











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink1. Introduction
Neuroptera, the group of lacewings, is character-ised by a rather unique type of larva. Lacewing larvae are generally fierce ambush predators. While adult lacewings resemble certain butterflies or dragonflies, lacewing larvae are quite special in their morphology. Their mouthparts are strongly forward directed (prognath) and very prominent. Each mandible, or upper jaw, is interlocked with one of the maxillae, lower jaws, to form a stylet, a combined venom-injecting and sucking tool. This pair of stylets can reach a large size and represent up to one third of the entire length of the larva in some lacewing ingroups. The maxillae lack distal parts, or palps. The labium, or lower lip, is deeply integrated into the head capsule and is mainly recognisable by its distal parts, the palps (MacLeod 1964; Gepp 1984; Aspöck and Aspöck 1999, 2007).
The most prominent examples of lacewing larvae are possibly those of antlions, which catch their prey in many cases with trap funnels bur-rowed in sandy ground (Badano, 2012). Another well-known fraction of lacewing larvae is that of aphid lions, the larvae of the groups Chrysopidae (green lacewings) and Hemerobiidae (brown lace-wings). They are, as the name suggests, specialised on feeding aphids (e.g.Tauber et al., 2003). As the latter are severe plant pests, aphid lions have been successfully introduced as pest control some years ago, also due to their effectiveness and ecological benefits (Tauber et al., 2000).
Neuroptera is generally considered an early-appearing lineage of Holometabola (Grimaldi and Engel, 2005, p. 335), the ingroup of Insecta with immature forms addressed as ‘larvae’ due to their highly specialised eyes (for more information, see discussion in Haug, 2020). As compression fossils, lacewings have been commonly found as adults, whose wings seem to have a high potential of preservation (e.g.Makarkin, 2018). The fossil record of lacewing larvae is much scarcer and is largely restricted to fossil resins. The amber fossil record was recently summarised by Pérez-de la Fuente et al., (2020) and remains growing (e.g.Haug et al., 2019a, 2019b, 2020a, 2020b). The only two occurrences from compression fossils include a single specimen from the Cretaceous Crato formation (Martins-Neto et al., 2007; Herrera-Flórez et al., 2020, including a discussion about another presumed specimen) and a single specimen from the Eocene Green River formation (Dayvault et al., 1995).
Ambers that have so far yielded lacewing larvae have provided quite a number of specimens, especially Cretaceous ambers: at least 11 specimens originated from Cretaceous Lebanese amber (Whalley, 1980; Pérez-de la Fuente et al., 2018, 2019), four specimens from Cretaceous Spanish amber (Pérez-de la Fuente, 2012, 2016, 2020), three specimens from Cretaceous French amber (Perrichot, 2003; Perrichot et al., 2010; Wang et al., 2016), a single specimen from Cretaceous Canadian amber (Engel and Grimaldi, 2008) and a single specimen from New Jersey amber (Grimaldi, 2000, p. 284, fig. 3). Amber from Myanmar (also known as “Burmese amber”; for a recent discussion of the political background of this amber, see Haug et al., 2020c) yielded an astonishing amount of lacewing larvae (Xia et al., 2015; Wang et al., 2016; Liu et al., 2016, 2018; Wichard 2017; Zhang 2017; Badano et al., 2018; Lu et al., 2018; Makarkin, 2018; Haug et al., 2018, 2019a, 2019b, 2019c, 2020a, 2020b).
Especially these findings from the Cretaceous indicate an enormous diversity of larval forms of lacewings more than 100 million years ago compared to the modern fauna. On that regard, it is interesting to compare the diversity of larval forms with the younger ambers to better understand when the diversity of lacewing larvae could have declined. However, comparisons must be treated carefully, since ecological bias has to be taken into account (Solórzano-Kraemer et al., 2015, 2018). The resin-producing trees during the Cretaceous and Eocene are representatives of the group Coniferopsida (gymnosperms), while during the Miocene resin-producing trees are representatives of the group Magnoliopsida (angiosperms; Seyfullah et al., 2018), thus faunal assemblages inhabiting the trees could be different (e.g.Peris et al., 2017a).
