEOGRÁFICA
Mammuthus columbi se ha reportado en casi todos los estados del país, con excepción de Quintana Roo, Yucatán, Campeche, Tabasco, y Colima (Arroyo-Cabrales et al., 2007). Para Oaxaca se tienen al menos seis reportes de la especie (Arroyo-Cabrales et al., 2003; Pérez-Crespo et al., 2008), de los que dos registros (Guadalupe Victoria y San Pablo Etla) son referibles a la región noroeste de los Valles Centrales (Pérez-Crespo et al., 2008). El registro de este trabajo extiende la distribución de la especie hacia el sur en aproximadamente 55 Km.
Por otra parte, el género Equus se ha registrado en casi todas las provincias morfotectónicas del país, con excepción de la Sierra Madre Occidental y la Planicie Costera del Golfo-Sur (Priego-Vargas et al., 2016). Equus conversidens se presenta en las provincias morfotectónicas de: Sierras y Planicies del Noroeste, Cordilleras y Planicies de Chihuahua-Coahuila, Meseta Central, Sierra Madre Oriental, Cinturón Volcánico Transmexicano, Planicie Costera del Golfo-Norte, Península de Yucatán, Sierra Madre de Chiapas y Sierra Madre del Sur (Priego-Vargas et al., 2016). En Oaxaca se ha reportado previamente la presencia de E. excelsus en Ejutla y Tehuantepec (Pérez-Crespo et al., 2008), E. mexicanus en Teposcolula (Pérez-Crespo et al., 2008) y la Mixteca Alta (Jiménez-Hidalgo et al., 2012). Finalmente, E. conversidens ha sido reportado únicamente para la Mixteca (Jiménez-Hidalgo et al., 2012). El posible registro de E. conversidens de San Dionisio Ocotepec conecta las localidades de la Mixteca Alta con las de la depresión central de Chiapas (Carbot-Chanona y Gómez-Pérez, 2014). Los bisontes (Bison spp.) están más o menos bien representados en México, sus registros son abundantes en las provincias morfotectónicas de Meseta Central, Sierra Madre Oriental y Cinturón Volcánico Transmexicano; mientras que hay pocos registros en las provincias de Península de Baja California, Sierras y Planicies del Noroeste, Cordilleras y Planicies de Chihuahua-Coahuila y Península de Yucatán (Ferrusquía-Villafranca et al., 2010). Por otro lado, se presenta sólo un registro de Bison tanto en la provincia de la Sierra Madre del Sur (Jiménez-Hidalgo et al., 2011, 2013) como en la Sierra Madre de Chiapas (Carbot-Chanona y Vázquez-Bautista, 2006). En Oaxaca, este registro corresponde con la fauna local de Viko Vijin en la Mixteca alta, donde se reporta la presencia de la especie más abundante en México, Bison antiquus (Jiménez-Hidalgo et al., 2013).
5.3. ESTRATIGRAFÍA
El análisis estratigráfico sugiere que los depósitos de San Dionisio son de carácter predominantemente fluvial. La presencia de concreciones de carbonato de calcio en forma de caliche y carbonatación sugiere la presencia de condiciones áridas (Schlesinger, 1982). Estas condiciones se encuentran en facies someras dominadas por el depósito de arcillas finas (en los estratos 2, 12 y 14), que sobreyacen sedimentos alóctonos de origen volcánico y perturbaciones estocásticas del régimen fluvial evidenciadas por la presencia de conglomerados y brechas. Estas condiciones sugieren que las facies someras presentaban pedogénesis in situ incompleta, que no dio origen a paleosuelos, pues este proceso se vio interrumpido en varias ocasiones por el depósito de nuevos sedimentos, culminando hacia la cima por el arrastre de sedimentos volcánicos alóctonos y la formación de suelos modernos.
5.4. TAFONOMÍA
Los restos fósiles de San Dionisio Ocotepec sugieren que el efecto de ponderación temporal (Time-averaging) es mínimo, particularmente en el sitio “El Pedregal”, donde se encuentran los taxones: Mammuthus columbi, Equus cf. E. conversidens y Bison latifrons. Estos restos fósiles presentan escasas evidencias de intemperismo: poseen una superficie ósea con poco agrietamiento, no se presenta descamación y las superficies articulares presentan grietas en mosaico; lo que sugiere que los restos tuvieron un tiempo de exposición al medio que va de los días a los tres años como máximo (Behrensmeyer, 1978). Adicionalmente, la evidencia muestra que el arrastre de las piezas fue escaso, los bordes son completos y hay presencia de fracturas en espiral (causadas por compresión) con márgenes angulosos (Fernández-Jalvo y Andrews, 2003; 2016). El radio de Bison latifrons se encuentra dentro del grupo II de Voorhies, lo que indica que su transporte fue moderado, principalmente como carga tractiva (Voorhies, 1969). Finalmente, los restos de M. columbi descritos tienen índices de transporte fluvial de entre 53.9 % y 86.8 % (Frison y Todd, 1986), estos elementos corresponden a un único individuo adulto y fueron encontrados asociados. Sin embargo, el resto del esqueleto no se pudo rescatar debido a la construcción de un pozo de riego.