Eocene Baltic amber has provided quite a number of larval lacewing specimens (Weidner, 1958; MacLeod, 1970; Larsson, 1978; Janzen, 2002; Weitschat and Wichard, 2002; Scheven, 2004; Weitschat, 2009; Wichard et al., 2009; Ohl, 2011; Makarkin et al., 2012; Wunderlich, 2012; Wedmann et al., 2013; Gröhn, 2015; Pérez-de la Fuente et al., 2020; Haug et al., 2020a, 2020b). Yet, there are still significantly fewer findings than from amber from Myanmar. Moreover, other Eocene ambers have hitherto not yielded lacewing larvae.
Even scarcer is the record from Miocene ambers. So far only eight specimens have been reported from Miocene Dominican amber (Wu, 1996; Poinar and Poinar, 1999; Grimaldi and Engel, 2005; Engel and Grimaldi, 2007). Dominican amber has been dated as early Middle Miocene (Iturralde-Vinent and MacPhee, 1996, 2019), whereas the age of Mexican amber from Chiapas is still in discussion; Solórzano-Kraemer, (2007) argued for a similar age to Dominican amber (~16 Mya) based on a correlation of foraminiferans, corals, decapodan crustaceans, mollusks, and pollen from different formations in Chiapas, Mexico, Hispaniola, and Puerto Rico. However, Serrano-Sánchez, et al. (2015) argued for an age of 22.8 million years based on the biostratigraphy of corals, mollusks, microfossils and strontium from one deposit, namely Campo La Granja mines, which belong to the La Quinta Formation. Ambers from Estrella de Belén deposit in the Salto de Agua Municipality, near Palenque, the origin of the amber described here, correspond also to those Miocene coarse quartz sands of the La Quinta Formation (Alvarado-Ortega et al., 2018). Both ambers, the Mexican and the Dominican ambers, come from similar environments, and the amber-producing tree in both instances belongs to species of the extant species group (genus) Hymenaea L. (Poinar, 1991; Poinar and Brown, 2002; Calvillo-Canadell et al., 2010).
Here we summarise the Miocene record of lacewing larvae from Dominican amber and report a new specimen from Miocene Mexican amber from the Estrella de Belén deposit.
2. Material and methods
2.1. Material
In the centre of this study is a single piece of amber from the Estrella de Belén deposit in the Salto de Agua Municipality, near to Palenque in Chiapas, Mexico. The specimen is stored in the amber collection at the Senckenberg Forschungsinstitut und Naturmuseum, Frankfurt am Main (SMF) under the accession number SMF Be 642a. Preserved inside the amber piece SMF Be 642, beside the lacewing larva, there are numerous syn-inclusions: one springtail (Collembola), six flies (Diptera: one of Psychodidae, four of Sciaridae, and one of Cecidomyiidae), one ant (Hymenoptera: Formicidae), one bark louse (Psocodea), as well as plant remains and quite some debris.
2.2. Documentation methods
Documentation was performed on a Keyence VHX 6000 digital microscope. The specimen was documented from one side only, the other side was inaccessible. It was illuminated in two different ways, once by coaxial cross-polarised light (Haug et al., 2013a) and once by unpolarised ring light. Under both illuminations, documentation was performed with a white and a black background. The combination providing the best contrast was used to be presented here. The built-in HDR function was used to optimise the images (cf. Haug et al., 2013b). Image stacks were recorded to over-come limitations in depth of field; several adjacent stacks were recorded to overcome limitations in field of view (Haug et al., 2008, 2011).
3. Results
All occurrences of depictions of lacewing larvae in Miocene ambers are listed chronologically. Cases in which the same specimen has been refigured are also included chronologically with reference to the original occurrence. While this includes a certain redundancy, it should represent the most complete way of cross-referencing, avoiding interpreting the same specimen as two independent occurrences.
Wu (1996) depicted a specimen of a lacewing larva preserved in Dominican amber (specimen 1; Figure 1A). The specimen (p. 136, fig. 224) is seen in dorso-lateral view; the body is slightly curled, but the head with the prominent stylets is well accessible. The author interpreted it as a larva of a brown lacewing (Hemerobiidae). Yet it remains unclear on which characters this interpretation is based. The overall appearance is in our view more compatible with an interpretation as a larva of a green lacewing (Chrysopidae).
Poinar and Poinar (1999) depicted two specimens of lacewing larvae preserved in Dominican amber. The first specimen (p. 86, fig. 94; figure plates between pp. 110, 111) was interpreted as the larva of an antlion (Myrmeleontidae). This specimen (specimen 2, Figure 1B) is well visible in ventral view. No indication of size was provided. The second specimen (figure plates between pp. 110, 111; p. 129, fig. 128) was interpreted as the larva of an owlfly (Ascalaphidae). The body of this specimen (specimen 3; Figure 1C) is less well visible due to camouflage with debris, yet the large mandibles are well apparent. No indication of size was provided. Poinar and Poinar (1999) did not provide the necessary details for a more accurate description of the systematic position, and also did not provide any accession number. The overall information of these specimens is therefore limited.
Scheven (2004) depicted one specimen of a lacewing larva preserved in Dominican amber (p. 142). The specimen (specimen 4; Figure 1D) was stated to be 2 mm in total length. The specimen is accessible in dorsal view.
Grimaldi and Engel (2005) depicted three specimens of lacewing larvae preserved in Dominican amber. The first one is a larva of the group Myrmeleontidae (accession number Morone Collection, M1282). The specimen (specimen 5; Figure 1E) was interpreted as a possible representative of the group Porrerus Navas, 1913. Length was stated to be 4.5 mm. A photomicrograph of the specimen shows that it is well preserved in clear amber, accessible in latero-dorsal view (Grimaldi and Engel 2005, p. 346, fig. 9.20). The second specimen is a representative of the group Ascalaphidae (accession number Morone Collection M3550). The specimen (specimen 6; Figure 2A) was interpreted as a representative of the group Ululodes Currie, 1900. Length was stated to be 8 mm. A photomicrograph of the specimen shows that it is well accessible in ventral view (p. 38, fig. 9.24). The third one is a larva of the group Chrysopidae (AMNH without further number), hence an aphid lion (specimen 7; Figure 2B). Length was stated to be 1.3 mm. A detailed drawing of the specimen shows that it is well preserved in latero-dorsal view, exhibiting short protrusions with long setae (p. 352, fig. 9.31). All three specimens have been re-figured in Engel and Grimaldi (2007).
Engel and Grimaldi (2007) depicted five specimens of lacewing larvae preserved in Dominican amber. The first one is a re-figure (p. 32, fig. 34) of the drawing of specimen 7, i.e. the aphid lion depicted in Grimaldi and Engel (2005). Here an accession number (DR-10-1454) was provided. The second one is a grey-scale version (p. 39, fig. 42) of the photomicrograph of specimen 5, the antlion larva depicted in Grimaldi and Engel (2005). Here accession number was given as MACT-1282. The specimen was interpreted as a possible representative of the species Porrerus dominicanus Stange and Poinar, 1996. Additionally an interpretive drawing (p. 40, fig. 43) was provided. The third specimen is a larva of the group Ascalaphidae (accession number MACT-1200). The specimen (specimen 8; Figure 2C) was interpreted as a representative of the group Ululodes, of a distinct species sp. 1. A photomicrograph (p. 45, fig. 47) and interpretive drawings of the habitus and the head (p. 46, fig. 48) were provided. Body length was stated to be about 5.9 mm (p. 44). The fourth specimen is also a representative of the group Ascalaphidae (accession number AMNH DR-10-2010). The specimen (specimen 9; Figure 2D) was interpreted as a representative of the group Ululodes and also of the distinct species sp. 1. An interpretive drawing of the head was provided (p. 47, fig. 49). Length of the head capsule was stated to be 1.5 mm. The fifth one is a grey-scale version (p. 49, fig. 50) of the photomicrograph of specimen 6, the owlfly larva depicted in Grimaldi and Engel (2005). Here accession number was given as MACT-3550. The specimen (specimen 6; Figure 2A) was interpreted as a representative of the group Ululodes and of the distinct species sp. 2. Additionally, an interpretive drawing of the head (p. 50, fig. 51) was provided.