5.5. DESGASTE DENTAL
El microdesgaste dental representa la preferencia dietaria con un margen de horas a días antes de la muerte (Solounias y Semprebon, 2002). Estos patrones reflejan el tipo de vegetación disponible cercana a la zona de depósito (Solounias y Moelleken, 1994). Por otro lado, el mesodesgaste refleja la preferencia dietaria a lo largo de la vida útil de la pieza dental analizada (Saarinen et al., 2015). Estos patrones de desgaste se emplean para llevar a cabo inferencias paleoambientales y predicciones dietarias para las especies con muestras de al menos diez individuos (Fortelius y Solounias, 2000). Estas aproximaciones ofrecen información ambiental limitada con tamaños de muestra inferiores a 10, incluyendo en ocasiones un individuo (Bernor et al., 2014; Danowitz et al., 2016; Hayek et al., 1992). Sin embargo, estas inferencias deben ser tomadas como tentativas y no ser empleadas para hacer generalizaciones paleoecológicas de las especies o del sitio de depósito.
Los resultados de los análisis de desgaste en el ejemplar de M. columbi indican que el individuo fue un pacedor no estricto que se alimentó preferentemente de pastos en los últimos días a horas antes de su muerte. Esto se corresponde con lo que muestran otros estudios de microdesgaste llevados a cabo con esta especie en otras regiones de México (Gutiérrez Bedolla et al., 2016). En el caso del mesodesgaste, el promedio de ángulos de los molares de M. columbi de este estudio fue de 125.6°, lo que sugiere una dieta mixta, dominada por plantas C4 (poáceas). Esto es distinto a lo encontrado para la misma especie en Rancho la Brea (Saarinen et al., 2015), con un promedio de ángulos de 135.5° y a lo encontrado en ejemplares del centro-este de Puebla, donde se obtuvo un promedio de 135° (Carbot-Chanona et al., 2018), lo que sugiere que en estos sitios M. columbi era un pacedor casi estricto, con un consumo de plantas C4 de más de 90 % (Saarinen et al., 2015). A pesar de sugerir la presencia de pastizales, la información del desgaste no es suficiente, por lo que se requieren de estudios adicionales, tales como mineralógicos y palinológicos.
5.6. PALEOAMBIENTE
Actualmente el relieve de la zona es irregular, con elevaciones pronunciadas hacia el sur (donde inicia la Sierra Madre del Sur) y hacia el norte, planicies que se conectan a la región fisiográfica de valles y lomeríos del centro del estado. Esta topografía debió de haber sido similar durante el Pleistoceno tardío, pues los afloramientos aledaños pertenecen a formaciones que datan del Cretácico tardío (ca. 91 - 71.5 Ma) y existen pocos indicios de depósitos posteriores más recientes.
La ocurrencia conjunta de M. columbi y Equus cf. E. conversidens también sugiere la presencia de zonas abiertas con pastizales en las partes planas, como lo indican diversos estudios con estos taxones en México (Bravo-Cuevas et al., 2011; Gutiérrez Bedolla et al., 2016; Marín-Leyva et al., 2016). Aparentemente esto entra en contraposición con el hábitat forestal sugerido para Bison latifrons (McDonald, 1981). Sin embargo, no existe evidencia que sugiera que efectivamente, este era el ambiente preferido de este bóvido y pudo habitar en otros tipos de ecosistema, como el inferido para San Dionisio Ocotepec.
6. Conclusiones
Los sedimentos de San Dionisio Ocotepec datan del Pleistoceno tardío, específicamente a la NALMA del Rancholabreano y consta de al menos tres taxones a nivel específico: Mammuthus columbi, Equus cf. E. conversidens y Bison latifrons. La presencia del mamut columbino y del onagro mexicano durante el Rancholabreano en México es relativamente común. La presencia de estas especies da conectividad a localidades de la región noroeste de Oaxaca con aquellas de Chiapas. Por otra parte, en este estudio corroboramos la presencia de Bison latifrons en los Valles Centrales del estado de Oaxaca, lo que marca la distribución más austral de la especie en el continente, extendiendo en más de 447 km su rango de distribución geográfica conocida.
La información estratigráfica sugiere que el ambiente sedimentario predominante en San Dionisio fue fluvial, con aporte principal de terrígenos y de forma secundaria, tobas, brechas y conglomerados. Considerando la evidencia de la topografía, la estratigrafía y de los restos fósiles, sugerimos que las partes planas de San Dionisio Ocotepec fueron ocupadas por pastizales.