Given the scarceness of data on lacewing larvae in Miocene amber, we used certain online resources as additional data sources (see discussion of use of websites as scientific source in Haug and Haug 2019; Haug et al. 2020a). In order to make these images available, we asked the copyright holders whether we could depict their images in this contribution. In total, we present images of six such specimens, all are preserved in Dominican amber. The first of these specimens is the larva of an owlfly (Ascalphidae). The image (Figure 3A) was kindly provided by TerraTreasures and Adventures 101 (http://terratreasures.com). Size of the specimen is unclear. The second of these specimens is the larva of an antlion (Myrmeleontidae). The image (Figure 3B) was kindly provided by Huang Yi-Jen (amber boy, huangyijen0708). Size of the specimen is unclear. Images of the third to sixth specimen were kindly provided by GlobalNatural-Resources. The third specimen (Figure 3C) is the larva of an antlion (Myrmeleontidae). It is slightly less than 3 mm long. The fourth specimen (Figure 3D) is the larva of an owlfly (Ascalaphidae). It is slightly longer than 4 mm. The fifth specimen (Figure 3E) is also a larva of an owlfly (Ascalaphidae). It is slightly less than 3 mm long. This image is also available in the online repository iStock by Getty Images (istockphoto.com). The sixth specimen (Figure 3F) is an aphid lion (Chrysopidae). It is about 4 mm long. These six specimens are not available for further detailed study, only the images are available. Due to the distinct scarcity of Miocene lacewing larvae, specimens found in different internet sources contribute to certain valuable information about the diversity that remains still to be discovered.
A new fossil specimen of a presumed owlfly larva (Ascalaphidae) is reported here (Figures 4A, 4B, and 5). The specimen originates from Mexican amber from Estrella de Belén deposit in the Salto de Agua Municipality, near Palenque in Chiapas, Mexico, and is part of the amber collection of the Senckenberg Forschungsinstitut und Natur-museum, Frankfurt am Main (accession number SMF Be 642a). The amber piece is rather large and thick.

Figure 1 Lacewing larvae preserved in Miocene Dominican amber from the literature. All simplified interpretive drawings; similar grey values indicate corresponding structures. Not to scale, specimens with more available details depicted larger. A. Specimen 1, aphid lion (Chrysopidae), based on Wu (1996, p. 136, fig. 224), size unknown. B, C. Based on Poinar and Poinar (1999), sizes unknown. B. Specimen 2 (p. 86, fig. 94 and figure plates between pp. 110, 111). C. Specimen 3 (figure plates between p. 110, 111; p. 129, fig. 128). D. Specimen 4, owlfly larva (Ascalaphidae), based on Scheven (2004, p. 142), size unknown. E. Specimen 5, antlion larva, possibly of Porrerus dominicanus Stange and Poinar. MACT-1282, based on Grimaldi and Engel (2005, p. 346, fig. 9.20; also Engel and Grimaldi 2007, p. 39, fig. 42 and p. 40, fig. 43), total size about 4.5 mm.

Figure 2 Lacewing larvae preserved in Miocene Dominican amber from the literature, continued. All simplified interpretive drawings; similar grey values indicate corresponding structures. Not to scale, specimens with more available details depicted larger. A, B. Based on Grimaldi and Engel (2005). A. Specimen 6 (p. 38, fig. 9.24; also Engel and Grimaldi 2007, p. 49, fig. 50), owlfly larva (Ascalaphidae), Ululodes sp. 2, MACT-3550, total size about 8 mm. B. Specimen 7 (p. 352, fig. 9.31; also Engel and Grimaldi 2007, p. 32, fig. 34), aphid lion (Chrysopidae), DR-10-1454, total size about 1.3 mm. C, D. Based on Engel and Grimaldi (2007). C. Specimen 8 (p. 45, fig. 47 and p. 46, fig. 48), owlfly larva (Ascalaphidae), Ululodes sp. 1, MACT-1200, body length about 5.9 mm. D. Specimen 9 (p. 47, fig. 49), owlfly larva (Ascalaphidae), Ululodes sp. 1, AMNH DR-10-2010, head capsule length about 1.5 mm.

Figure 3 Images of additional specimens of lacewing larvae preserved in Miocene Dominican amber. A. Owlfly; image from TerraTreasures and Adventures 101 (terratreasures.com). B. Antlion; image from Huang Yi-Jen (amber boy, huangyijen0708). C-F. Images from GlobalNaturalResources. C. Antlion. D, E. Owlfly. F. Aphid lion. Not to scale.

Figure 4 New owlfly larva from Miocene Chiapas amber, specimen SMF Be 642a. A. Composite image, black background, unpolarised ring light. B. Colour-marked interpretation of structures, arrowheads indicate stemmata. Abbreviations: 1-3 = teeth on stylet; ad = abdomen (posterior trunk); hc = head capsule; ms = mesothorax; mt = metathorax; pr = process or protrusion; pt = prothorax; sy = stylet (compound structure formed by mandible and maxilla); ta = trunk appendage (“leg”); te = trunk end.

Figure 5 Simplified restoration of the new owlfly larva from Miocene Chiapas amber, based on specimen SMF Be 642a.
Description: The larva is visible in dorsal view. Body is organised into (presumably) 20 segments (ocular segment + 19 post-ocular segments) and differentiated into head and trunk.
Head: formed by ocular segment and post-ocular segments 1-5. Head with distinct head capsule. Head capsule and protruding structures longer than trunk. Head capsule roughly square-shaped in dorsal view. Lateral rims with numerous prominent setae. Ocular segment recognisable by a pair of groups of simple eyes. Each group with at least three simple eyes (stemmata). Groups antero-laterally on head capsule, on distinct protrusions (“eye hills”). No external structures of post-ocular segments 1 and 2 apparent. Append-ages of post-ocular segments 3 and 4 presumably forming compound structures (stylets), yet only appendages of post-ocular segment 3 accessible, mandibles. Mandibles large, almost 1.5 times as long as head capsule, protruding forward (prognath). They are curved in the distal region, the tip is facing inwards. Three prominent teeth are present along the inside of the mandible. Additionally numerous setae are present on the inner and outer edge of the mandible.
Trunk: longer than wide. Anterior three segments (thorax) larger than further posterior ones (abdomen). Trunk segment 1 (prothorax; post-ocular segment 6) narrower than head. With a smaller protrusion on each side, bearing numerous setae. Trunk segment 2 (mesothorax; post-ocular segment 7) about as wide as the head. With two finger-like protrusions on each side. Each protrusion with numerous long setae. Trunk segment 3 (metathorax; post-ocular segment 8) sub-similar to preceding segment. Protrusions slightly shorter. Trunk segments 4-10 (abdomen segments 1-7; post-ocular segments 9-15) sub-similar. Shorter than preceding segments. Progressively narrower towards the posterior. Each segment with a finger-like protrusion on each side. Each protrusion with numerous long setae. Trunk segment 11 (abdomen segment 8; post-ocular segment 16) slightly longer than preceding segments, strongly narrowing towards the posterior. With a finger-like protrusion on each side. Each protrusion with numerous long setae. Trunk end (most likely representing undifferentiated abdomen segments 9-11; post-ocular segments 17-19) slender, longer than wide. With numerous long setae along the posterior end. More distal regions or trunk appendages (legs) are apparent. Details remain largely unclear, subdivision only partly apparent.
4. Discussion
4.1. Identity of the specimen
The overall shape and especially the mandible morphology allow to identify the specimen as a larva of Neuroptera and of Myrmeleontiformia (the group with all antlion-like lacewings). Three prominent teeth are in the modern fauna only found in larvae of antlions (Myrmeleontidae) and owlflies (Ascalaphidae). Yet, three teeth have also been present in other representatives of Myrmeleontiformia (Haug et al., 2019c).
Myrmeleontidae and Ascalaphidae share the presence of a tibio-tarsus (“fusion”) on trunk append-age 3 (metathorax leg). In the fossil described here it remains partly unclear whether there is a tibio-tarsus, this region of the appendage it is not well visible, but partly concealed under a bubble. Also, other features characterising the group Myrmeleontidae + Ascalaphidae (as put forward by Badano et al. 2017, p. 100) are not visible. Still the overall morphology makes it most likely that the specimen is the larva of a species of Ascalaphidae. In any case, this represents the second report of a lacewing from Mexican amber (the other one is an adult mantis lacewing, Mantispidae; Engel and Grimaldi 2007; p. 10-13, figs. 4-8), the first report of a lacewing larva from this amber, and possibly the first report of an owlfly (Ascalaphidae).
4.2. The Amber Record of Lacewing Larvae: Cretaceous Amber
Amber has fascinated various scientists, especially entomologists. Ambers from deep geological time have been considered as especially important because they provide data on the ecology of forest ecosystems millions of years old and on the evolutionary history of organisms and their behaviour which are otherwise rarely preserved in the fossil record. On that regard, finding the oldest fossil of a specific group is quite significant.
Triassic ambers have so far only yielded one representative of Insecta, a midge (Diptera), and two species of eriophyoidean mites (Schmidt et al., 2012). Therefore, the insect-rich Cretaceous ambers have been given quite some attention. This is also true for lacewing larvae. Lacewing larvae from the Lebanon represent the oldest such larvae (Pérez-de la Fuente et al., 2018, 2019). Younger Cretaceous ambers from New Jersey, Canada, Spain, France and Myanmar have yielded an astonishing diversity of lacewing larvae. Some of these larvae resemble modern forms, but others represent astonishingly different, aberrant-appearing forms. Myanmar amber is unparalleled concerning the output of lacewing larvae (Wang et al., 2016: about 30 specimens; mostly single specimens in Liu et al., 2016, 2018; Makarkin, 2018; Haug et al., 2019a, 2019b, 2019c, 2020a, 2020b; see also Pérez-de la Fuente et al., 2020). It appears that overall clearly more than 200 specimens are currently available in different Cretaceous amber collections. Given numerous additional specimens offered via internet by various amber traders, this seems to be only the tip of the iceberg.
4.3. The Amber Record of Lacewing Larvae: Baltic Amber
Younger ambers from the Eocene, especially Baltic amber, have likewise yielded lacewing larvae. For some lineages, Baltic amber provides the oldest record, as for example Sisyridae (at least two specimens; Wichard et al., 2009) and Coniopterygidae (at least one specimen so far; Weitschat and Wichard, 1998, 2002; more seem to be present). For some other lineages the Eocene record is more extensive concerning numbers of specimens than for the Cretaceous amber, as for example Nevrorthidae (at least 20 specimens so far; Haug et al., 2020b; seen more in the possession of traders). Further finds include larvae of Mantispidae (one specimen; Ohl, 2011), Berothidae (at least five specimens; Wedmann et al., 2013), Chrysopidae (three specimens: Scheven, 2004, p. 68, 69; Weitschat, 2009, p. 254, fig. 45). Hemerobiidae (one specimen; Makarkin et al., 2012), Ascalaphidae (at least two specimens; MacLeod, 1970; Weitschat and Wichard, 2002), Nymphidae (one specimen; MacLeod, 1970), Psychopsidae (at least twelve specimens; Haug et al., 2020a) and also some specimens that are clearly larvae of Neuroptera, but the exact affinity remains unclear (e.g.Larsson, 1978, see discussion in Haug et al., 2020a). In total, there are (at least) about 50 specimens of lacewing larvae preserved in Baltic amber. This number is strongly influenced by Nevrorthidae (especially one piece with seven larval specimens; Wichard et al., 2009, p. 113, fig. 07.19a) and Psychopsidae (silky lacewings), still this is a quite higher number compared to the Miocene record.
4.4. Why is the Number of Lacewing Larvae Lower for the Miocene Record?
The fossil record available in the literature and in the internet shows an apparent decrease in the presence of lacewing larvae in amber forests from the Cretaceous to younger fossil deposits. The Eocene record of lacewing larvae is less extensive than the Cretaceous one. The Miocene is even less extensive. Besides the specimen reported here, the Miocene record was so far restricted to specimens preserved in Dominican amber. However, Miocene amber deposits with bioinclusions, besides the Mexican amber, are known from Ethiopia (Bouju and Perrichot, 2020), from China (Shi et al., 2014), from Peru (Antoine et al., 2006), and from New Zealand (Schmidt et al., 2018), all discovered relatively recent so that the amount of amber with inclusions is still small. In younger copal or resin, only one case of a lacewing larva is known from Colombian copal or resin (age unknown) (Kobbert, 2013, p. 55, fig. 206). Adults of the group Coniopterygidae have been described from copal or resin of unknown age from Madagascar and the African mainland (Engel, 2004), a group also reported in Miocene Dominican amber and in Eocene Cambay amber (Grimaldi et al., 2013). Dominican amber has been considered to be a “highly fossiliferous” amber (e.g.Arillo and Ortuño, 2005; Penney, 2016); in the light of this view, it is somehow surprising that there are so few specimens of lacewing larvae.
The simple numbers give a pattern that shows a decrease of available lacewing larvae over time: The highest number is available for the Cretaceous, the number is lower in the Eocene, and finally lowest in the Miocene (Figure 6). In how far could this represent a true pattern and not a sampling or ecological bias? The Cretaceous offers several ambers with a wide geographic coverage (yet this is also the case for Miocene and Eocene ambers) including North America (Canadian amber), Europe (French and Spanish amber) and Asia (Lebanon, Myanmar). Especially Myanmar amber has provided a high output of overall specimens due to several factors (Haug et al., 2020c).
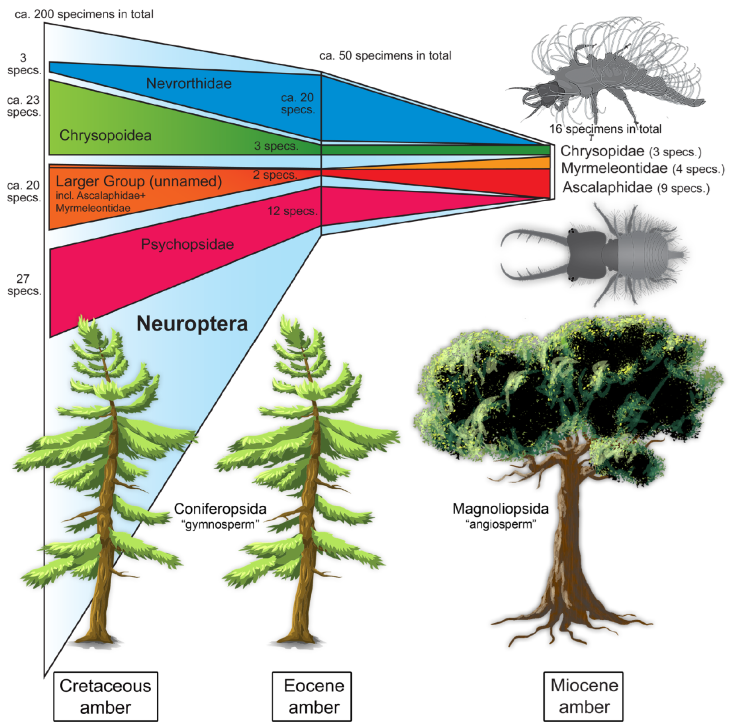
Figure 6 Depiction of number of larval lacewing specimens preserved in amber per time slice. Numbers given for the entire group of Neuroptera, as well as for some ingroups: Nevrorthidae, Psychopsidae (silky lacewings), Chrysopoidea (including its ingroup Chrysopidae; green lacewings, aphid lions; upper right image) and a larger unnamed group including Myrmeleontidae (antlions) and Ascalaphidae (owlflies; lower right image). The different sources of amber are indicated by schematic representatives of the groups Coniferopsida (“gymnosperms”) and Magnoliopsida (“angiosperms”). Note the dominance of the groups Psychopsidae and Nevrorthidae in the Eocene.
It may therefore not be surprising that so many new lacewing larvae have been reported from Myanmar amber (Engel and Grimaldi, 2008; Xia et al., 2015; Wang et al., 2016; Liu et al., 2016, 2018; Zhang, 2017; Badano et al., 2018; Haug et al., 2018, 2019a, 2019b, 2019c, 2020a, 2020b; Herrera-Flórez et al., 2020). Yet, the other Cretaceous ambers have also provided quite a number of significant specimens (Engel and Grimaldi, 2008; Pérez-de la Fuente, 2012, 2016, 2018, 2019, 2020; Haug et al., 2020a). The resin-producing trees, which later generated the Cretaceous ambers, were representatives of Coniferopsida (gymnosperms), mostly of the ingroup Araucariaceae but also of the ingroups Cupressaceae, Cheirolepidiaceae or Pinaceae (Seyfullah et al., 2018).
The Eocene Baltic amber has a long research tradition. Hence it should not be surprising that there are quite some lacewing larvae known from Baltic amber. The resin-producing tree of the Eocene Baltic amber was also a representative of the group Coniferopsida of the ingroup Pinaceae or Sciadopityaceae (Wolfe et al., 2016). Yet, if we look closer, the relatively high number of larvae is mostly due to specific lineages within lacewings, Psychopsidae and Nevrorthidae (see above). Larvae of other groups are quite rare, as there is, for example, only a single larva of Nymphidae known (MacLeod, 1970). We might speculate that larvae of Nevrorthidae and Psychopsidae became trapped in resin more easily than other lacewing larvae.
Today Nevrorthidae and Psychopsidae show quite restricted ranges of occurrence that makes it more unlikely to find them in Miocene American ambers. Representatives of Nevrorthidae nowadays occur in Mediterranean Europe, China, Japan and Australia (Aspöck and Aspöck, 2010). Representatives of Psychopsidae occur today in South-East Asia, southern Africa and Australia (Peng et al., 2011). There are several examples of groups, that nowadays have representatives restricted to Australasia, but which have fossil forms preserved in Dominican amber, in total 18 species groups (genera); this includes lacewings of the ingroup Aleuropteryginae (Grimaldi et al., 2013). Still, we cannot expect to find larvae of Psychopsidae and Nevrorthidae, but we can also not fully exclude it. The occurrence of representatives of Nevrorthidae in Dominican amber is furthermore unlikely as the deposit is notoriously poor concerning representatives of merolimnic groups of Insecta, i.e. those with aquatic larvae (Grund, 2006). The absence of these two lineages might, in any case, be part of the effect that the number of known lacewing larvae from this amber is rather low.
The high number of larvae from Cretaceous ambers may not be easily explained by sampling bias. An ecological bias of lacewings in general was discussed by Engel and Grimaldi (2007) relating their abundance and diversity in ambers to the abundance of sternorrhynchan hemipterans.
The latter are rare in Miocene ambers. Actuotaphonomic studies also support the low abundance of lacewings (larvae and adults, however within the low abundance there are more adults than larvae) and sternorrhynchan hemipterans in modern-day resin and sticky traps in Hymenaea-type forests (ingroup of Magnoliopsida, “angiosperm”) in Mexico and in Madagascar (Solórzano-Kraemer et al., 2015, 2018). Notably, in the sticky traps in the Hymenaea-type forest in Mexico, where sternorrhynchans were lightly abundant (89 specimens), five lacewings (four larvae and one adult) were collected (Solórzano-Kraemer et al., 2015). In Malagasy resin collected in 2013, only one adult of the group Neuroptera (0.02%) has been recognized from 3,923 representatives of Euarthropoda (Solórzano-Kraemer et al., 2018). On the contrary, in modern resins originating from species of Pinus L. and Picea A. Dietr, 1824 (both ingroups of Coniferopsida, “gymnosperms”) collected by Zherikhin et al., (2009) in Northern Eurasia lace-wings were more common, from 0.3 to 3.7% in the different samples.
However, it has to be noticed that these data are not specific of larvae. Pérez-de la Fuente et al., (2020) related the scarcity of antlion larvae (Myrmeleontidae) in amber to their soil-dwelling life-styles. This implies resin production at soil level or the drip of the resin in the soil to preserve this kind of organism. The antlion larvae seem to be more frequent in Miocene amber than in Cretaceous or Eocene ambers.
It is well known that the more active organisms living in or in closer relation to the resin-producing tree have a higher probability to become entombed in resin and therefore preserved in amber (Solórzano-Kraemer et al., 2018). Miocene ambers were produced by trees of the groups Fabaceae and Dipterocarpaceae, both ingroups of Magnoliopsida (“angiosperms”) with their own specific faunas.
Some elements of the fauna abundant during the Miocene are today abundant and associated with trees of the group Hymenaea (ingroup of Fabaceae and Magnoliopsida, “angiosperms”) but absent in forests dominated by trees of the group Coniferopsida (“gymnosperms”; e.g.Peris et al., 2015, 2017a).
Some other faunal elements related to trees of the group Coniferopsida during the Cretaceous, such as pollinators, including lacewings, did not undergo an evolutionary shift to trees of the group Magnoliopsida (Peris et al., 2017b). In this regard, it is interesting that the most common forms in Miocene ambers are larvae of owlflies (nine specimens) and antlions (four specimens; additionally, three aphidlion specimens). Owlfly(-like) and ant-lion larvae are quite rare in Eocene ambers, and in the case of antlions also in Cretaceous ambers. Hitherto, there is no study of differing ecological biases concentrated on larvae on resin-producing trees of the groups Magnoliopsida vs. Coniferopsida. Such an actuotaphonomic study could test whether the abundance of lacewing larvae is related to an ecological bias. For further exploring this aspect, we will need more quantitative data from unbiased amber, copal and resin collections.
5. Outlook
Fossils from Cretaceous and Eocene amber have already been used for quantitatively comparing morphology of lacewing larvae through time (Haug et al., 2020a). Such studies can reveal interesting patterns for improving our understanding of the changes of lacewing diversity in particular, but also losses of diversity in general. The Miocene could potentially add another time slice to such a comparison, yet the overall low number of available lacewing larvae is partly challenging in this aspect.
It may still be a matter of focussed interest in adults that simply leads to the effect that larvae, yet being in fact present, do not surface. We hope that the present contribution can also serve to encourage more people, professional and amateur, to report on more lacewing larvae from Miocene ambers. In this way, we hope to be able to integrate these ambers into larger quantitative analyses.

